人工苗圃鹭类繁殖集群研究
2017-11-16韦定菊赵俊松姚永芳
韦定菊 赵俊松 李 波 姚永芳
(1.四川农业大学动物科技学院,成都,611130;2.四川农业大学生命科学院,雅安,625014)稿件运行过程
人工苗圃鹭类繁殖集群研究
韦定菊1,2赵俊松1,2李 波2姚永芳1*
(1.四川农业大学动物科技学院,成都,611130;2.四川农业大学生命科学院,雅安,625014)稿件运行过程
广汉;人工苗圃;鹭类;繁殖集群
2014年4~6月,采用直接观察法对广汉市南丰镇双庙子村村头的一块人工苗圃内繁殖的4种鹭科鸟类(简称鹭类)(牛背鹭Bubulcusibis,白鹭Egrettagarzetta,池鹭Ardeolabacchus,夜鹭Nycticoraxnycticorax)的繁殖集群进行了监测,对鹭类鸟巢的分布密度、种群结构以及鹭类鸟巢的垂直分布和鸟巢大小进行了系统研究与分析。据统计,该苗圃有鹭类鸟巢1 580个,其中牛背鹭类巢(653,41.31%)为最多,高巢位的是夜鹭类巢。树木的高度和林木郁闭度是影响鹭类巢址选择的主要因素,而林地面积对鹭类巢址的选择影响不明显。影响鹭类巢穴大小的主要因素是鹭类本身体型、树木高度和在巢穴中的居住时间,但鸟巢中新增的幼鸟数量对巢穴大小无影响。
苗圃是培植植物幼株(苗木)的场所,它的主要任务是为人工林建设和城乡绿化提供优质苗木[1]。大型的苗圃除了具有种植苗木,获取一定的经济报酬之外,还具有保护环境,提高环境质量,保护水土和庇护动物等多种生态功能[2-3]。鹭类适合在稻田或其他涉水地带生活、取食,且对环境的变化具有高度的敏感性,因此通常在评价湿地生态系统时,常选用鹭类作为指示性物种[4-5]。靠近农田、湿地等的苗圃,往往可能成为鹭类栖息地的选择对象之一。
鹭类在中国是广布种,为晚成鸟,也是典型的林栖集群繁殖鸟类[6],朱红星[7]曾对6种鹭类混群营巢的情况进行了调查,发现浅色系的鹭类通常占据营巢地中心地位,而有色和深色系的通常分布在周边,这一混群情况与叶芬等[8]在进行广西防城7种鹭类混群繁殖的空间生态位研究以及谭飞等[9]对福建漳江口鹭类的巢址选择研究结果相类似;但是朱红星等[7]在研究中观察到混群的鹭类中牛背鹭居于高位鸟巢,而叶芬等[8]和谭飞等[9]观测到的现象与朱曦等[10]一致,白鹭(Egrettagarzetta)和夜鹭(Nycticoraxnycticorax)居于高巢位。陈旭健等[11]和王博等[12]分别在厦门和广西均观测到大部分鹭类在3月进入繁殖期,周立志等[13]报道合肥鹭类在4月底进入繁殖期。赵俊松等[14]研究云南大理古镇的3种鹭类在2月进入繁殖期,5月底鹭类离巢。由此可见,鹭类的繁殖集群方式不是单一的局限于某一地域、气候以及降水等条件,并且集群繁殖情况也不仅仅只局限于一种形式,而是存在着3~7种的混群方式[7-14]。鸟类的集群繁殖可以使其降低被捕食的风险、共享繁殖地以及交换信息[15-16]。多个鹭类集群繁殖地的研究发现,鹭类进入同一繁殖栖息地的时间存在非同步性,例如:最先迁徙到紫蓬山区的是白鹭,接着是夜鹭,最后池鹭(Ardeolabacchus)[13]。为减少物种之间的竞争压力,鹭类通过将鸟巢修筑在同一地点不同空间上[17]。
目前,利用苗圃作为鹭类繁殖地的相关研究尚未见报道。该苗圃距离广汉市鸭子河湿地公园5 km,周围环绕皆是农田,因此吸引了大群鹭类在此栖息。根据当地村民反映,鹭类在该地繁殖栖息已有18 a之久。为此我们对其进行了为期3 a的连续监测,意在探究该苗圃鹭类的集群繁殖机制。
1 研究方法
1.1 苗圃概况
