CO2气调胁迫下烟草甲谷胱甘肽S-转移酶基因的表达分析
2017-11-14许抗抗丁天波严毅李灿杨文佳
许抗抗,丁天波,严毅,李灿,杨文佳*
(1.贵阳学院生物与环境工程学院/贵州省山地珍稀动物与经济昆虫重点实验室,贵阳550005;2.青岛农业大学植物医学学院,山东青岛266109)
CO2气调胁迫下烟草甲谷胱甘肽S-转移酶基因的表达分析
许抗抗1,丁天波2,严毅1,李灿1,杨文佳1*1
(1.贵阳学院生物与环境工程学院/贵州省山地珍稀动物与经济昆虫重点实验室,贵阳550005;2.青岛农业大学植物医学学院,山东青岛266109)
根据烟草甲(Lasioderma serricorne)转录组数据信息,克隆获得3个谷胱甘肽S-转移酶(glutathione S-transferases,GSTs)基因的cDNA全长序列,同源性比对和系统进化分析表明,这3个基因分别属于GSTs基因Theta、Delta和 Sigma家族,分别命名为 LsGSTt1、LsGSTd1和 LsGSTs1(GenBank登录号:KY549655、KY549656和KY549657),并分析了其在烟草甲不同发育阶段和CO2气调胁迫后的表达特征,为研究烟草甲GSTs基因的生物学功能提供依据。实时荧光定量聚合酶链式反应(quantitative real-time polymerase chain reaction,qRT-PCR)结果显示:烟草甲LsGSTt1、LsGSTd1和LsGSTs1基因在不同发育阶段均有表达,且在低龄幼虫中表达量最高,显著高于其他时期;经LC10、LC30和LC503种CO2浓度处理低龄幼虫6 h后,烟草甲LsGSTd1表达量相对于对照组无明显变化,而LsGSTt1和LsGSTs1表达量显著高于对照组,推测这2个基因可能主要参与了烟草甲对CO2气调胁迫的应激响应。
谷胱甘肽S-转移酶;烟草甲;基因克隆;表达模式
谷胱甘肽S-转移酶(glutathione S-transferases,GSTs,EC 2.5.1.18)是一种多功能的超基因家族酶,广泛存在于微生物、植物和动物等生物体内[1]。GSTs作为重要的次级代谢酶,其主要功能是降解内源性和外源性有毒物质[2]和保护细胞免受氧化损伤[3]。GSTs也会参与生物体内其他生理途径,例如细胞内物质的运输和储存[1]、细胞信号通路的调节[4]以及嗅觉调控[5]等。根据其底物特异性、同源性和免疫反应,可将昆虫GSTs分为7个家族,即Delta、Epsilon、Omega、Sigma、Theta和Zeta 6个已知家族及未知家族[6]。目前,已有多种昆虫GSTs基因被克隆鉴定,研究发现,它们在杀虫剂抗性[7-8]、激素合成[9]和抗氧化应激[10]过程中发挥着重要作用。
烟草甲[Lasioderma serricorne(Fabricius)],属鞘翅目(Coleoptera)窃蠹科(Anobiidae),其寄主范围广泛,可危害烟草、粮食、茶叶、中药材等多种储藏物,是一种世界性分布的仓储害虫[11]。烟草甲主要通过幼虫潜居寄主体内进行蛀食,严重影响储藏物的产量和品质,其发生危害具有较强的隐蔽性[12]。目前烟草甲的防治主要以化学药剂熏蒸为主,不仅对食品安全和人类安全存在隐患,而且导致烟草甲对杀虫剂产生了不同程度的抗药性,因此,需要探索新的仓储害虫防治策略[13]。CO2气调技术具有安全、有效、无残留等特点,在仓储害虫防治中应用广泛[14-15]。已有研究表明,CO2气调处理可以有效控制烟草甲危害,但不同浓度CO2对烟草甲的毒理作用不同,其中高浓度CO2对烟草甲的控制效果更佳[16]。本课题组前期研究发现,CO2气调处理后烟草甲体内GSTs酶活性显著升高,推测GSTs在害虫应对气调胁迫过程中起着重要作用[17]。目前有关GSTs参与CO2气调胁迫应激响应的分子机制研究较少,因此,鉴定烟草甲GSTs基因并研究其与气调胁迫之间的关系,对于防治仓储害虫具有重要的理论和实践意义。
