珍稀濒危植物十齿花属的花药发育和雄配子体发生及其系统学意义
2017-11-11张梓袁刘晓晓孔冬瑞
张梓袁 刘晓晓 李 璐* 孔冬瑞 彭 华
(1.西南林业大学云南生物多样性研究院,昆明 650224; 2.鲁东大学生命科学学院,烟台 264025; 3.中国科学院昆明植物研究所东亚植物多样性与生物地理学重点实验室,昆明 650201)
珍稀濒危植物十齿花属的花药发育和雄配子体发生及其系统学意义
张梓袁1刘晓晓1李 璐1*孔冬瑞2彭 华3
(1.西南林业大学云南生物多样性研究院,昆明 650224;2.鲁东大学生命科学学院,烟台 264025;3.中国科学院昆明植物研究所东亚植物多样性与生物地理学重点实验室,昆明 650201)
十齿花属(Dipentodon)为东亚特有单型属,是国家二级保护植物,其系统位置长期有争议。本研究连续三年的野外观察结果表明十齿花(Dipentodonsinicus)的结籽率较低(4.31%)。同时利用常规石蜡制片技术,观察了十齿花的花药发育和雄配子体发育过程。十齿花的花药四室,花药壁发育类型为基本型。成熟花药壁有6层,由表皮,药室内壁,2层中层和2层绒毡层组成,绒毡层为腺质绒毡层。花药成熟时,纤维状加厚发生于药室内壁,便于花药开裂散粉。小孢子母细胞减数分裂的孢质分裂方式为同时型,小孢子四分体多为四面体型,稀左右对称型。成熟花粉粒为二细胞型,具3孔沟。在小孢子发育的减数分裂、小孢子四分体和二细胞型花粉时期,观察到发育不正常的现象,分别占40%、48%和36%。通过与其他亲缘类群的胚胎学特征比较,支持最新分子系统学把它独立成科,放在十齿花目的观点。雄配子体发育过程中,十齿花存在着花粉败育现象,这可能是导致其结实率低的原因之一。
花药发育;十齿花属;小孢子发生;系统学;珍稀濒危物种
十齿花属(Dipentodon)仅十齿花(D.sinicus)一个种[1],为东亚特有种,分布于我国西南部以及毗邻的缅甸、印度东北部和越南北部,呈斑块状间断分布[2],生长在海拔800~2 800 m,气候凉爽湿润的山地。
自十齿花属建立以来,其系统位置一直有争议。由于它独特的形态学特征,曾被放在不同的目和科,如卫矛目的卫矛科(Celastraceae)[3];或堇菜目(Violales)的大风子科(Flacourtiaceae)(还包括了天料木科Samydaceae)[4~6],或独立为十齿花科(Dipentodontaceae)[7],把它放在蔷薇目(Rosales)内的金缕梅科(Hamamelidaceae)和蔷薇科(Rosaceae)之间[8~11]。Cronquist认为十齿花属应该放在檀香目(Santalales)[12]。值得一提的是,Dunn在1911年建立十齿花属时并没有指定模式标本,直到近100年后才被确定了后选模式标本[13]。
面对十齿花属所面临的分类学难题,比较形态学结果认为它与上述几个科都没有较近的亲缘关系,建议把它作为卫矛科中一个独立的亚科来看待[14]。早期的分子系统学证据也不支持十齿花属与这些类群有任何亲缘关系,把它视为系统位置不确定的类群[15~16]。基于叶绿体基因的分子研究认为,十齿花属与从省沽油科(Staphyleaceae)分离立出来的瘿椒树属(Tapiscia)为姐妹群,更靠近锦葵目(Malvales)和无患子目(Sapindales)[1]。来自于核基因和叶绿体基因的系统树进一步把核子木属(Perrottetia)从卫矛科中分离出来,连同十齿花属和瘿椒树属组成一个新的分支[17]。随着证据的不断完善,最近的分子系统[18~19]认为十齿花属和瘿椒树属应该独立为十齿花科,放在十齿花目(Huerteales)。