本研究苗圃位于广汉市区城郊(N104°15′21″,E30°02′57″),占地面积2 hm2,周围是大片农田,并且园区内有一条宽约3 m的河贯穿中央(图1)。该地属于亚热带湿润气候区,年降水量900 mm左右,年均约气温16℃,年平均日照时数1 200 h,无霜期300 d左右。园内有银杏(Ginkgobiloba)、樟(Cinnamomumcamphora)、天竺葵(Pelargoniumhortorum)、桂花(Osmanthusfragrans)、楠木(Phoebezhennan)、紫薇(Lagerstroemiaindica)、腊梅(Chimonanthuspraecox)等20种乔木和灌木,为在此栖息的鹭类提供了营巢、栖息环境;其中银杏树、桂花树和樟树在园内形成3片同质林,分别占据3块较大面积,其覆盖面积约为苗圃的三分之二。且近几年人为的干预保护,在6月中旬农民给稻谷喷洒农药期间,以及冬季食物匮乏期,苗圃工作人员则会进行人工投食,且在其栖息地周围建立围栏,使人类活动的干扰得以降低,该生境为鹭类的栖息繁殖提供了充足的食物资源和安全的筑巢环境。
1.2 研究方法
1.2.1 数量监测
2012年9月~2015年6月监测4种鹭类数量的变化,分别于每年的4~6月、9~10月,每个星期监测1次,每次持续进行3 d。其他月份每月1~2次,每次2~3 d,共计57次156 d。运用18×42倍的尼康望远镜、佳能70 D相机以及佳能EF28-300 mm f /3.5-5.6L IS USM的长焦镜头进行鸟类的观察记录,运用直接计数法统计鹭类鸟巢的数量,参考《四川鸟类原色图鉴》[18]进行鹭类种类的鉴定。
2012年9月~2013年9月为预观测期,在这一时间段中发现鹭科鸟类在每年6月中达到繁殖高峰期,数量不再出现大的波动,鹭类育雏时间通常为1个月左右,与文祯中等[19],刘华全[20]和周立志等[13]研究中的鹭类育雏时间相近,自雏鸟孵化后的1~6周,雏鸟通常与成鸟合巢而居,待其具备一定的飞行能力,即6周后(8月下旬),雏鸟开始离巢。2014年6~8月为鹭类集群繁殖观察期,我们运用单、双筒望远镜和单反相机相结合的方法观察统计每个鸟巢,统计成鸟与幼鸟的总数。运用求平均数的方法计算单位鸟巢幼鸟数量。2014年9月~2015年为鹭类数量监测期,主要通过双筒望远镜和单反相机监测鹭类的数量。
1.2.2 样方设计
2013年9月~2014年6月运用样方法在园区内进行调查。苗圃内种植着人工林,树木之间的距离比较整齐,并且鹭类主要聚集在银杏树、桂花树和香樟树修筑鸟巢,根据苗圃内的园林道路设计和不同的植被类型,我们把有鸟巢的苗圃依树种划分为3块(图1),其中鸟巢聚集地1的面积是0.56 hm2,主要植被是杏树;鸟巢聚集地2是0.08 hm2,主要植被是樟树;鸟巢聚集地3是0.05 hm2,主要植被是桂花树,样方面积为6 m×6 m。在白鹭园内用25 m长的皮尺量出16块样方,其中鸟巢聚集地1选取8块样方,鸟巢聚集地2、3各4块样方,总面积占整个鸟巢聚集地的8.18%,利用AS600电子测距仪逐个测量并记录每块样方内有鸟巢的树木高度、鹭类鸟巢离地高度,用2 m的皮尺测量有鸟巢树木的胸径,通过望远镜和单反相机观察并记录鸟巢的类型、估测鸟巢大小。用望远镜观察鸟巢在林内的分布和混群情况。运用望远镜、单反相机和目测计数法相结合记录鸟巢数量。

图1 苗圃概况Fig.1 The sketch map of the nursery
1.3 数据处理
1.3.1 Pielou均匀性指数[21]
J=H/Hmax
J为均匀性指数,H为实测多样性值,Hmax=log2S,S为群落鸟类种数。