本研究以转录组测序获得的GSTs序列为基础,应用反转录聚合酶链式反应(reverse transcriptionpolymerase chain reaction,RT-PCR)技术克隆了烟草甲3条不同家族GSTs基因的cDNA全长序列,并对这些基因的分子特性进行了分析。利用实时荧光定量PCR技术分析了烟草甲体内GSTs基因在不同发育阶段和CO2气调胁迫后的表达特性,为研究GSTs基因在烟草甲体内的功能奠定基础,也为阐明昆虫在气调胁迫下的应激机制提供科学依据。
1 材料与方法
1.1 供试虫源
本研究所用烟草甲种群于2010年采自贵州省贵阳市,将其置于温度(28±1)℃、相对湿度(75±5)%、光照14 h、黑暗10 h的人工气候箱内,以中药材当归(Angelica sinensis)为食料连续饲养繁殖40代以上。
1.2 总RNA提取与第1链cDNA合成
按照TRIzol试剂(Invitrogen公司,美国)说明书提取烟草甲成虫的总RNA,利用Nanodrop 2000核酸浓度测定仪(Thermo Scientific公司,美国)和琼脂糖凝胶电泳检测总RNA的浓度和完整性。取1 μg总RNA,利用DNA酶(Promega公司,美国)去除基因组DNA,并按照PrimeScript®RT试剂盒(大连宝生物工程有限公司)说明书合成第1链cDNA,稀释2倍保存于-20℃冰箱备用。
1.3 基因克隆
从贵州省山地珍稀动物与经济昆虫重点实验室构建的烟草甲转录组数据库中获得3个GSTs基因的cDNA序列,经BLAST分析后,采用Primer 5.0软件设计3对全长验证引物(表1)进行开放阅读框的扩增。PCR反应体系为:10×PCR缓冲液2.5 μL,MgCl2(25 mmol/L)2.5 μL,dNTPs(2.5 mmol/L)2 μL,上下游引物(20 μmol/L)各1 μL,cDNA模板1.5 μL,Taq酶(5 U/μL)0.25 μL,加 ddH2O 补至总体积25 μL。反应条件为:95℃预变性3 min,95℃变性30 s,55℃退火30 s,72℃延伸1 min,共35个循环;最后72℃延伸10 min。扩增产物用1%琼脂糖凝胶电泳检测,按照胶回收试剂盒(Gel Extraction Mini Kit,Omega公司,美国)说明书回收目的条带,并连接至pGEM-T Easy载体(Promega公司,美国),再转化到DH5α大肠杆菌感受态细胞中,经蓝白斑筛选和PCR鉴定,将阳性克隆送成都擎科梓熙生物技术有限公司进行测序。

表1 本研究使用的引物Table 1 Primers used in this study
1.4 序列分析
采用DNAMAN 6.03(Lynnon Biosoft公司,美国)对测序结果进行编辑和分析,推导的氨基酸采用BLAST工具(http://www.ncbi.nlm.gov/BLAST/)进行同源性比对分析。利用ProtParam(http://web.expasy.org/)和 NetNGlys 1.0 Server(http://www.cbs.dtu.dk/services/NetNGlyc/)分析编码蛋白的理化性质和N-糖基化位点,并利用SMART程序(http://smart.emblheidelberg.de/)分析结构域。利用MEGA 5.0软件中的邻接法(neighbor-joining)构建系统进化树,各分支均进行1 000次重复检验[18]。
1.5 实时荧光定量PCR检测
分别收集各个发育阶段的烟草甲,包括低龄幼虫、高龄幼虫、蛹和成虫,每个虫态设置4个生物学重复,每个重复40头试虫。参照本实验室CO2气调处理方法[17],取烟草甲低龄幼虫80头放入培养盒,加入40 g当归作为饲料,加盖密封,通过气调混配器,用导管通入不同浓度的CO2气体,待CO2浓度配比达到要求后停止通气。将培养盒进出气口密封后放入人工气候箱培养观察,并统计不同浓度CO2处理后烟草甲的存活与死亡数。根据本课题组前期CO2气调毒理测定结果[16],采用2个亚致死浓度LC10(30%CO2+70%空气)和LC30(70%CO2+30%空气),以及致死中浓度LC50(90%CO2+10%空气)的CO2气调处理试虫,6 h后挑取存活的试虫,以相同条件下的空气处理作为对照,每组处理设置4个重复。