术后疼痛对大鼠认知功能障碍的影响及其机制
2017-11-16任建光吴雅娟王文军
任建光,吴雅娟,王文军
(儋州市人民医院,海南儋州571799)
术后疼痛对大鼠认知功能障碍的影响及其机制
任建光,吴雅娟,王文军
(儋州市人民医院,海南儋州571799)
目的探讨术后疼痛对大鼠认知功能障碍的影响及其机制。方法选择SD大鼠80只,采用随机数字表法分为模型组、对照组,每组40只。模型组右后足按Brennan法制作趾部切口疼痛模型,对照组仅麻醉,趾部不制作疼痛模型。模型组术后1(t1)、3(t2)、7(t3)、14天(t4),对照组同时间,各随机取10只,分别检测右后足机械缩足阈值(MWT)、热缩足潜伏期(TWL),通过Morris水迷宫实验检测穿越平台次数、平台停留时间。检测完上述指标,断头处死,取海马区脑组织,采用Western blotting法检测突触后膜致密蛋白95(PSD95)、N- 甲基- D- 天冬氨酸受体2B亚型(NR2B)表达。结果模型组t1~t4MWT、TWL均低于对照组同期,穿越平台次数、平台停留时间均低于对照组同期,海马区PSD95、NR2B相对表达量均低于对照组同期(P均<0.05)。结论术后疼痛可导致大鼠认知功能障碍发生,其机制可能与疼痛刺激使海马区PSD95、NR2B表达降低有关。
认知功能障碍;疼痛;海马区;突触后膜致密蛋白95;N- 甲基- D- 天冬氨酸受体2B亚型;大鼠
术后认知功能障碍是患者全麻术后出现的一种中枢神经系统并发症,多发生于老年人[1]。其发病机制至今仍不十分清楚。研究发现,人脑组织海马区存在N- 甲基- D- 天冬氨酸(NMDA)受体,该受体可调节海马神经元发育和突触形成,在学习、记忆和认知过程中具有重要作用[2,3]。突触后膜致密蛋白95(PSD95)可结合NMDA受体2B亚型(NR2B)形成突触后膜蛋白复合物,在海马回的突触重塑过程中发挥重要作用[4]。2016年9月~2017年2月,本研究观察术后疼痛对大鼠认知功能障碍的影响,并探讨其可能机制。
1 材料与方法
1.1 材料 健康雄性SD大鼠80只,SPF级,18月龄,体质量(400±30)g,购自斯贝福(北京)生物技术有限公司,动物许可证号:SCXK(京)2016- 0002。所有大鼠单笼饲养,自由摄食饮水,室温22~25 ℃,湿度55%~65%,12 h昼夜交替。Morris水迷宫系统,上海欣软信息科技有限公司;电泳仪、转膜仪,北京六一仪器厂;PL- 200热刺痛仪,成都泰盟科技有限公司;Von Frey电子测痛仪,北京中昊万通科技有限公司;酶标仪,上海一恒科技有限公司。BCA蛋白浓度测定试剂盒,北京碧云天生物科技有限公司;鼠抗PSD95、兔抗NR2B一抗,美国Sigma公司;鼠抗β- actin一抗、HRP标记的山羊抗鼠及山羊抗兔二抗,北京中杉金桥生物技术有限公司。
1.2 动物分组及模型制备 所有大鼠适应性喂养1周,采用随机数字表法分为模型组、对照组,每组40只。模型组右后足按Brennan法制作趾部切口疼痛模型。具体步骤:大鼠采用20%水合氯醛400 mg/kg腹腔注射麻醉,右后肢备皮,安尔碘消毒,铺无菌洞巾,用15号刀片从足跟近端0.5 cm处向趾部作一长约1.0 cm的纵向切口,尽量避开血管以减少出血,逐层切开皮肤,分离筋膜和肌肉,用镊子挑起足底肌肉,纵向切开,纱布按压减少出血,用0号丝线缝合肌肉和皮肤,建模完成。整个手术过程不超过5 min。对照组同法麻醉,趾部不制作疼痛模型。两组均于温暖环境中自然苏醒,避免强光和噪音刺激。
1.3 相关指标观察