1.3.2 种群分布型
鹭科鸟类种群分布型,采用方差/平均数,即S2/m来判断。S2/m=0:均匀分布;S2/m=1:随机分布;S2/m明显>1:成群分布。
1.3.3 物种生态位宽度[22]
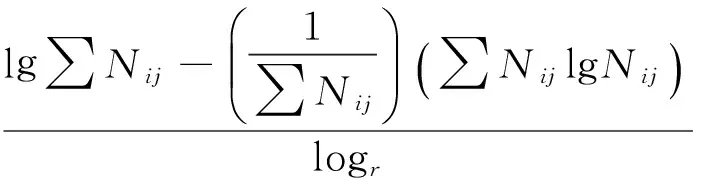
r为资源位数目,Nij为i种鸟类利用j资源位的数值。
1.3.4 生态位重叠

Pxi、Pyi为物种x、y对资源位i的利用比例,Cxy的变化范围为0(无重叠)~1(完全重叠)
2 研究结果
2.1 鹭类鸟巢数量
2014年4~6月,直接计数统计苗圃内总鸟巢数
1 580个,鸟巢聚集地1、2、3的16个样方中,分别有鹭类鸟巢101、35、77个(表1)。根据样方中鹭类鸟巢比例(样方中每个物种量/样本总数),牛背鹭(Bubulcusibis)、白鹭、夜鹭和池鹭分别是41.31%、33.8%、21.6%、3.29%,因该苗圃鸟巢总数为1 580,因此推测苗圃内共有牛背鹭类巢653个、白鹭类巢534个、夜鹭类巢341个、池鹭类巢52个。
2.2 群体数量
6月中旬鹭类繁殖进入末期,鹭类数量趋于顶峰,数量波动趋于平稳,且幼鸟与成年鹭类同居,易于统计单位鸟巢鹭类。据调查,在苗圃内,白鹭的每巢鹭只数为4.21只(n=24,4.21±1.76)、牛背鹭为3.73只(n=37,3.73±1.69)、夜鹭为3.21只(n=29,3.21±1.32)、池鹭为2.72只(n=22,2.72±1.08)(表2)。结合各种鹭类鸟巢个数,计算出白鹭园内白鹭2 243只,牛背鹭2 416只,夜鹭1 092只,池鹭140只,鹭科鸟类总计5 891只(表2)。
2.3 鸟巢的垂直分布
利用测距仪测量鸟巢聚集地1、2、3的平均树高分别为13.17 m(n=79,13.17±2.577)、12.73 m(n=28,12.73±1.728)、8.81 m(n=27,8.81±2.127),由于树木的高度导致了鸟巢高度分布的差异性,据此将鹭类鸟巢高度分为5个等级(表3)。
不同种类的鹭科鸟类中,白鹭是广布种,鸟巢高度在划分的5个高度等级之间均有分布,并且每个等级的鸟巢几乎均匀分布于各个聚集地中,受树高树种的影响最小,因此生态位宽度最大;夜鹭类巢偏高,在最低高度等级没有分布,但是较低高度等级的鸟巢只分布在鸟巢聚集地3,而较高高度等级的鸟巢只分布在鸟巢聚集地1,呈现出很明显的鸟巢高度与树高的正相关性;牛背鹭的鸟巢分布同夜鹭相似;池鹭类巢高度是4种鹭类中最低的,只有分布在I级区间内,且白鹭园树木的高度对其鸟巢高度无明显影响,它的鸟巢高度仅局限于7~9 m之间,因此生态位宽度极小。
表1 4种鹭类16个样方中鸟巢的统计量

Tab.1 The statistical data of the nests for 4 kinds of egrets in 16 sampling quadrats

续表1
表2 白鹭园内繁殖群体数量

Tab.2 Egret Park community composition
表3 白鹭园鹭科鸟类鸟巢的垂直分布

Tab.3 The vertical distribution of birds nests
注:a:鸟巢聚集地1鸟巢数;b:鸟巢聚集地2鸟巢数;c鸟巢聚集地3鸟巢数
Note :a:number 1 aggregation of nests;b:number 2 aggregation of nests;c:number 3 aggregation of nests