按照1.2节方法分别提取所有样品的总RNA并反转录合成cDNA用于实时荧光定量PCR。使用Primer 3.0软件设计特异性表达引物,内参基因采用烟草甲LsEF1α基因(GenBank登录号:KY549658),引物序列信息如表1。实时荧光定量PCR反应体系(20 μL)为:GoTaq®qPCR Master Mix(Promega公司,美国)10 μL、ddH2O 7 μL、cDNA模板1 μL和10 μmol/L上下游引物各1 μL。混匀后轻微离心,反应在CFX96TM实时荧光定量PCR仪(Bio-Rad公司,美国)中进行,反应条件为:95℃预变性5 min;然后95℃变性15 s,60℃退火30 s,72℃延伸30 s,共40个循环;最后运用熔解曲线保证反应的特异性。根据2-ΔΔCT法计算LsGSTt1、LsGSTd1和LsGSTs1基因在烟草甲不同发育阶段和不同浓度CO2气调处理后的相对表达量[19]。
1.6 数据分析
烟草甲不同发育阶段间及CO2气调处理间mRNA表达差异采用单因素方差分析(ANOVA),平均数的多重比对采用邓肯新复极差法(Duncan’s multiple range test,DMRT),显著性水平为p<0.05,数据处理软件采用SPSS 23.0。
2 结果与分析
2.1 LsGSTt1、LsGSTd1和LsGSTs1的克隆及序列分析
根据实验室前期获得的烟草甲转录组数据,利用RT-PCR扩增获得3个GSTs基因的cDNA全长序列,通过同源性比对确定这3个基因分别属于GSTs基因Theta、Delta和Sigma家族,分别命名为LsGSTt1、LsGSTd1和 LsGSTs1(GenBank登 录 号 :KY549655、KY549656和KY549657)。LsGSTt1开放阅读框为564 bp,编码187个氨基酸,预测蛋白质分子质量为22.4 kDa,理论等电点为8.54。根据NetNGlyc 1.0 Server分析结果,LsGSTt1可能存在2个N-糖基化位点,分别为N106和N111。利用SMART程序搜索,发现LsGSTt1具有2个保守结构域:N末端结构域(第4~80位氨基酸)和C末端结构域(第93~181位氨基酸)(图1A)。LsGSTd1开放阅读框为651 bp,编码216个氨基酸,预测蛋白质分子质量为24.5 kDa,理论等电点为5.95。LsGSTd1不存在糖基化位点,其氨基酸序列第3~76位为保守的N末端结构域,第90~207位为C末端结构域(图1B)。LsGSTs1开放阅读框为696 bp,编码231个氨基酸,预测蛋白质分子质量为26.2 kDa,理论等电点为5.06。LsGSTs1不存在糖基化位点,其氨基酸序列第32~101位为保守的N末端结构域,第111~213位为C末端结构域(图1C)。
2.2 LsGSTt1、LsGSTd1和LsGSTs1的序列比对和系统进化分析
将LsGSTt1、LsGSTd1和LsGSTs1编码的氨基酸序列进行BLAST比对。结果发现:LsGSTt1与其他昆虫Theta家族GSTs的同源性较高,其中与墓地甲虫(Nicrophorus vespilloides)(XP_017784965)、赤拟谷盗(Tribolium castaneum)(XP_008196475)和黄粉虫(Tenebrio molitor)(AIL23552)的相似性分别为62%、59%和58%;LsGSTd1与同为鞘翅目的圣甲虫(Oryctes borbonicus)(KRT80357)Delta家族 GSTs的同源性最高,为81%;与赤拟谷盗(T.castaneum)(XP_974273)、黄粉虫(T.molitor)(AIL23531)以及中欧山松大小蠹(Dendroctonus ponderosae)(XP_019755130)的相似性分别为79%、77%和70%;LsGSTs1与其他昆虫Sigma家族GSTs的同源性较高,与墓地甲虫(N.vespilloides)(XP_017775172)、白蜡窄吉丁(Agrilus planipennis)(XP_018334284)和赤拟谷盗(T.castaneum)(XP_967475)的相似性分别为57%、56%和53%。