另一方面,由于生长环境受到人为活动的干扰,十齿花的居群数量不断下降,生长范围不断缩减,被列为中国珍稀濒危植物[20],《中国物种红色名录》也把它列为易危物种[21]。在植物系统进化上,十齿花属是一个古老的高级孤立类群,几乎没有姊妹分类群,保持着许多原始的性状[22],染色体的高度对称性表明了十齿花在进化上的原始性[23]。由于单型属十齿花属在植物系统进化方面具有重要的科学意义,对生物多样性和进化历史的贡献比其它多型类群更大,应该受到优先保护[24]。
目前,十齿花的研究主要有基于形态比较的分类[14],谱系地理学和保护遗传学[23,25],系统与进化[1,17]群落物种多样性[26~27],化学成分分析[28~29],组织培养与快速繁殖[30]等方面,胚胎学研究至今尚属空白。本文观察了十齿花属小孢子发生及雄配子体发育过程,为解决它的系统学问题提供胚胎学证据;同时胚胎学的研究也有助于揭示该物种濒危的繁殖生物学机制。
1 材料和方法
1.1 野外采集调查地点
十齿花的实验材料采自云南省昭通市彝良县小草坝(104°16′0″E,24°78′1″N;海拔1 773 m)凭证标本馆藏于中国科学院西双版纳热带植物园标本馆(HITBC No.154856)。栖息地的野外观察发现,十齿花仅零星分布于云南省昭通市彝良县小草坝的山地、路边等地,高大乔木状的十齿花在林中较少见,且多呈灌木状。当地居民对十齿花了解甚少,常被作为薪材砍伐,人类活动及修路等原因导致生境毁坏及森林片段化,使其遭到严重破坏,在分布区资源通常较稀少。
1.2 研究方法
(1)开花结实调查。从2014~2016年,连续三年的5月份花期、6月份幼果期和8月份成熟果期,进行野外开花和结果率的调查。每次调查数据,取结实盛期(树龄10~20年),生长健康的代表性样株10棵,标记后,每株按照不同方位随机选择10条花枝,记录其开花期、幼果期和成熟果期的花朵和果实数目;统计每个枝条上的开花数和果实数的平均值、最大值和最小值;计算幼果期的座果率和成熟果期的结实率。
(2)石蜡切片。从2014年6月至2016年6月,对十齿花进行野外观察,采集不同发育时期的花蕾和花朵以及幼果等。固定于FAA固定液(50%酒精∶冰醋酸∶甲醛=90∶5∶5)中,系列酒精浓度脱水,常规石蜡切片法制片,切片厚度约为5~8 μm,爱式苏木精染色,中性树胶封片,参考植物组织制片学[31]的方法。在Leica DM750型显微镜进行镜检和照相。花药的发育类型判断按照被子植物生殖生物学标准[32]。
2 研究结果
2.1 开花结实的野外观察
在云南省昭通市彝良县小草坝,十齿花多为小乔木或灌木状,生长在溪流畔或路边。它为落叶树种,三月份萌动枝芽及花芽,四月份在当年生枝条上现蕾,五月份盛花期,六月份开始坐果,七八月份果实相继成熟,九月份叶子变黄凋落,十月份进入休眠期。
连续3年(2014~2016)的野外观察表明,十齿花开花繁茂,花朵数目繁多,结实率低;居群树龄平均为成年树,很少见到林下小苗,表现出种质衰退之趋势。每个枝条上的开花数量的平均值为352朵;在幼果期和成熟果期,每个枝条上的座果率和结实率平均值分别为16.83%和4.31%,说明在不同的发育阶段存在着果实和种子的败育现象。三年的统计数据(表1)说明以下几点:
