凯氏拟小球藻己糖激酶基因的克隆及其在不同培养条件下的表达分析
2017-11-21崔红利赵佳琳
陈 军, 崔红利, 赵佳琳, 秦 松
凯氏拟小球藻己糖激酶基因的克隆及其在不同培养条件下的表达分析
陈 军1, 2, 崔红利3, 赵佳琳1, 2, 秦 松1
(1. 中国科学院烟台海岸带研究所海岸带生物学与生物资源利用重点实验室, 山东烟台 264003; 2. 中国科学院大学, 北京 101418; 3. 大连大学生命科学与技术学院, 辽宁大连 116622)
为研究凯氏拟小球藻()糖代谢分子机制, 本研究采用RT-PCR和RACE技术从凯氏拟小球藻中克隆了己糖激酶基因HK(GenBank ID: AHF54566), 并对其自养、异养、混养条件下的转录表达进行分析。结果表明, 该序列的cDNA全长为1 844 bp, 开放阅读框1 389 bp, 编码462个氨基酸。该蛋白的相对分子质量为49.73, 等电点为6.98。实时荧光定量PCR结果显示, 以自养培养条件为对照, 异养培养和混养培养条件下,HK均能够发生明显上调, 且混养条件下上调量比异养条件下上调量更多, 说明HK可能在凯氏拟小球藻利用外源糖的过程发挥重要作用, 并且光信号对于凯氏拟小球藻利用外源糖可能存在调控作用。这些研究结果为进一步阐明HK的功能及其作用机制奠定了分子基础。
凯氏拟小球藻(); 己糖激酶; 基因克隆; 表达分析
凯氏拟小球藻()属于绿藻门, 颤藻目, 小球藻科。Huss等[1]基于形态学特征、DNA碱基组成以及利用分子标记18S rRNA和SSU rRNA进行系统发育分析等多种手段, 认为Fott et Nova′kova应属于小球藻属。Krienitz等(2004)通过克隆18S和ITS2序列, 分别用18S rRNA、ITS2以及18S rRNA和ITS2联合分子标记, 构建系统发育树, 结合形态学特征认为其应属于一个新属——拟小球藻属(), 也是目前较为公认的划分方法[2-3]。凯氏拟小球藻能够利用光合作用进行自养, 也可利用外源葡萄糖进行异养, 以及在光和外源葡萄糖的作用下进行混养。它生长速率快, 含有丰富的脂肪酸, 目前广泛应用在废水处理[4-5], 生物能源[6], 生物固碳[7]等领域。
Wang等[8]模拟有机废水培养凯氏拟小球藻生产微藻油脂, 发现以自养培养为对照, 混养条件下小球藻能够积累更多的脂肪酸以及更多以C16和C18为主的中性脂。Wang等[9]通过分别或同时添加外源葡萄糖和硝酸盐, 发现凯氏拟小球藻能够明显提高脂肪酸含量。Li等[4]发现不同光强可影响凯氏拟小球藻去除废水中有机质的效果, 且相对于自养条件下, 加入光照后混养条件下均能高效去除有机质。基于以上报道, 可见添加外源葡萄糖会促进凯氏拟小球藻生长及脂肪酸合成, 且添加光照和不添加光照情况下, 凯氏拟小球藻生长及脂肪酸代谢也不相同, 说明凯氏拟小球藻利用外源葡萄糖的代谢机制并不相同。而且凯氏拟小球藻代谢外源葡萄糖的分子机制尚不清晰。
己糖激酶是一类能够催化己糖发生磷酸化作用的酶类, 既能够调控植物体内贮存糖和游离糖的速率, 也能够调控糖酵解和氧化戊糖磷酸途径的代谢速率, 对于植物碳流分配具有重要作用。同时, 己糖激酶也可以作为细胞葡萄糖感受器感知糖信号, 从而触发糖信号传递, 即具有催化功能和调节功能[10-11]。在生物体内, 己糖经己糖激酶磷酸化后可进入糖酵解途径, 进而为植物的生理活动提供能量和中间代谢产物[12]。目前, 己糖激酶已在拟南芥、水稻等生物中被克隆鉴定[13-14], 但仍未在藻类中被同源克隆、鉴定。作者通过利用已经被功能鉴定的己糖激酶序列作为源序列, 利用同源比对的方法在莱茵衣藻()、团藻()、胶球藻C-169(C-169)、可变小球藻(NC64A)、蓝隐藻()、海洋球石藻()、裂殖壶藻()等微藻基因组序列进行搜索, 发现编码己糖激酶的基因序列广泛存在。