将上述所得的氨基酸序列与GenBank登录的其他昆虫(赤拟谷盗、黑腹果蝇和家蚕)的GSTs氨基酸序列进行比对,并利用MEGA 5.0中的邻接法构建系统进化树。结果(图2)发现,上述4种昆虫GSTs可分为6 个家族(Delta、Epsilon、Omega、Sigma、Theta和Zeta),本文克隆获得的烟草甲LsGSTt1、LsGSTd1和LsGSTs1基因分别属于Theta、Delta和Sigma家族,且与同为鞘翅目的赤拟谷盗亲缘关系最近。

图1 烟草甲LsGSTs基因的cDNA序列及推导的氨基酸序列Fig.1 cDNA and deduced amino acid sequences of LsGSTs in Lasioderma serricorne

图2 烟草甲和其他昆虫GSTs的系统聚类分析Fig.2 Phylogenetic analysis of GSTs from Lasioderma serricorne and other insects
2.3 LsGSTt1、LsGSTd1和LsGSTs1在不同发育阶段的表达
利用实时荧光定量PCR技术对3个GSTs基因在烟草甲不同发育阶段的相对表达量进行分析。结果(图3)表明:LsGSTt1、LsGSTd1和LsGSTs1基因在烟草甲各个时期均有表达,其中LsGSTt1 mRNA在低龄幼虫、高龄幼虫及蛹中的表达量分别是成虫期的31.5、7.29和4.13倍,且在低龄幼虫期的表达量显著高于其他发育阶段(p<0.05);LsGSTd1 mRNA在低龄幼虫期的表达量分别是高龄幼虫、蛹和成虫的 1.93、2.99和 4.16倍,且差异显著(p<0.05);LsGSTs1 mRNA在低龄幼虫、高龄幼虫及成虫期的相对表达量显著高于蛹期(p<0.05),蛹期的mRNA表达量分别为上述时期的15%、31%和64%。
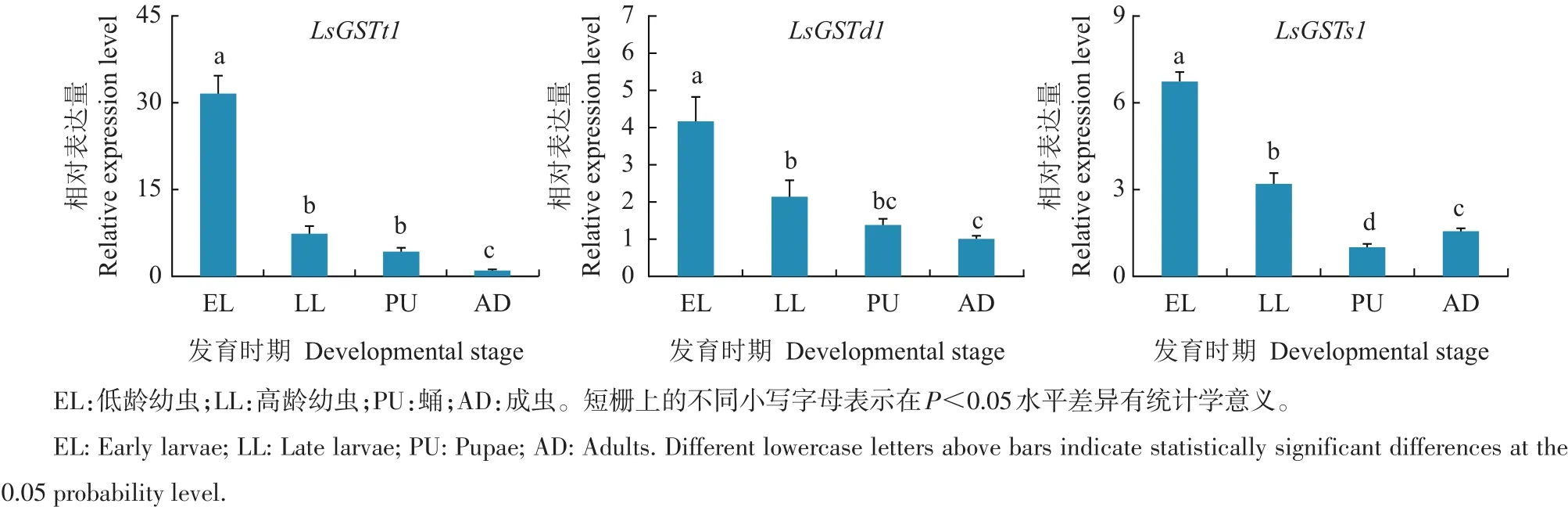
图3 烟草甲不同发育阶段LsGSTs的相对表达量Fig.3 Relative expression levels of LsGSTs in different developmental stages of Lasioderma serricorne
2.4 不同浓度CO2对LsGSTt1、LsGSTd1和LsGSTs1基因表达的影响
采用LC10、LC302个亚致死浓度和致死中浓度LC50的CO2气调胁迫处理烟草甲低龄幼虫。结果(图4)显示:6 h后LsGSTt1基因表达量分别为对照组的2.25、4.64和19.12倍,LC30和LC50处理组的表达量显著高于对照组(p<0.05);CO2处理后LsGSTd1基因表达量与对照组表达量在统计学上差异不显著(P>0.05);LsGSTs1 mRNA的相对表达量分别为对照的1.48、1.06和3.13倍,且在LC50气调胁迫下,mRNA表达量显著高于对照、LC10及LC30处理(p<0.05),但对照、LC10及LC30处理间mRNA相对表达量的差异并不显著。
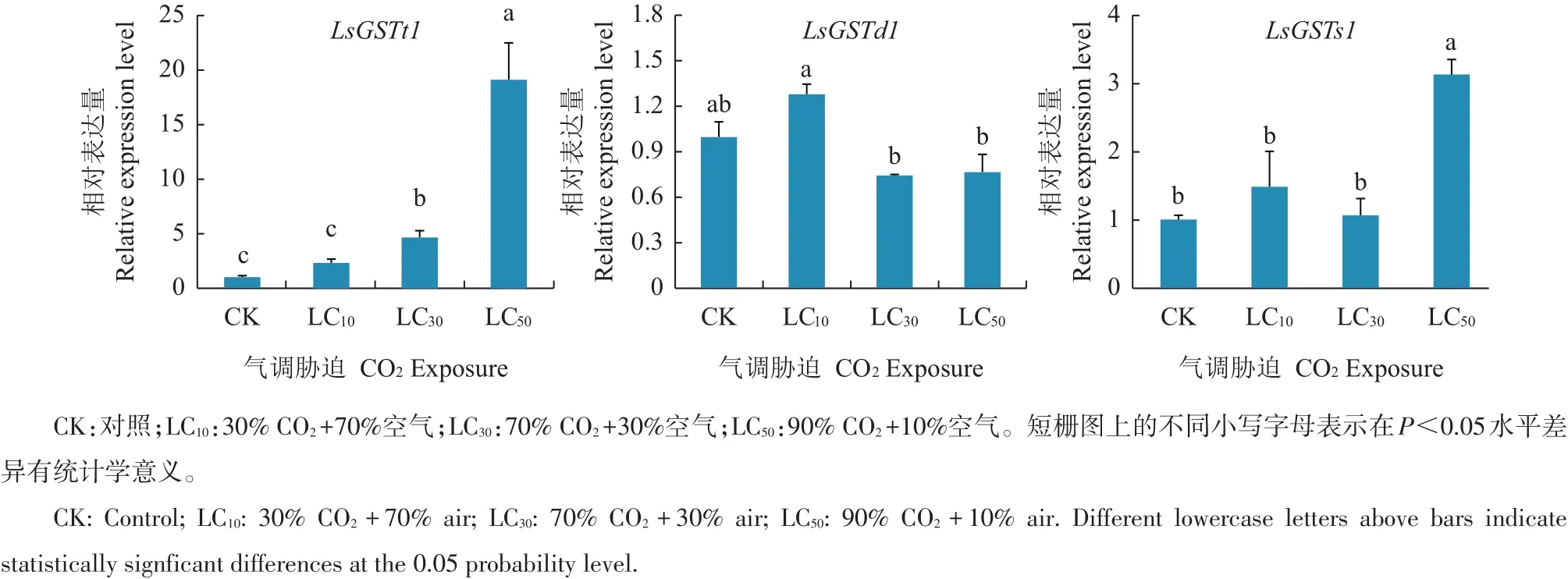
3 讨论
昆虫GSTs是一类多功能超基因家族酶系,在内源和外源化合物的解毒代谢、防御氧化、激素合成等生命活动过程中发挥着重要作用[7,9-10]。随着昆虫基因组计划开展以来,多种昆虫不同家族GSTs基因被陆续克隆,如冈比亚按蚊(Anopheles gambiae)[20]、黑腹果蝇(Drosophila melanogaster)[21]、家蚕(B.mori)[6]、赤拟谷盗(T.castaneum)[22]等。转录组测序也为昆虫GSTs基因的鉴定提供了方便,研究人员分别从斜纹夜蛾(Spodoptera litura)[23]、飞蝗(Locusta migratoria manilensis)[24]、摇蚊(Chironomus tentans)[25]和橘小实蝇(Bactrocera dorsalis)[8]的转录组数据库中鉴定获得了8、10、11和17个GSTs基因,为深入研究GSTs基因的生物学功能奠定了基础。本研究在烟草甲转录组测序基础上,通过RT-PCR技术克隆获得3个GSTs基因的cDNA全长序列,经与其他昆虫GSTs氨基酸进行序列比对和聚类分析,发现3个烟草甲GSTs分别归属于Delta、Theta和Sigma家族。其中Delta是昆虫所特有的家族,在多种杀虫剂的代谢和昆虫抗药性形成过程中起着重要作用[26],Theta和Sigma家族GSTs在昆虫中广泛分布,可能发挥着基本的解毒代谢作用[23]。