(1)每个枝条上的花序数平均值为12,最大值为16,最小值为8;组成一个花序的小花数量,平均值为35,最高值为41,最低值为32;每个枝条上的小花数目平均值为352,最高值为376,最低值为302。
(2)六月份幼果期,每个枝条上的果实数目,平均值为59个,最高值为72.67,最低值为0,平均座果率16.83%。
(3)八月份果实成熟期,当年生枝条上的果实数目,平均值为15,最大值为34,最小值为0,平均座果率为4.31%,每个成熟果实仅含一枚种子。
因此,成熟果期的座果率相当于结实率。连续三年的野外调查,未见到文献报道的十齿花果实成熟自动裂开,种子飞弹出去的现象[23]。
表1十齿花连续三年(2014~2016),当年生枝条上的开花、幼果期和成熟果的数目统计表(n= 30)
Table1Thenumberoftheflowers,immatureandmaturefruitsonthe1-yearoldbranchesforsuccessivethreeyears(2014-2016)(n=30)

年份Year花期的开花数目Thenumberoftheflowers幼果期的果实数Thenumberoftheimmaturefruits成熟果期的果实数Thenumberofthematurefruits平均座果率(果实数/小花数%)Themeannumberofthefruit⁃setting(ratio=thenumberoffruits/thenumberofflowers)平均值Themeannumber最大值Themaxiumnumber最小值Theminiumnumber平均值Themeannumber最大值Themaxiumnumber最小值Theminiumnumber平均值Themeannumber最大值Themaxiumnumber最小值Theminimnumber幼果期Thestageofimmaturefruits成熟果期Thestageofthematurefruits2014321343269566801629017.444.982015352367313638101742017.904.832016383418324586901231015.143.13平均值Mean35237630259.0072.6701534016.834.31
2.2 花序和小花的形态特征
十齿花具有圆头状聚伞花序,腋生于当年新枝,总花梗长达10厘米,盛开时高出枝条叶面,排成整齐的两列(图版Ⅰ:A,F),每个花序有平均有35朵小花。幼果期,当年生枝条上每个花序存有0~7个幼果(图版Ⅰ:B)。成熟果期,一个果序上仅有0~1个尚未开裂的成熟果,其余皆为凋落后仅存的果梗(图版Ⅰ:C,D)。小花为两性花,花萼和花瓣分别为6~7枚(图版Ⅰ:E),黄绿色或白色;雄蕊6~7枚(稀10枚),着生于花盘边缘,与基部肉质裂片互生,花药黄色,内向纵列(图版Ⅰ:G);子房不完全三室,每室有胚珠两枚。蒴果基部宿存齿状花萼,顶部宿存花柱发育为果喙(图版Ⅰ:H);每个子房仅有一枚胚珠发育为种子,光滑饱满,呈椭圆形(图版Ⅰ:I)。
2.3 花药壁的发育