本研究利用聚合酶链式扩增(PCR)和cDNA末端快速扩增技术(RACE, rapid-amplification of cDNA ends)克隆获得凯氏拟小球藻己糖激酶的基因编码序列, 并运用实时荧光定量表达技术对不同培养(自养、异养、混养)条件下凯氏拟小球藻转录水平的表达情况进行了分析, 以期为探讨凯氏拟小球藻利用外源葡萄糖代谢分子机制提供参考, 为通过代谢工程手段构建优良藻株奠定基础。
1 材料和方法
1.1 实验材料及培养条件
本研究所用凯氏拟小球藻()保存于中国科学院烟台海岸带研究所海岸带生物学与生物资源利用重点实验室。凯氏拟小球藻接种于f/2培养基, 于25℃光照条件下静置培养, 光照强度为2 000 lx, 光/暗周期为12 h/12 h。异养培养葡萄糖质量浓度为10.0 g/L, 24 h黑暗培养; 混养培养葡萄糖质量浓度为10.0 g/L, 于25℃光照条件下静置培养, 光照强度为2000 lx, 持续给光。
1.2 凯氏拟小球藻己糖激酶基因的克隆
取处于对数生长期的凯氏拟小球藻细胞, 用TRIzol试剂提取总RNA, 使用1%的琼脂糖凝胶电泳进行检测, 并利用超微量分光光度计测定RNA浓度和纯度(260/280)。随后, 参照TaKaRa公司的PrimeScriptTMRT reagent Kit with gDNA Eraser (Perfect Real Time)试剂盒说明书合成cDNA, 用作克隆己糖激酶cDNA的模板。从GenBank中下载莱茵衣藻()、团藻()、胶球藻C-169(C-169)、可变小球藻(NC64A)编码己糖激酶基因的cDNA序列进行比对分析, 设计简并引物, 引物由上海生工生物工程有限公司合成。
以cDNA为模板, 用合成的简并引物进行RT-PCR 扩增HK基因的cDNA片段。反应程序为: 94℃预变性5 min; 94℃ 45 s, 55℃ 30 s, 72℃ 130 s, 30个循环; 72℃延伸10 min, 4℃终止。反应结束后, PCR反应产物经1%的琼脂糖凝胶电泳检测。对目的片段进行胶回收后连接于pMD18-T载体上, 转化大肠杆菌DH5α, 挑取阳性菌落, 菌液PCR鉴定正确后由上海生工生物工程有限公司测序。
根据简并引物扩增得到的已知序列, 分别设计HK的3′RACE和5′RACE引物(HK31/HK32和HK51/HK52)。利用Clontech公司的SMARTerTMRACE cDNA Amplication Kit试剂盒, 按照说明书分别扩增HK的3′端和5′端。扩增获得的结果经过连接、转化后, 挑取阳性菌落由上海生工生物工程有限公司测序。根据同源克隆片段和RACE测序结果, 拼接获得目的基因全长。
1.3 己糖激酶基因的生物信息学分析
将测序获得的片段, 利用软件DNAStar7.1 (DNASTAR Inc. USA)进行拼接得到目的基因cDNA全长序列。利用序列处理在线工具包(http: //www. bio-soft.net/sms/)将得到的cDNA序列翻译成氨基酸序列, 并用ORF查找器找到目的基因的ORF。通过在线BLAST程序对目的基因进行鉴定, 然后下载同源性相似序列, 利用本地ClustaW软件与目的基因进行多序列比对。利用软件ExPASy Compute pI/Mw tool[15]用来检测目的基因编码的蛋白相对分子质量(Mw)和等电点(pI)。最后, 基于多序列比对结果选择保守区, 利用jModelTest软件选择进化模型, 然后利用phyML软件中的最大似然法(Maximum likelihood)构建进化树。
1.4 不同培养条件下, 己糖激酶基因表达量分析
根据HK的cDNA序列设计荧光定量表达特异性引物(HKQF/HKQR), 同时选择凯氏拟小球藻18s rRNA序列作为内参, 设计荧光定量表达特异性引物(18SF/18SR), 引物序列见表1。设计引物后, 利用普通PCR对其进行验证, 确保没有引物二聚体出现以及引物的特异性。采用SYBR Premix Ex Taq II荧光定量试剂盒进行荧光定量PCR。每个样品设置3个平行, 荧光定量PCR数据采用相对定量2–DDCt法进行分析, 最后利用Origin 8.0进行标准偏差计算和作图。

表1 本研究所用的引物
2 结果与分析
2.1 凯氏拟小球藻己糖激酶全长cDNA克隆