昆虫的GSTs基因在不同发育阶段的表达水平存在一定的差异,说明其在生长发育过程中发挥着不同的作用。本研究发现,3个GSTs基因在烟草甲各发育阶段均有表达,且低龄幼虫期的表达量显著高于高龄幼虫、蛹期和成虫期,这可能与它们在低龄幼虫期发挥着重要作用有关。进一步分析发现,低龄幼虫刚由卵孵化形成,需要大量取食以满足器官发育,这一时期外源化合物的摄取和代谢产生的物质随之增多,因此相关的解毒代谢酶如GSTs的mRNA表达量相应升高。橘小实蝇的5个GSTs基因(BdGSTe4、BdGSTe2、BdGSTd5、BdGSTd6和BdGSTo1)在幼虫期的表达量明显高于成虫期[8],此类现象同冈比亚按蚊[20]、小菜蛾(Plutella xylostella)[27]、斜纹夜蛾[23]等昆虫的研究结果相一致。不同物种体内的GSTs在各发育阶段的表达量变化不尽相同:如柑橘全爪螨6个GSTs基因在卵期表达量较高,推测其可能与卵发育过程中激素调节相关[28];QIN等[24]应用半定量RT-PCR分析了10个GSTs基因在飞蝗卵、一龄幼虫至成虫期的表达情况,发现LmGSTd1、LmGSTs1、LmGSTs3、LmGSTs5和LmGSTt3在各发育阶段的表达量没有明显的变化,说明这些酶属于组成型表达酶,在飞蝗整个生命过程中均起作用。
气调技术被广泛应用于仓储害虫的管理中,它主要利用高浓度CO2和低浓度O2的协同作用来提高控制害虫的效果[14,29]。CO2主要通过触杀途径起效,使昆虫神经过度兴奋,刺激昆虫气门开放,改变通风速率,导致虫体水分丧失而出现死亡[30]。不仅如此,CO2气调还可以引起昆虫体内解毒代谢酶系和靶标酶的变化,并导致其体内能源物质不断被消耗[11,15-16]。本课题组前期研究发现,CO2气调处理烟草甲成虫6 h后,GST酶活性显著高于对照,说明GSTs在烟草甲应对CO2气调胁迫过程中发挥着关键作用[17]。本研究以GSTs基因高表达的低龄幼虫为研究对象,发现经CO2气调处理6 h后,LsGSTt1和LsGSTs1表达量显著高于对照组,而LsGSTd1表达量相对于对照无显著差异,我们推测CO2气调胁迫所引起的损伤修复需要多个GSTs基因参与,通过转录和翻译合成多种GST同工酶以共同应对外源胁迫。多种GSTs基因的过量表达可能是烟草甲应对CO2气调胁迫耐受性形成的重要因素。
本研究根据烟草甲转录组信息,成功克隆了LsGSTt1、LsGSTd1和LsGSTs1基因的cDNA全长序列,这3个基因在烟草甲各发育时期均有表达,且在低龄幼虫期的表达量最为活跃。经CO2气调胁迫后,LsGSTt1和LsGSTs1表现出了明显的应激表达反应。下一步将研究烟草甲体内其他GSTs基因在气调胁迫过程中的作用,为系统阐明烟草甲GSTs基因家族对气调胁迫应答的分子机制奠定基础。
[1] SHEEHAN D,MEADE G,FOLEY V M,et al.Structure,function and evolution ofglutathione transferases:Implicationsfor classification of non-mammalian members of an ancient enzyme superfamily.Biochemical Journal,2001,360(1):1-16.