十齿花的花药为四室花药(图版Ⅱ:A)。当花药原基发育到横切面呈四边形时,角隅处的表皮细胞下分化出多个孢原细胞。孢原细胞经1次平周分裂和多次垂周分裂,形成初生周缘细胞和初生造孢细胞。初生周缘细胞平周分裂产生内外两层次生周缘细胞(图版Ⅱ:B)。其中,外层继续进行1次平周分裂和多次垂周分裂,形成药室内壁和中层(图版Ⅱ:C),内层也进行1次平周分裂和多次垂周分裂,形成中层和绒毡层(图版Ⅱ:D)。由于两层中层分别来源于内层和外层次生周缘细胞,因此,十齿花的花药壁的发育类型为基本型[33]。在小孢子母细胞时期,花药壁发育完整,从外到内分别是表皮、药室内壁、2层中层和绒毡层(图版Ⅱ:E),绒毡层细胞初为单核(图版Ⅱ:E),后为2核(图版Ⅱ:G)。
随着小孢子母细胞的发育,花药壁的各层细胞发生了不同的变化。在减数分裂时,中层细胞变得细长,即将退化;绒毡层细胞径向伸长,体积增大,原生质增多,液泡化明显(图版Ⅱ:F)。在成熟花粉粒阶段,中层细胞已退化,表皮细胞逐渐拉长变扁平;药室内壁细胞增大;绒毡层细胞原地向花粉囊内释放营养物质,逐渐被吸收,细胞轮廓模糊(图版Ⅱ:G,H),因此绒毡层为腺质型[33]。花粉粒成熟时,表皮细胞退化至波浪状,药室内壁细胞壁纤维性加厚,绒毡层完全被吸收(图版Ⅱ:I)。

图版Ⅰ 十齿花(D.sinicus)的形态图 A.盛花期,当年生枝条上的花序呈两列排列,花序数平均为15;B,C.幼果期,每个枝条上仅有部分小花发育为幼果,平均座果率为16.83%;D.果实发育后期的果枝,平均座果率降低为4.31%;E.早期的头状聚伞花序,外面有一轮苞片;F.发育后期的头状聚伞花序;G.示花被、雄蕊数目、花盘肉质裂片等特征,侧面观(左),极面观(右);H.果实发育早期,花萼和花柱宿存;I.成熟果期,宿存萼片逐渐变小成齿状,宿存柱头形成果喙,一个果实里仅含有一枚种子Plate Ⅰ The morphology of D.sinicus A. Flowering stage,there are about 15 inflorescences arranging in two lines on a branch; B,C. Early stage of developing fruit,the average ratio of fruit-setting is low with an average number of 16.83%; D. Late stage of developing fruit,the average ration of fruit-setting is descending to 4.31%; E. Early stage of cymose inflorescence with bracts; F. Late stage of cymose inflorescence; G. Indicating the number and the arrangement of perianth and stamens in a flower,side view(left),polar view(right); H. Early stage of developing fruit,both perianth and style are persistent; I. Late Stage of developing fruit,indicating the persistant perianth becoming small and stigma forming a pod beak,each capsule contains only one seed