1.3.1 机械缩足阈值(MWT)和热缩足潜伏期(TWL) 模型组术后1、3、7、14天(分别记为t1、t2、t3、t4),对照组同时间,各随机取10只,检测右后足MWT和TWL。所有大鼠置于Von Frey电子测痛仪或PL- 200热刺痛仪上适应20 min,并排除光线、噪音、环境等干扰因素。MWT检测:待大鼠安静后,用Von Frey电子测痛仪的刺激针头从筛网下方垂直刺激术侧后爪中部,逐渐增加刺激强度,最大不超过40 g。大鼠出现逃避性抬足时的数值即为MWT。TWL检测:将PL- 200热刺痛仪置于大鼠术侧足底中部,用红外光近距离照射足底,照射时间最长15 s。从照射开始到大鼠出现逃避性抬腿的时间即为TWL。检测时间为9:00~12:00。每只大鼠重复检测5次,每次刺激间隔5 min,取其平均值。
1.3.2 穿越平台次数及平台停留时间 采用Morris水迷宫实验。Morris水迷宫泳池分为四个象限,平台位于其中一个象限中央。两组均于术前进行为期5天的获得性训练。将大鼠头朝向池壁轻轻放入水中,记录大鼠水下找到平台的时间,并让其在平台上停留30 s;若60 s内未能找到平台,引导其上平台并在平台停留30 s。每天训练4次,每次间隔大于30 min。两组检测完MWT、TWL后进行探查训练。撤除Morris水迷宫平台,将大鼠从原入水点的对侧放入水中,并通过摄像系统记录60 s内大鼠游过目标象限(原平台所在象限)的次数(即穿越平台次数)及时间(平台所在象限停留时间)。
1.3.3 海马区PSD95、NR2B表达 采用Western blotting法。各组完成Morris水迷宫实验后,20%水合氯醛400 mg/kg腹腔注射麻醉,断头处死,迅速剥离枕骨和顶骨,完整取海马区脑组织,- 80 ℃冰箱保存。所有脑组织收集完毕后置于蛋白裂解液和蛋白酶抑制剂中,电动匀浆,室温裂解30 min,4 ℃、13 000×g离心20 min,取上清液,BCA法进行蛋白定量后,加入5×上样缓冲液,旋涡混匀,沸水浴变性10 min。取上样蛋白20 μg,行SDS- 聚丙烯酰胺凝胶电泳,当目的蛋白跑至凝胶底部时停止电泳,并转印至PVDF膜;封闭液封闭,然后分别加入鼠抗PSD95(1∶500)、兔抗NR2B(1∶500)、鼠抗β- actin(1∶2 000)一抗,4 ℃孵育过夜;TBST洗膜10 min×3次,加入 HRP标记的山羊抗鼠(1∶2 000)或山羊抗兔(1∶2 000)二抗,37 ℃孵育1 h;TBST洗膜10 min×3次,ECL发光,暗室显影、定影,Image J软件分析各蛋白电泳条带灰度值。以β- actin内参,以目的蛋白电泳条带灰度值与β- actin蛋白电泳条带灰度值的比值作为目的蛋白相对表达量。

2 结果
2.1 两组MWT、TWL比较 见表1。
2.2 两组穿越平台次数、平台停留时间比较 见表2。
2.3 两组海马区PSD95、NR2B表达比较 见表3。
3 讨论
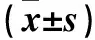
表1 两组各时点MWT、TWL比较
注:与对照组同期比较,*P<0.05;与同组t1时比较,#P<0.05;与同组t2时比较,△P<0.05;与同组t3时比较,▲P<0.05。
表2 两组各时点穿越平台次数、平台停留时间比较
注:与对照组同期比较,*P<0.05;与同组t1时比较,#P<0.05;与同组t2时比较,△P<0.05。
表3 两组各时点海马区PSD95、NR2B 相对表达量比较
注:与对照组同期比较,*P<0.05;与同组t1时比较,#P<0.05;与同组t2时比较,△P<0.05;与同组t3时比较,▲P<0.05。