不同高度等级的鸟巢中,根据鸟巢数与该种鹭类总数的比例发现,池鹭在Ⅰ级鸟巢中数量占据了绝对优势,所有的池鹭类巢均分布在这一高度组上;夜鹭在Ⅲ、Ⅴ组中的数量占优势,数量优势的鸟巢分布地分别是在聚集地3和1。树木平均高度最高的鸟巢聚集地1中分布着所有高度等级的鸟巢,并且Ⅴ和Ⅳ这2个等级的鸟巢占了50%,而在树木高度较低的鸟巢聚集地3分布的鸟巢偏低,83%的鸟巢是在低等级的Ⅰ和Ⅱ。
根据4种鹭类的垂直分布情况划分出3种混群现象,即:白鹭—池鹭—牛背鹭;白鹭—牛背鹭—夜鹭;白鹭—夜鹭。5个不同的巢穴高度等级中,鸟巢分布均匀性越高,则越适合鹭类生存,11~13 m鸟巢数是74个为最多,也是5个高度等级中最高的,我们认为11~13 m这一高度是最适宜鹭类居住的高度;反之在15~17 m鸟巢数为11个,是所有高度等级中,数量最少的,在15~17这一高度上,并不适大部分的鹭鸟筑巢。
表4 白鹭园4种鹭类鸟巢垂直生态位重叠

Tab.4 4 kinds of egrets nests the vertical of niche overlap
白鹭在3个混群中皆存在,并且与牛背鹭和夜鹭有两种混群现象,所以白鹭与这两种鹭类的垂直生态位重叠率较高,分别是0.82和0.85(表4);池鹭和牛背鹭存在一种混群现象,因此这两种鹭类的垂直生态位重叠率较低,是0.16;池鹭与夜鹭不存在混群现象,在垂直生态位上无重叠,牛背鹭和夜鹭存在一种混群现象,但是混群中两个物种的数量较多,生态位重叠率是0.7。据此数据可知,物种之间交集越多,数量越大,生态位重叠率越高,反之重叠率越低甚至无重叠。
2.4 鹭类的鸟巢大小
鹭类的鸟巢是由枝条或者带有树叶的枝条筑造较大凌乱的浅巢,最简易的鸟巢由十几根无叶树枝搭建,最大是由带树叶的枝条搭建的外部体积5 L以上的鸟巢(外部长和宽350 mm以上,外部鸟巢高度200 mm以上),鸟巢的大小易用望远镜放大区分。将鹭类的鸟巢大小划分了5个等级(表5)。Ⅲ级鸟巢数量在各个等级中居首位,是鹭类的优势鸟巢。不同种类的鹭类占据不同的优势鸟巢:白鹭和牛背鹭优势鸟巢是Ⅲ级;夜鹭是Ⅳ级;池鹭是Ⅱ级。进一步分析发现白鹭Ⅲ、Ⅳ级鸟巢数量之和比Ⅰ、Ⅱ级之和鸟巢多18个,而牛背鹭巢多8个,白鹭的大型鸟巢比牛背鹭多。因此推断这4种鹭类鸟巢大小的关系是:夜鹭>白鹭>牛背鹭>池鹭。优势鸟巢主要分布于鸟巢聚集地1,由此推断鸟巢聚集地1是鹭类巢址优势地。并且鸟巢聚集地1作为优势地现象明显,优势鸟巢是Ⅲ,且各等级鸟巢均有分布;而其余2个鸟巢聚集地皆有不同程度的某一等级不分布的现象,鸟巢聚集地2优势鸟巢是Ⅲ级,不分布Ⅰ和Ⅴ级鸟巢;鸟巢聚集地3优势鸟巢是Ⅱ,无Ⅴ级鸟巢。
表5 鹭科鸟类鸟巢大小

Tab.5 Birds nests size
注:Ⅰ级:十几根树枝搭建;Ⅱ级:体积:1~2 L,长宽为150~250 mm,高50~100 mm;Ⅲ级:2~4 L,长宽为250~350 mm,高50~100 mm;Ⅳ级:4~5 L,长宽为250~450 mm,高100~150 mm;Ⅴ级:5 L以上,长宽约为350 mm以上,高200 mm以上
Note:Ⅰ levels:as a dozen or tree branches build;Ⅱ levels:Volume:1-2 L,length and width:150-250 mm,high:50-100 mm;Ⅲ levels:Volume:2-4 L,length and width:250-350 mm,high:50-100 mm;Ⅵ levels:Volume 4-5 L,length and width:250-450 mm,height:100-150 mm;Ⅴ levels:Volume above 5 L,length and width:above 350 mm,high above:200 mm