图版Ⅱ 十齿花(D.sinicus)的花药壁发育 A.花药具4个药室;B.初生壁细胞平周分裂,形成外层和内层。箭头所指为一个正在平周分裂的初生壁细胞;C.外层细胞平周分裂(箭头);D.内层细胞平周分裂(箭头);E. 5层花药壁,由外到内分别为表皮、药室内壁、2层中层和绒毡层;F.中层细胞变得扁平细长,绒毡层细胞发达;G,H. 2核绒毡层细胞逐渐解体,药室内壁细胞变宽;I.散粉时的花药壁,表皮解体和药室内壁纤维性加厚 (en.药室内壁;ep.表皮;il.内生周缘细胞层;ml.中层;ol.外生周缘细胞层; ta.绒毡层)Plate Ⅱ Anther wall development of D.sinicus A.The anther has four sporangia; B.The primary parietal layer undergoes periclinal division(arrow points to),resulting in the outer and inner parietal layers; C.The cell of the outer secondary parietal layer is undergoing periclinal division (arrow points to); D.The cell of the inner secondary parietal layer is undergoing periclinal division (arrow points to); E.Five-layered anther wall comprising epidermis,endothecium,two middle layers and tapetum; F.The middle is thin and flat and the tapetum is well-developed; G,H. The tapetum with 2 nuclei in each cell disintegrated slowly and the endothecium is widen; I.Crushed epidermis and fibrous endothecium at the stage of disperse (en.Endothecium; ep.Epidermis; il.Inner secondary parietal layer; ml.Middle layer; ol.Outer secondary parietal layer; ta.Tapetum)
2.4 小孢子发生及雄配子体发育
早期初生造孢细胞有丝分裂形成次生造孢细胞,进而发育成小孢子母细胞(图版Ⅲ:A),其体积大,核仁大而显著,多边形,细胞质浓厚。小孢子母细胞经过减数分裂(图版Ⅲ:B-G)发育成小孢子四分体。由于在小孢子母细胞减数分裂前期Ⅱ,两核间不伴随细胞板的形成(图版Ⅲ:D,E),因此,小孢子母细胞孢质分裂为同时型。四分体由胼胝质壁包被和分隔,多四面体型,稀左右对称型(图版Ⅲ:G)。随后胼胝质解体,小孢子从四分体中释放出,刚释放的小孢子呈球状,细胞质较浓,细胞核位于中央。散粉时,成熟花粉粒为二细胞型,三萌发孔(图版Ⅲ:H)。
2.5 花药发育过程中的异常现象