POCD是全麻术后出现的中枢神经系统并发症,主要表现为精神错乱、焦虑、人格改变以及记忆受损等,是老年人术后常见的并发症之一,可导致患者生活质量下降、住院时间延长、医疗成本增加、病死亡率升高等[5]。随着我国人口老龄化加剧,老年人因各种疾病入院接受全麻手术者逐渐增多,POCD的发生率亦逐渐升高。目前,对POCD产生的原因主要考虑麻醉药物、患者年龄等因素,而对手术本身疼痛刺激的研究较少。
术后持续性疼痛是临床上常见的一种病理性疼痛,一般认为是由伤害感受器持续性兴奋而引发的中枢敏感化而致。研究发现,手术相关的伤害性刺激可促进POCD的发生,伤害性刺激后的痛觉过敏也与POCD密切相关[6,7]。本研究模型组按Brennan法制作大鼠趾部切口疼痛模型,对照组仅麻醉,不作趾部切口疼痛模型,观察两组疼痛敏感性变化;结果发现,模型组术后1~14天MWT、TWL均明显降低,表明趾部切口手术可诱发术后痛觉过敏现象。本研究通过Morris水迷宫实验评价小鼠学习记忆能力,结果发现,在伤害性刺激存续期间,模型组穿越平台次数较对照组明显减少,平台停留时间较对照组明显缩短,说明模型组已经出现了认知功能障碍,且可能与手术伤害性刺激有关。
NMDA受体广泛分布于哺乳动物的中枢神经系统中,并与突触可塑性及神经兴奋性毒性密切相关。NMDA受体分为NR1、NR2和NR3三种亚单位,其中NR2亚单位中的NR2B亚基在中枢痛敏的形成和神经系统的学习记忆突触机制中至关重要[8,9]。手术相关的伤害性刺激能阻断NMDA受体,抑制突触前膜谷氨酸的释放,使大脑神经元发生退行性病变[10]。NR2B亚基在脊髓以上大脑皮层密度最高。有研究表明,存在认知功能障碍的动物可出现与认知相关的脑区皮层重排[11],如内侧前额叶、前扣带回和海马区等,这可能是出现认知功能障碍的基础。海马作为情绪、学习和记忆中枢,在工作记忆、情感记忆及注意力调节等脑功能中发挥重要作用[12]。有研究发现,疼痛模型大鼠的认知功能减退与海马-前额叶皮层的通路结构连接减少有关。Qiu等[13]研究发现,海马区NMDA受体NR2B亚基有助于海马神经元发育和突触形成。PSD95是一种突触后膜标记物,可代表突触的形态和密度。有研究发现,PSD95表达降低与突触数量减少或突触丢失以及学习和记忆功能损害有关[14]。NMDA受体活性受PSD95蛋白家族调节,PSD95本身没有蛋白酶活性,但可结合NMDA受体亚基NR2B,形成PSD95/NR2B复合物,参与NMDA受体介导的突触信号传递和突触可塑性调节等[15]。阚红军等[16]研究发现,神经病理性疼痛可引起依赖于NMDA受体的PSD95重新分布,而大鼠出现空间记忆障碍与内侧前额叶部位突触小体PSD95的重塑有关。本研究模型组海马区各时间点PSD95、NR2B相对表达量均低于对照组,说明术后疼痛刺激能够抑制海马区突触重塑,且可阻断NMDA受体,抑制突触前膜谷氨酸释放,增加神经兴奋性毒性,与上述观点基本一致。
综上所述,术后疼痛可致大鼠认知功能障碍发生,其机制可能与疼痛刺激使海马区PSD95、NR2B表达降低有关。本研究为海马区PSD95、NR2B有可能作为术后认知功能障碍的治疗靶点提供了理论基础。
[1] Cao XZ, Ma H, Wang JK, et al. Postoperative cognitive deficits and neuroinflammation in the hippocampus triggered by surgical trauma are exacerbated in aged rats[J]. Prog Neuropsychopharmacol Biol Psychiatry, 2010,34(8):1426- 1432.