[2] LI X C,SCHULER M A,BERENBAUM M R.Molecular mechanism of metabolic resistance to synthetic and natural xenobiotics.Annual Review of Entomology,2007,52:231-253.
[3] PARKES T L,HILLIKER A J,PHILIPS J P.Genetic and biochemical analysis of glutathione-S-transferases in the oxygen defense system of Drosophila melanogaster.Genome,1993,36(6):1007-1014.
[4] AGIANIAN B,TUCKER P A,SCHOUTEN A,et al.Structure of a Drosophila sigma class glutathione S-transferase reveals a novel active site topography suited for lipid peroxidation products.Journal of Molecular Biology,2003,326(1):151-165.
[5] ROGERS M E,JANI M K,VOGT R G.An olfactory-specific glutathione-S-transferase in the sphinx moth Manduca sexta.Journal of Experimental Biology,1999,202(12):1625-1637.
[6] YU Q Y,LU C,LI B,et al.Identification,genomic organization and expression pattern of glutathione S-transferase in the silkworm,Bombyx mori.Insect Biochemistry and Molecular Biology,2008,38(12):1158-1164.
[7] QIN G H,JIA M,LIU T,et al.Characterization and functional analysis of four glutathione S-transferases from the migratory locust,Locusta migratoria.PLoS One,2013,8(3):e58410.
[8] HU F,DOU W,WANG J J,et al.Multiple glutathione S-transferase genes:Identification and expression in oriental fruit fly,Bactrocera dorsalis.Pest Management Science,2014,70(2):295-303.
[9] GULLIPALLI D,ARIF A,APAROY P,et al.Identification of a developmentally and hormonally regulated delta-class glutathione S-transferase in rice moth Corcyra cephalonica.Comparative Biochemistry and Physiology Part B:Biochemistry and Molecular Biology,2010,156(1):33-39.
[10]ZHANG Y Y,GUO X L,LIU Y L,et al.Functional and mutational analyses of an omega-class glutathione S-transferase(GSTO2)that is required for reducing oxidative damage in Apis cerana cerana.Insect Molecular Biology,2016,25(4):470-486.