本研究以凯氏拟小球藻总RNA反转录得到的cDNA为模板, 设计引物HXF/HKR(表1), 进行PCR扩增, 获得cDNA中间片段, 长度为237 bp。将该片段氨基酸序列与胶球藻(C-169)的HK序列比对发现同源性为57%、与微囊藻(CCMP1545)的同源性为68%、与莱茵衣藻()的同源性为58%。根据中间片段, 分别设计特异性引物(HK51/HK52和HK31/HK32), 通过cDNA末端快速扩增技术扩增, 获得该序列5′端和3′端, 长度分别为311 bp和1 404 bp。利用所获得的中间片段、5′端和3′端序列进行拼接, 获得己糖激酶基因的cDNA全长。该基因cDNA全长1 844 bp(NCBI 注册号: AHF54566.1), 包含1 389 bp的开放阅读框, 编码462个氨基酸, 5′非编码区(5′-UTR)序列长39 bp, 3′非编码区(3′-UTR)序列长436 bp(图1)。利用ExPASy Compute pI/Mw tool软件分析显示, 该序列的相对分子质量为49.73 ku, 等电点为6.98。
2.2 凯氏拟小球藻己糖激酶氨基酸序列比对及蛋白结构分析
利用翻译获得的己糖激酶氨基酸序列, 与来自其他9种藻类的己糖激酶氨基酸序列(图1)进行比对分析, 发现序列间同源性较高。功能结构域分析表明,HK蛋白含有ATP结合位点(CD1: ADLGGTNRV; CD4: IGTGIN; CD7: XDGXF), 己糖激酶结合位点(CD2: LGFFSF), 葡萄糖激酶结合位点(NDEE), 详见图2。由此, 提示凯氏拟小球藻己糖激酶基因序列不仅仅可以催化葡萄糖, 也可以催化其他己糖。利用软件SOPMA[16]对HK基因编码蛋白的二级结构预测, 预测结果显示, α螺旋占的比例最高, 为38.1%, 其次为无规卷曲, 占比31.82%, 延伸链和β-折叠分别为18.83%和11.26%。由此, 说明己糖激酶主要由α螺旋和无规卷曲组成, 属于混合型蛋白。
2.3 凯氏拟小球藻己糖激酶基因进化分析
本文比对了来自细菌、藻类和拟南芥等不同物种共24个己糖激酶基因的氨基酸序列, 并利用mega5和Phyml软件构建进化树, 分析了凯氏拟小球藻HK基因的分类归属以及该基因与其他物种之间的基因进化关系(图3)。从图中可以看出, 25个物种的HK共聚成7组, 自展值分别为57/86/100/100/ 88/99/100/88/88。依据己糖激酶基因进行分子系统发育分析, 可见凯氏拟小球藻与可变小球藻(NC64A)聚在一起, 然后和可变小球藻()、胶球藻(C-169)聚在一起, 与传统分类较为一致, 说明己糖激酶基因的保守性较强, 能够很好地反映各物种之间的进化距离。

图1 凯氏拟小球藻中编码己糖激酶的核苷酸序列和氨基酸序列。
阴影部分代表ATP和己糖结合位点, 方框标记的分别代表起始密码子、终止密码子、加尾信号序列
The initiation (ATG) and termination codons (TAG) are marked with an underline. Crucial residues of ATP and hexose-binding sites are highlighted by gray boxes
2.4 不同培养条件下CkeHK基因转录水平的变化
为探讨凯氏拟小球藻利用外源糖代谢机制, 本研究利用实时荧光定量表达技术, 以自养培养为对照, 对异养和混养条件下的HK基因转录水平进行了分析(图4)。从图4可以看出, 在异养情况下, 当培养至8 h,HK基因转录本上调1.38倍。培养至20~44 h,转录本出现降低。随后, 当培养44~56 h, 转录本逐渐升高且在第56小时达到最高, 是正常表达量的6.33倍。继续延长培养时间, 己糖激酶转录本表达量有所下降; 相对于自养情况为对照, 在混养情况下, 己糖激酶转录本表达量在第44小时开始出现明显上调, 培养至56 h达到最高, 为正常表达量的13.69倍, 随后出现下降。由此可见, 相比较自养, 在异养和混养培养下, 己糖激酶HK可能在凯氏拟小球藻利用外源糖方面发挥重要作用。同时, 混养培养过程中,HK的转录上调表达量远高于异养培养过程中的上调表达量, 可见混养培养过程中光信号的参与, 对己糖激酶代谢外源糖可能也具有调控作用。