[2] Zhang X, Xin X, Dong Y, et al. Surgical incision- induced nociception causes cognitive impairment and reduction in synaptic NMDA receptor 2B in mice[J]. J Neurosci, 2013,33(45):17737- 17748.
[3] Liu L, Wong TP, Pozza MF, et al. Role of NMDA receptor subtypes in governing the direction of hippocampal synaptic plasticity[J]. Science, 2004,304(5673):1021- 1024.
[4] Moore CL, Nelson PL, Parelkar NK, et al. Protein kinase A- phosphorylated KV1 channels in PSD95 signaling complex contribute to the resting membrane potential and diameter of cerebral arteries[J]. Circ Res, 2014,114(8):1258- 1267.
[5] Pais- Vieira M, Mendes- Pinto MM, Lima D, et al. Cognitive impairment of prefrontal- dependent decision- making in rats after the onset of chronic pain[J]. Neuroscience, 2009,161(3):671- 679.
[6] Landro NI, Fors EA, Vapenstad LL, et al. The extent of neurocognitive dysfunction in a multidisciplinary pain centre population. Is there a relation between reported and tested neuropsychological functioning[J]. Pain, 2013,154(7):972- 977.
[7] Moriarty O, McGuire BE, Finn DP. The effect of pain on cognitive function: a review of clinical and preclinical research[J]. Prog Neurobiol, 2011,93(3):385- 404.
[8] 龚琴,胡兴国,刘艳阳,等.脊髓背角p38 MAPK活化在大鼠切口痛形成中的作用[J].中国疼痛医学杂志,2009,15(4):226- 229.
[9] Vorhees CV, Williams MT. Assessing spatial leaming and memory in rodents[J]. ILAR J, 2014,55(2):310- 332.
[10] Nona CN, Li R, Nobrega JN. Altered NMDA receptor subunit gene expression in brains of mice showing high vs. low sensitization to ethanol[J]. Behav Brain Res, 2014(260):58- 66.
[11] Xiao C, Huang Y, Dong M, et al. NR2B- selective conantokin peptide inhibitors of the NMDA receptor display enhanced antinociceptive properties compared to non- selective conantokins[J]. Neuropeptides, 2008,42(5):601- 609.
[12] Bustos FJ, Varela- Nallar L, Campos M, et al. PSD95 suppresses dendritic arbor development in mature hippocampal neurons by occluding the clustering of NR2B- NMDA receptors[J]. PLoS One, 2014,9(4):e94037.
[13] Qiu S, Li XY, Zhuo M. Post- translational modification of NMDA receptor GluN2B subunit and its roles in chronic pain and memory[J]. Semin Cell Dev Biol, 2011,22(5):521- 529.
[14] Seo MK, Lee CH, Cho HY, et al. Effects of antidepressant drugs on synaptic protein levels and dendritic outgrowth in hippocampal neuronal cultures[J]. Neuropharmacology, 2014(79):222- 233.
[15] Liao GY, Wagner DA, Hsu MH, et al. Evidence for direct protein kinase- C mediated modulation of N- methyl- D- aspartate receptor current[J]. Mol Pharmacol, 2001,59(5):960- 964.
[16] 阚红军,于剑锋.神经病理性疼痛引起大鼠空间记忆障碍和内侧前额叶PSD95以及CaMK Ⅱ- Thr305的表达升高[J].中国疼痛医学杂志,2015,21(3):182- 188.
10.3969/j.issn.1002- 266X.2017.36.013
R749.1
A
1002- 266X(2017)36- 0043- 03
2016- 09- 05)