十齿花在小孢子发生及雄配子体发育过程中,观察到败育现象,在所统计的500张切片中,约40%的小孢子在减数分裂过程出现细胞轮廓不清楚,细胞核形态各异(图版Ⅲ:I)。小孢子四分体时期时,所统计的500个片子中,约48%的细胞形态不正常,二核形态不均一,呈不规则形(图版Ⅲ:J)。单核花粉与二细胞型花粉时期,所统计的500张花药横切片中,每个花药横切面上,约有36%的细胞表现出败育特征,花粉细胞轮廓呈不规则圆形(图版Ⅲ:K,L)。
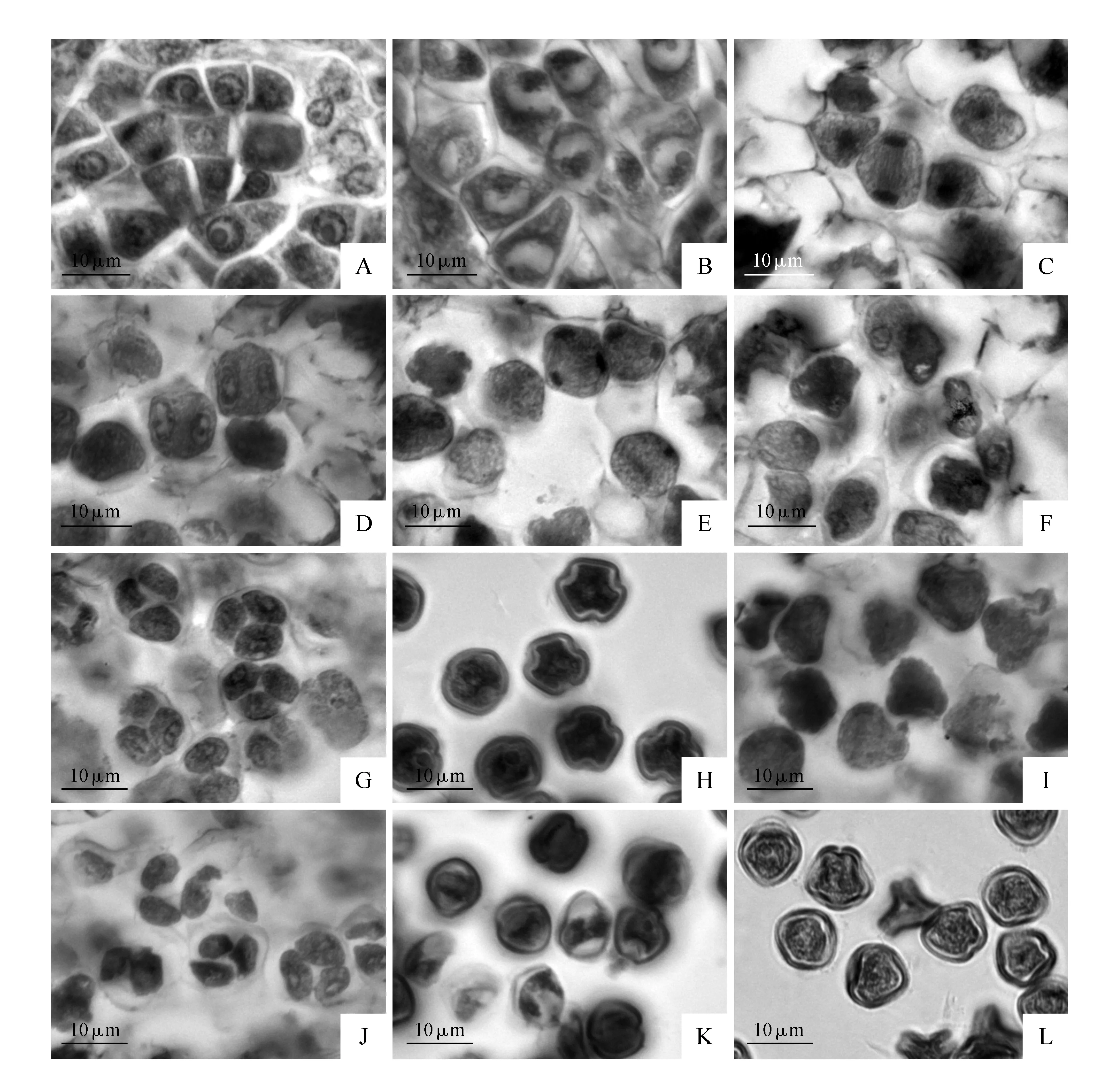
图版Ⅲ 十齿花(D.sinicus)的雄配子体发育 A.次生造孢细胞,箭头示次生造孢细胞有丝分裂;B-F.小孢子母细胞减数分裂:B.减数分裂Ⅰ前期;C.减数分裂Ⅰ后期;D.减数分裂Ⅱ前期;E.减数分裂Ⅱ后期;F.减数分裂Ⅱ末期;G.小孢子四分体;H.成熟花粉粒为二细胞型,具3孔沟;I.小孢子母细胞减数分裂过程中的异常现象;J.部分小孢子四分体发育不正常;K,L.从四分体中释放出来的小孢子,部分细胞形态不规则Plate Ⅲ Microsporogenesis of D.sinicus A. The structure of secondary sporogenous cells,the arrow refers to the secondary sporogenous cell mitosis; B-F. Meiosis of microspore mother cell; B. Prophase Ⅰ; C. Anaphase Ⅰ; D. Prophase Ⅱ; E. Anaphase Ⅱ; F. Telophase Ⅱ; G. Microspore tetrads; H. The mature pollen grains are two-celled with three colpates; I. Abnormality of meiotic anaphase Ⅱ stage; J. Abnormal microspore tetrads; K,L. Abnormal microspores released from tetrads
3 讨论
在被子植物胚胎学综述类书籍中,十齿花的胚胎学研究一直是空白[34]。本文详细报道了十齿花的小孢子发生和雄配子体的发育过程,它具有以下胚胎学特征。花药四室,花药壁发育类型为基本型。花药壁由表皮、药室内壁、中层(2层)和绒毡层组成,绒毡层为腺质绒毡层。花药成熟时,纤维状加厚发生于药室内壁,便于花药开裂散粉。小孢子母细胞减数分裂的孢质分裂方式为同时型,小孢子四分体多为四面体型,稀左右对称型,成熟花粉粒为二细胞型,具3孔沟。在小孢子发育的减数分裂、小孢子四分体和二细胞型花粉粒时期,观察到花粉发育不正常的现象,分别占40%、48%和36%。
比较胚胎学在解决一些系统位置不确定的关键类群方面,一直发挥着重要的作用[32~37]。本文把十齿花属和传统分类系统中相关类群中已有的胚胎学资料[34]做了比较分析。结果表明,在花药壁发育和雄配子体发生方面,十齿花属和卫矛科、大风子科、金缕梅科和铁青树科等具有一些共同特征,如花药壁发育为基本型,腺质性绒粘层,花药壁纤维加厚发生在药室内壁;小孢子减数分裂具同时型胞质分裂,四分体多为四面体型等。然而,部分花药发育特征也存在明显差异,不支持它和这些科具有较近的亲缘关系。例如,本研究结果中观察到十齿花的花药为四室,并不支持它放置于檀香目内,因为后者以花药二室为典型特征,如铁青树科和檀香科[34]。金缕梅科的绒毡层细胞内壁上有球状体,而十齿花属并无此特征。从现有的胚胎学证据来看,结合形态分析的推论[14],十齿花属与卫矛科的胚胎学特征较为相似,倾向于支持两者之间有较近的亲缘关系。