图2 凯氏拟小球藻CkeHK的序列比对及结构域分析
●代表ATP结合位点 ▲代表己糖激酶结合位点
Crucial residues of the ATP and glucose-binding sites are highlighted by black circles and black triangles

图3 不同物种来源的己糖激酶序列系统进化分析
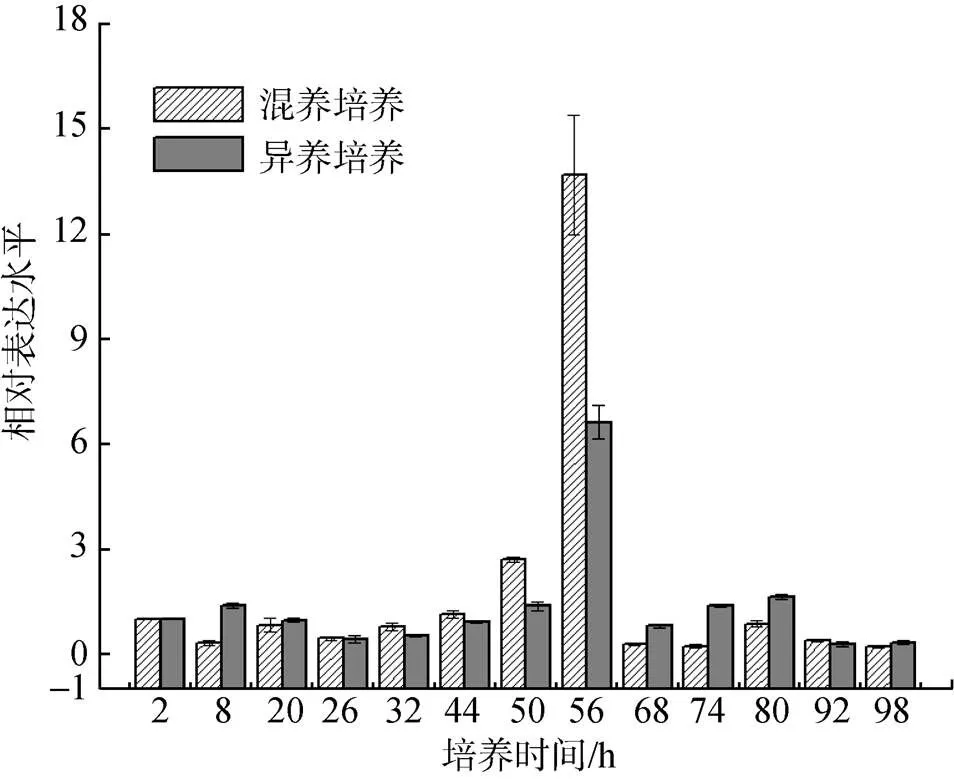
图4 不同培养条件下CkeHK的转录表达情况
3 讨论
糖类物质对微藻及植物的生长、发育、胁迫应激等方面有重要作用, 它不仅作为呼吸底物为微藻及植物提供生长发育所需要的能量, 也是合成氨基酸、脂肪酸、淀粉等重要物质的碳库。同时, 糖类还被认定为具有类似植物激素作用的信号分子, 参与植物的糖信号转导过程, 对于合成脂肪酸等代谢产物具有重要调控作用[17]。己糖激酶是植物体呼吸代谢过程中的关键酶, 可通过磷酸化作用催化外源糖类物质。同时, 通过添加己糖激酶抑制剂D-甘露酮糖可阻断2-脱氧葡萄糖介导的对光合基因表达的抑制作用, 说明己糖激酶是植物糖信号感知的感受器[18-19]。最近, 通过解析拟南芥中己糖激酶结构, 也证明己糖激酶既具有催化作用也具有感知作用[20]。
本研究以能够进行自养、异养、混养的凯氏拟小球藻为研究材料, 克隆获得己糖激酶基因的cDNA序列, 同源性分析显示, 其与莱茵衣藻中己糖激酶序列相似性较高。结构分析发现, 该基因编码氨基酸序列氮端具有多样性, 这些特点与在拟南芥和酵母中发现的己糖激酶具有相似性。同时, 依据己糖激酶序列构建的系统发育树可见, 各物种的聚类关系与传统经典分类较为一致, 说明己糖激酶基因在各个物种中是相对保守的, 也提示己糖激酶在凯氏拟小球藻的糖代谢过程中发挥重要作用。该基因是首次从微藻中被克隆获得, 也将为研究其他微藻糖代谢提供参考。
Chen等[21]通过异养、混养培养小球藻(), 观测细胞分裂及淀粉合成、脂肪酸合成, 提示混养培养小球藻过程相较于异养培养, 光可能发挥两方面的作用: (1)光能够促进细胞增殖和生物量提高; (2)加光照后, 小球藻细胞合成脂肪酸含量减少, 淀粉合成量增加。由此, 说明光照对于小球藻如何利用外源葡萄糖具有调控作用, 但具体调控机制目前尚不清晰。该结果与本研究中发现的光信号能够调控凯氏拟小球藻利用外源葡萄糖较为一致。此外, Jang等[12, 22]发现拟南芥中己糖激酶能够抑制编码光合作用相关蛋白(核酮糖-1, 5-二磷酸羧化酶/加氧酶和捕光叶绿素a/b结合蛋白)基因的表达, 提示己糖激酶是拟南芥中光信号与糖信号协同调控网络的关键元件。光照和外源葡萄糖都具有双重作用, 一方面都可以为藻株提供能量, 另一方面也都是具有调控作用的信号因子。开展光和糖如何协同调控小球藻生长及脂肪酸合成研究, 将对于创新培养工艺提高小球藻生物量及脂肪酸合成, 以及构建性能优良的藻株等都具有重要意义, 这也是日后工作将继续深入的重点。
4 结论
[1] Huss V A R, Frank C, Hartmann E C, et al. Biochemical taxonomy and molecular phylogeny of the genus(Chlorophyta)[J]. Journal of Phycology, 1999, 35: 587-598.
[2] Yamamoto M, Kurihara I, Kawano S. Late type of daughter cell wall synthesis in one of the Chlorellaceae,(Chlorophyta, Trebouxiophyceae)[J]. Planta, 2005, 221: 766-775.