[11]LI C,LI Z Z,CAO Y,et al.Partial characterization of stressinduced carboxylesterase from adults of Stegobium paniceum and Lasioderma serricorne(Coleoptera:Anobiidae)subjected to CO2-enriched atmosphere.Journal of Pest Science,2009,82(1):7-11.
[12]OMAE Y,FUCHIKAWA T,NAKAYAMA S,et al.Life history and mating behavior of a black-bodied strain of the cigarette beetle Lasioderma serricorne(Coleoptera:Anobiidae).Applied Entomology and Zoology,2012,47(2):157-163.
[13]PIMENTEL M A G,FARONI L R D,TÓTOLA M R,et al.Phosphine resistance,respiration rate and witness consequences in stored-product insects.Pest Management Science,2007,63(9):876-881.
[14]GUNASEKARAN N,RAJENDRAN S.Toxicity of carbon dioxide to drugstore beetle Stegobium paniceum and cigarette beetle Lasioderma serricorne.Journal of Stored Products Research,2005,41(3):283-294.
[15]李灿,李子忠,周波,等.高浓度二氧化碳对药材甲和烟草甲乙酰胆碱酯酶活性的影响.植物保护学报,2007,34(6):642-646.LI C,LI Z Z,ZHOU B,et al.Effect of carbon dioxide enriched atmosphere on the activity of acetylcholinesterase in adults of Stegobium paniceum and Lasioderma serricorne.Journal of Plant Protection,2007,34(6):642-646.(in Chinese with English abstract)
[16]曹宇,杨文佳,孟永禄,等.CO2气调对烟草甲的毒理作用及其能源物质的含量和利用率.西北农林科技大学学报(自然科学版),2015,43(11):123-128.CAO Y,YANG W J,MENG Y L,et al.Toxicity of CO2to Lasioderma serricorne and content and utilization of its energy substances.Journal of Northwest A&F University(Natural Science Edition),2015,43(11):123-128.(in Chinese with English abstract)
[17]李灿,李子忠.气调胁迫下3种中药材储藏期害虫谷胱甘肽转移酶活性研究.植物保护,2009,35(2):91-94.LI C,LI Z Z.GSTs activity of three pests in stored Chinese medicinal materials.Plant Protection,2009,35(2):91-94.(in Chinese with English abstract)
[18]TAMURA K,PETERSON D,PETERSON N,et al.MEGA5:Molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance,and maximum parsimony methods.Molecular Biology and Evolution,2011,28(10):2731-2739.
[19]LIVAK K J,SCHMITTGEN T D.Analysis of relative gene expression data using real-time quantitative PCR and the2-ΔΔCTmethod.Methods,2001,25(4):402-408.
[20]DING Y C,ORTELLI F,ROSSITER L C,et al.The Anopheles gambiae glutathione transferase supergene family:Annotation,phylogeny and expression profiles.BMC Genomics,2003,4:35.
[21]TU C P D,AKGÜL B.Drosophila glutathione S-transferases.Methods in Enzymology,2005,401:204-226.
[22]SHI H X,PEI L H,GU S S,et al.Glutathione S-transferase(GST)genes in the red flour beetle,Tribolium castaneum,and comparative analysis with five additional insects.Genomics,2012,100(5):327-335.
[23]HUANG Y F,XU Z B,LIN X Y,et al.Structure and expression of glutathione S-transferase genes from the midgut of the common cutworm,Spodoptera litura(Noctuidae)and their response to xenobiotic compounds and bacteria.Journal of Insect Physiology,2011,57(7):1033-1044.
[24]QIN G H,JIA M,LIU T,et al.Identification and characterisation of ten glutathione S-transferase genes from oriental migratory locust,Locusta migratoria manilensis(Meyen).Pest Management Science,2011,67(6):697-704.
[25]LI X W,ZHANG X,ZHANG J Z,et al.Identification and characterization of eleven glutathione S-transferase genes from the aquatic midge Chironomus tentans(Diptera:Chironomidae).Insect Biochemistry and Molecular Biology,2009,39(10):745-754.
[26]LOW W Y,FEIL S C,NG H L,et al.Recognition and detoxification of the insecticide DDT by Drosophila melanogaster glutathione S-transferase D1.Journal of Molecular Biology,2010,399(3):358-366.
[27]CHEN X E,ZHANG Y L.Identification and characterization of multiple glutathione S-transferase genes from the diamondback moth,Plutella xylostella.Pest Management Science,2015,71(4):592-600.
[28]LIAO C Y,ZHANG K,NIU J Z,et al.Identification and characterization of seven glutathione S-transferase genes from citrus red mite,Panonychus citri(McGregor).International Journal of Molecular Sciences,2013,14(12):24255-24270.
[29]WONG-CORRAL F J,CASTANÉ C,RIUDAVETS J.Lethal effects of CO2-modified atmospheres for the control of three Bruchidae species.Journal of Stored Products Research,2013,55:62-67.