3 讨论
3.1 鹭类鸟巢数量和密度分析
苗圃环境稳定,外界干扰相对较少,鹭类对其集群繁殖地具有高度的依恋性,通常会连续多年使用[23],鹭类每年都有重复使用旧巢的习惯,除环境波动及干扰较严重情况下一般不会放弃原住处[24],为此可以认为鹭类集群繁殖观察期的数据具有代表性。
种群数量决定了鸟巢数量,鸟巢数量影响着鸟巢密度。苗圃中鹭类鸟巢密度并非均匀分布,除面积越大,鹭类鸟巢分布越稀疏外,鹭类并非对所有生境组成部分全盘接收,而是仅选择对其有利的区域中修筑鸟巢,其有利因素包括:(1)树木高度,苗圃中所有的鹭类巢穴均高于7 m,且鹭类的鸟巢多位于11~15 m处,由此可知高于7 m的树木,且高11~15 m的树木为鹭类优先选择的树木,而低于7 m,远高于15 m的树木,被鹭类选择用于鸟巢修筑地的可能性很低,由此可知,鹭类仅筑鸟巢于特定高度的树木上。(2)苗圃树木密闭,树木枝干可为鹭类提供鸟巢安置支撑点,树叶可为鹭类提供遮挡,有利于鹭类躲避天敌以及在炎热的夏天躲避烈日,而分枝少,树叶稀疏的树木被鹭类选为巢址地的可能性低。(3)觅食,水稻田的分布对鹭类繁殖地有影响[25]。鹭类筑巢地靠近农田,除方便鹭类觅食外,也避免了鹭类进行较长的取食距离飞行,即减少了觅食消耗过多的体力,且在繁殖期,就近觅食,可使得成年鹭类更有效地育雏,并可降低幼雏的死亡率,所以,远离水稻田(取食地)的区域没有鹭类的筑巢。(4)人为干扰,影响鹭类栖息的最大因素是人类活动,苗圃内有工作人员工作区,区内又修筑有水泥小道,这里人为活动频繁,鹭类为更好地保护自己,很少在这些地方停息,更少筑巢,而距离人活动较少的林地中央,以及林地偏僻处,鹭类鸟巢较为密集。
4种鹭类的数量之间存在很大的差异性,牛背鹭巢穴653个,而池鹭只有52个,这一数量的差异与各种鹭类在该地的生活状态有关,除了池鹭外,其他3种鹭类均全年生活在苗圃中,居住时间对种群数量有积极影响,在有限面积的苗圃中,种群数量的增加必然引起鸟巢密集度的增加,且留鸟比候鸟更优先选择最有利的巢址。这一结果与朱红星等[7]研究的结论相一致。
3.2 群落组成分析
研究结果显示,随着巢址面积的增加,单位面积鹭类数呈现递减现象,因在同一片苗圃中,鹭类对巢址地也有选择性。巢址选择影响鸟类的生存和繁殖能力,并且随着栖息环境的不同,同种鸟类对巢址的选择会有所变化[6]。鹭类的营巢地有针阔混交林、杉树林、针叶混交林、毛竹林等多种生境类型,生境的不同,巢址选择树种也不同[10,26-27]。在白鹭园中,鹭类的巢址地优先考虑的是较高树木,且栽种面积最大的杏树林即鸟巢聚集地1,其次是鸟巢聚集地2,最后是鸟巢聚集地3,而苗圃中的其他区域是不被考虑的。
3.3 鹭类垂直分布分析
鹭科鸟类垂直分布现象无论是从不同鹭科鸟类、鸟巢不同高度等级或者是不同巢址地中的结果都显示出与李建国等[28-31]报道结果类似,即高位鸟巢鹭类为大体型鹭类,反之低位鸟巢鹭类的体型小。夜鹭在4种鹭类中体型和重量最大[6],居于最高巢位,而池鹭在4种鹭类中体型最小,巢位最低。这一现象验证Burger[13,32]研究所得巢位与身体长度之间具有正相关关系,大个体物种巢位高,小个体物种巢位低的结论相一致;同时也印证了Price[33]所报道的生态上近似的物种之间采取的平衡生存策略。
3.4 影响鹭科鸟类鸟巢大小的因素
鹭类的鸟巢大小有明显的差别,而有关这方面的报道很少。在Zhang等[34]的研究中,夜鹭类巢体积最大,池鹭体积最小,但是几种混群鹭类的鸟巢体积大小差距不大,相对均匀。