[3] Krienitz L, Hegewald E H, Hepperle D, et al. Phylogenetic relationship ofandgen. nov (Chlorophyta, Trebouxiophyceae)[J]. Phycologia, 2004, 43: 529-542.
[4] Li Y C, Zhou W G, Hu B, et al. Effect of light intensity on algal biomass accumulation and biodiesel production for mixotrophic strainsandcultivated in highly concentrated municipal wastewater[J]. Biotechnology and Bioengineering, 2012, 109: 2222-2229.
[5] Ruan M, Li Y, Cheng Y. 模拟自然光周期下外源性CO2对城市污水藻类生长和污水处理的影响(摘选)[J]. 农业工程, 2013, 3(1): 101-104. Ruan Megan, Li Yecong, Cheng Yanling. Effect of exogenous CO2on algae growth and wastewater treatment under simulated natural light/dark cycle using municipal wastewater as feedstock ( Extracts)[J]. Agricultural Engineering, 2013, 3(1): 101-104.
[6] 江丽丽, 温小斌, 耿亚洪, 等.一株产油微藻的筛选及分子鉴定[J].水生生物学报, 2013, 37(4): 606-612. Jiang Lili, Wen Xiaobin, Geng Yahong, et al. A newly selected lipid-rich microalgae strain and its molecular identification[J]. Acta Hydrobiologica Sinica, 2013, 37(4): 606-612.
[7] 李林, 王帅, 郑立. 海洋微藻固碳及其培养技术的研究进展[J]. 海洋科学, 2015, 39(3): 135-140. Li lin, Wang Shuai, Zheng Li. Research progress of marine microalgae on carbon dioxide biofixation and its cultivation technology[J]. Marine Sciences, 2015, 39(3): 135-140.
[8] Wang Y, Chen T, Qin S. Differential fatty acid profiles ofgrown with organic materials[J]. Journal of Chemical Technology and Biotechnology, 2013, 88: 651-657.
114 Mechanical valve dysfunction caused by warfarin anticoagulant: a case report
[9] Wang Y, Chen T, Qin S. Heterotrophic cultivation offor fatty acids production by carbon and nitrogen supplements[J]. Biomass & Bioenergy, 2013, 47: 402-409.
[10] Moore B D, Sheen J. Plant sugar sensing and signaling - acomplex reality[J]. Trends in Plant Science, 1999, 4: 250-250.