[30]MITCHAM E J,MARTIN T A,ZHOU S.The mode of action of insecticidal controlled atmospheres.Bulletin of Entomological Research,2006,96(3):213-222.
Expression analysis of glutathione S-transferase genes in Lasioderma serricorne(Coleoptera:Anobiidae)subjected to CO2-enriched atmosphere.Journal of Zhejiang University(Agric.&Life Sci.),2017,43(5):599-607
XU Kangkang1,DING Tianbo2,YAN Yi1,LI Can1,YANG Wenjia1*
(1.College of Biology and Engineering of Environment,Guiyang University/Guizhou Provincial Key Laboratory for Rare Animal and Econimic Insects of the Mountainous Region,Guiyang 550005,China;2.College of Plant Health and Medicine,Qingdao Agricultural University,Qingdao 266109,Shandong,China)
glutathione S-transferase;Lasioderma serricorne;gene cloning;expression pattern
S 435.72;Q 965
A
10.3785/j.issn.1008-9209.2017.02.261
Summary The cigarette beetle,Lasioderma serricorne(Fabricius)(Coleoptera:Anobiidae),is a destructive stored pest distributed worldwide.Females typically oviposit in the stored materials,and the developing larvae tunnel through the stored food products on which they feed,resulting in tremendous damages to stored grains and seeds,tobacco,pet food,and dried flowers.Currently,excessive use of fumigation with insecticides has developed resistance to commonly insecticides in this insect,leading to environmental risk and its resurgence.
Controlled atmosphere(CA)treatments using low oxygen(O2)and high carbon dioxide(CO2)have been used commercially to control stored pests.The application of CA is a safe alternative way to chemical pesticides,and has been used to control the cigarette beetles.Our earlier studies showed that L.serricorne displayed a great capacity to withstandCO2stress.The activities of glutathione S-transferases(GSTs)in L.serricorne were significantly increased after exposed to CO2enriched atmosphere,implying that GSTs might be critical for tolerating to CO2stress.However,little is known about the molecular mechanism of GSTs in response to CO2exposure in L.serricorne.
国家自然科学基金(31460476);贵州省科学技术基金联合基金(黔科合LH字〔2014〕7167);贵州省高层次创新型人才培养(黔科合人才〔2016〕4020);贵阳学院高层次人才科研启动费(校人才2014003)。
杨文佳(http://orcid.org/0000-0003-4339-2158),E-mail:yangwenjia10@126.com
(First author):许抗抗(http://orcid.org/0000-0002-4203-911X),E-mail:kkxu1988@163.com
2017-02-26;接受日期(Accepted):2017-06-20
In this study,three full-length cDNAs of GSTs in L.serricorne were cloned by reverse transcription-polymerase chain reaction(RT-PCR).The deduced amino acid sequence analysis of the three genes was performed by bioinformatics methods.Quantitative real-time polymerase chain reaction(qRT-PCR)was used to establish expression profiles for those genes from different developmental stages and in response to CO2exposure.
According to the predicted GST sequences from the transcriptome datesets of L.serricorne,the full-length cDNAs of three novel genes were obtained and named LsGSTt1,LsGSTd1 and LsGSTs1(GenBank accession numbers:KY549655,KY549656,KY549657),respectively.The LsGSTt1 cDNA contained an open reading frame(ORF)of 564 nucleotides,encoding a polypeptide of 187 amino acids with an estimated molecular mass of 22.4 kDa and an isoelectric point(pI)of 8.54.The LsGSTd1 cDNA contained an ORF of 651 nucleotides,encoding a polypeptide of 216 amino acids with an estimated molecular mass of 24.5 kDa and a pI of 5.95.The LsGSTs1 cDNA contained an ORF of 696 nucleotides,encoding a polypeptide of 231 amino acids with an estimated molecular mass of 26.2 kDa and a pI of 5.06.Homology analysis indicated that the predicted amino acid sequences of the three genes had typical features of GSTs,including an N-terminal domain and a C-terminal domain.Phylogenetic analysis revealed that the three genes belonged to three different cytosolic classes,including Theta(LsGSTt1),Delta(LsGSTd1)and Sigma(LsGSTs1).Temporal expression profile revealed that all the three GSTs genes were constitutively expressed in the testing stages,exhibited similar developmental expression patterns,with the highest expression level measured in the early larval stage.Compared with the control,the mRNA levels of LsGSTd1 did not change significantly following exposure to CO2stress.However,when exposed to 50%lethal concentration(LC50)of CO2,the expression levels of LsGSTt1 and LsGSTs1 were significantly increased.
In conclusion,the results suggest that LsGSTt1 and LsGSTs1 in L.serricorne might play important roles in defense responses challenged by CO2stress.This study provides the basis for clarifying the response mechanism of insects to CO2stress.