根据种群的居留分布显示,夜鹭是苗圃中的常见留鸟,一年四季数量波动不明显,为满足长期居住的需求,长时间不间断地修筑鸟巢,因此鸟巢更饱满、坚固;白鹭和牛背鹭大部分时间数量较稳定,但是除繁殖季节外的其余时间,数量相对波动较大,鸟巢体积小于夜鹭;池鹭是旅鸟,只有繁殖期见于白鹭园,其鸟巢相对较小且简单。由此可知,鹭科鸟类在苗圃的生活时间影响鸟巢体积,生活时间越长越恒定,鸟巢体积越大,反之鸟巢体积小。
根据苗圃面积与鹭类鸟巢大小研究发现,1、2、3鸟巢聚集地的树高和面积依次递减,虽鸟巢聚集地1和2优势鸟巢均为Ⅲ,进一步分析,鸟巢聚集地2鸟巢大部分小于鸟巢聚集地1。但鸟巢聚集地1的树木郁闭度低于鸟巢聚集地2、3,而2、3的巢穴体积均小于1,为此难以认定树木高度以及林地面积与鹭类巢穴的关系。这现象表明苗圃树木高度、面积对鹭类鸟巢体积影响是不明显的。
本调查发现,幼雏刚出生前6周与成鸟共处一个鸟巢中,6周后开始单独生活,由于这个时间比较短,且在此期间幼雏体型不大,原有的2只成鸟生活的鸟巢可以供它们共同使用,且在育雏期间,成鸟主要任务是哺育幼鸟,无过多的时间去修筑鸟巢,且实际统计的数据也未出现明显的单位鸟巢鹭类数与鸟巢体积的关系,因此鸟巢体积不受单位鸟巢鹭类数量增加的影响。
致谢:在广汉市白鹭园鹭鸟集群现象调查过程中,得到白鹭园主人廖全福大力支持,在此一并致谢!
[1] 陈俊愉,戴思兰.几种菊属植物的引种研究[J].北京园林,1996(2):17-22.
[2] 孙胜利.绍兴“通缉”“加拿大一枝黄花” [J].中国花卉园艺,2004 (14):11.
[3] 俞益武,张建国,崔会平.观赏果树的概念、特征与功能[J].福建林业科技,2007,34(3):199-201,233.
[4] 赵洪峰,雷富民.鸟类用于环境监测的意义及研究进展[J].动物学杂志,2002,37(6):74-78.
[5] 约翰·马敬能,卡伦·菲利普斯,何芬奇,等.中国鸟类野外手册[M].长沙:湖南教育出版社,2000.
[6] 文祯中,王庆林,孙儒泳.鹭科鸟类种间关系的研究[J].生态学杂志,1998,17(1):27-34.
[7] 朱红星,聂继山,赵耀宗.六种鹭鸟混群营巢地的调查[J].四川动物,1994,13(3):123-124.
[8] 叶芬,黄乘明,李汉华.广西防城7种鹭类混群繁殖的空间生态位研究[J].四川动物,2006,25(3):577-583.
[9] 谭飞,吴秋城,林英华,等.福建漳江口红树林鹭科鸟类巢址选择[J].动物学杂志,2013,48(2):212-219.
[10] 朱曦,杨士德,邹小平,等.夜鹭繁殖习性与生长发育研究[J].动物学研究,2000,21(1):58-64.
[11] 陈旭健.广西博白县白鹭岛鹭科鸟类资源保护[J].玉林师范学院学报:自然科学版,2003,24(4):94-96.
[12] 王博,陈小麟,林清贤,等.厦门鹭类集群营巢地分布及其生境特性的研究[J].厦门大学学报:自然科学版,2005,44 (5):734-737.
[13] 周立志,宋榆钧,马勇.紫蓬山区三种鹭繁殖生物学研究[J].动物学杂志,1998,33(4):34-38.
[14] 赵俊松,李廷野,房以好,等.云南大理古城3种繁殖鹭类的巢空间隔离[J].四川动物,2015,34(3):357-363
[15] Ward P,Zahavi A.The importance of certain assemblages of birds as “information‐centres” for food‐finding[J].Ibis,1973,115(4):517-534.