[11] Moore B, Zhou L, Rolland F, et al. Role of theglucose sensor HXK1 in nutrient, light, and hormonal signaling[J]. Science, 2003, 300: 332-336.
[12] Claeyssen E, Rivoal J. Isozymes of plant hexokinase: Occurrence, properties and functions{Claeyssen, 2007 #55}[J]. Phytochemistry, 2007, 68, 6: 709-731.
[13] Karve A, Rauh B L, Xia X X, et al. Expression and evolutionary features of the hexokinase gene family in[J]. Planta, 2008, 228: 411-425.
[14] Cho J I, Ryoo N, Ko S, et al. Structure, expression, and functional analysis of the hexokinase gene family in rice (L.)[J]. Planta, 2006, 224: 598-611.
[15] Bjellqvist B, Hughes G J, Pasquali C, et al. The focusing positions of polypeptides in immobilized pH gradients can be predicted from their amino-acid-sequences[J]. Electrophoresis, 1993, 14: 1023-1031.
[16] Geourjon C, Deleage G. SOPMA: Significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments[J]. Computer Applications in the Biosciences, 1995, 11: 681-684.
[17] Koch K E. Carbohydrate-modulated gene expression in plants[J]. Annual Review of Plant Physiology and Plant Molecular Biology, 1996, 47: 509-540.
[18] Loreti E, De Bellis L, Alpi A, et al. Why and how do plant cells sense sugars[J]. Annals of Botany, 2001, 88: 803-812.
[19] Jang J C, Sheen J. Sugar sensing in higher plants[J]. Trends in Plant Science, 1997, 2: 208-214.
[20] Feng J, Zhao S, Chen X M, et al. Biochemical and structural study ofhexokinase 1[J]. Acta Crystallographica Section D-Biological Crystallography, 2015, 71: 367-375.
[21] Chen T P, Liu J, Guo B B, et al. Light attenuates lipid accumulation while enhancing cell proliferation and starch synthesis in the glucose-fed oleaginous microalga[J]. Scientific Reports, 2015, 5: 14936.
[22] Jang J C, Sheen J. Sugar sensing in higher plants[J]. Plant Cell, 1994, 6: 1665-1679.
Molecular cloning of hexokinase fromand its expression analysis under different trophic modes
CHEN Jun1, 2, CUI Hong-li3, ZHAO Jia-lin1, 2, QIN Song1
(1. Key Laboratory of Coastal Biology and Bioresource Utilization, Yantai Institute of Costal Zone Research, Chinese Academy of Sciences, Yantai 264003, China; 2. University of Chinese Academy of Sciences, Beijing 101418, China; 3. College of Life Science, Dalian University, Dalian 116622, China)
cDNA (GenBank ID: AHF54566) encoding hexokinase (termedHK) from the green algawas cloned and sequenced to understand the response to the presence and uptake of glucose at the molecular level. The transcriptional expression patterns ofHK were observed under phototrophic, heterotrophic, and mixotrophic culture conditions. The results indicated that theHk cDNA was 1, 844 base pairs (bp) long, with an open reading frame (ORF) of 1 389 bp encoding 462 amino acids, with a calculated molecular mass of 49.73 kDa and an estimated isoelectric point of 6.98. Under the phototrophic condition as control, the transcriptional profiles ofHk showed that it could be upregulated under the heterotrophic and mixotrophic culture conditions. The mRNA level ofHK under the mixotrophic culture condition could be upregulated more than that under the heterotrophic culture condition. These results suggest that light may play an important role in regulating the role of hexokinase in metabolizing exogenous glucose. These findings provide us valuable information for exploring the mechanism of metabolizing glucose in.
; hexokinase; molecular clone; transcript expression
(本文编辑: 梁德海)
Q786
A
1000-3096(2017)07-0001-08
10.11759/hykx20170225001
2016-12-22;
2017-03-10
国家重点研发计划项目(2016YFF0202304); 国家自然科学基金面上项目(41376139); 烟台市科技计划项目(2016JHZB007)
[National Key Research and Development Program-China, No.2016YFF0202304; National Natural Science Foundation of China, No.41376139; Science and Technology Program of Yantai City, No.2016JHZB007]
陈军(1989-), 男, 安徽阜阳人, 博士生, 主要从事微藻功能基因组学研究, E-mail: junchen@yic.ac.cn; 秦松(1968-), 通信作者, 男, 山东莱州人, 研究员, 博士生导师, 主要从事分子藻类学研究, E-mail: sqin@yic.ac.cn
Dec. 22, 2016