[16] Roberts D F.The History and geography of human genes[J].Journal of the Royal Anthropological Institute,1996,2(1):165-167.
[17] Burger J.Competition between cattle egrets and native North American herons,egrets,and ibises[J].The Condor,1978,80(1):15-23.
[18] 李桂垣.四川鸟类原色图鉴[M].中国林业出版社,1993.
[19] 文祯中,夏敏.黄嘴白鹭的繁殖生态学研究[J].信阳师范学院学报:自然科学版,1996,9(3):279-286.
[20] 刘华全.三种鹭类繁殖、孵化及雏鸟生长发育的初步研究[D].厦门:厦门大学,2010.
[21] Hurlbert S H.The nonconcept of species diversity:a critique and alternative parameters[J].Ecology,1971,52(4):577-586.
[22] 戈峰.现代生态学[M].北京:科学出版社,2002.
[23] Kelly J P,Etienne K,Strong C,et al.Status,trends,and implications for the conservation of heron and egret nesting colonies in the San Francisco Bay area[J].Waterbirds,2007,30(4):455-478.
[24] Blaker D.Behaviour of the cattle egert ardeol ibis [J].Ostrich-Journal of African Ornithology,1967,40 (3) :75-129.
[25] Tourenq C,Benhamou S,Sadoul N,et al.Spatial relationships between tree-nesting heron colonies and rice fields in the Camargue,France[J].The Auk,2004,121(1):192-202.
[26] 朱曦,马水龙,戴永样,等.池鹭繁殖种群数量、活动规律和生物生产量的研究[J].生态学报,1994,14(1):75-79.
[27] 朱曦,杨士德,邹小平,等.浙江鹭类资源现状及其保护对策[J].动物学杂志,2001,36(4):37-40.
[28] 李建国,余志伟,邓其祥,等.繁殖期中鹭类混合群体的协调与维持[J].野生动物,1985,6 (5):21-24.
[29] 王中裕,薛江楠,史力军,等.鹭科鸟类混群营巢地的调查[J].野生动物,1990,11 (5):22-23,47.
[30] 伍烈,陈小麟,胡慧娟,等.厦门白鹭自然保护区鹭类繁殖的空间分布[J].厦门大学学报:自然科学版,2001,40(4):979-983.
[31] 王辉,董元华,李涛,等.无锡鼋头渚4种鹭类繁殖期的生态学特征研究[J].农村生态环境,2001,17(4):17-21,38.
[32] Burger J.Resource partitioning:nest site selection in mixed species colonies of herons,egrets and ibises[J].American Midland Naturalist,1979,101(1):191-210.
[33] Price P W.昆虫生态学[M].北京:人民教育出版社,1981.
[34] Zhang Longsheng,Liu Zuomo,Zhang Feng.Breeding ecology and biology of four species of egret and heron[J].Acta Ecologica Sinica,1994,14(1):80-83.
Guanghan;Nursery;Egrets;Breeding colony
Ardeidae Breeding Colony in Artificial Nursery of Sichuan
Wei Dingju1,2Zhao Junsong1,2Li Bo2Yao Yongfang1*
(1.Animal Science and Technology,Sichuan Agriculture University,Chengdu,611130,China;2.Biology College,Sichuan Agriculture University,Yaan,625014,China)
We investigated a breeding colony comprised of four species of the family Ardeidea (cattle egretBubulcusibis,little egretEgrettagarzetta,Chinese pond heronArdeolabacchus,and night heronNycticoraxnyrticorax) by direct observation from March to June 2014 in Shuangmiaozi village,Nanfeng township,Guanghan City,China.We calculated nest density,population structure,vertical distribution and size of nest. We recorded 1 580 nests,including cattle egret nests (653,41%).The vertical distributions showed night herons nests in the forest canopy.The height of trees and canopy density were the main factors influencing nest-site selection,but forest area was not a major factor influencing colony selection.Nest size was affected by the body size of spieces,the height of trees,and residence time,but not by number of nestlings.
2016-04-22
修回日期:2017-05-04
发表日期:2017-11-10
Q958.1
A
2310-1490(2017)04-610-07
四川农业大学学科建设双支计划项目(03570346)
韦定菊,女,25岁,硕士研究生;主要从事野生动物遗传资源保护与利用研究。E-mail:594508017@qq.com
*通讯作者:姚永芳,E-mail:yaoyongf@126.com