然而,十齿花属与卫矛科的亲缘关系得不到分子证据的支持。近年来的分子系统学证据并不认为十齿花属与上述传统类群间有任何关系,而是把它置于被子植物分子系统位置不确定的科[15~16]。无独有偶,当Merrill在1941年把十齿花属提升为十齿花科时[7],也认为该科在双子叶植物系统中难以找到合适的位置。基于叶绿体基因的分子系统学研究[1]认为,十齿花属和瘿椒树属为姐妹群,应该放在真蔷薇目Ⅱ(euroside Ⅱ)的锦葵目支(Mavids),接近锦葵目(Malvales)和无患子目(Sapindales)。来自于核基因和叶绿体基因的系统树[17]进一步把卫矛科的核子木属(Perrottetia)分离出来,连同十齿花属和瘿椒树属组成一个新的分支。最新的被子植物系统学研究[18~19]认为十齿花属和核子木属组成十齿花科,放在十齿花目Huerteales内,包括4个科5个属,除了十齿花科有2个属外,其余3个皆为单型属[38]。令人深思的是,组成十齿花目的5个属在分类史上都被视为系统位置不确定的类群。例如,核子木属原来放置于卫矛科,有15个种,分布较广,亚洲、澳洲和美洲都有。瘿椒树属曾被放在省沽油科(Staphyleaeceae),为我国特有属,现提升为瘿椒树科。非洲单型科(Gerrardinacea)内的属Gerrardina曾被放在大风子科;中美洲的Petenaeaceae科中的属Petennae原归为杜英科Elaeocarpaceae。
因此,关于十齿花属的比较胚胎学的研究范畴又转到分子系统所支持的十齿花目中的四个科五个属内。由于这五个属的胚胎学资料仅有瘿椒树属有报道,其余三个属尚需补充资料,因此本文仅把十齿花属与瘿椒树属的胚胎学[39]进行比较。结果表明,在花药发育和雄配子体发方面,两个属拥有高度一致的特征:花药四室,花药壁有五层,花药壁发育为基本型,绒毡层为腺质性绒毡层;小孢子母细胞减数分裂具同时型胞质分裂,小孢子四分体为正四面体,二细胞型花粉粒。因此,从现有的胚胎学资料来看,本文研究结果支持分子系统学的观点,即十齿花属和瘿椒树属为姐妹群[1]。当然,倘若有其它三个属的胚胎学资料,尤其是得到十齿花科的另一个属(核子木属)的胚胎学证据,那么关于十齿花科的范畴,五个属的系统位置问题,将会得到进一步的澄清。
在小孢子发育过程中,十齿花存在花粉败育现象。这说明,在小草坝居群中,十齿花的结籽粒低和花粉败育有一定的关联。为了进一步揭示十齿花的结籽粒低是否还存在着其它繁殖机制问题,本项目将继续完成它的雌配子体和胚胎发育研究。这将为十齿花面临着的座果率低、结籽粒低、幼苗更新少、种群濒危等科学问题提供较为系统的繁殖生物学依据。
致谢西南林业大学云南生物多样性研究院和国家林业局西南地区生物多样性保育重点实验室提供实验平台;实验结果的图像采集借助西南林业大学大型仪器共享平台生物正置荧光显微镜Leica DM2500完成;云南省林业勘探设计院的董洪进博士采集部分实验材料;中科院昆明植物研究所的向春雷博士对本文提出宝贵意见。
1.Peng Y L,Chen Z D,Gong X,et al.Phylogenetic position ofDipentodonsinicus:evidence from DNA sequences of chloroplastrabL,nuclear ribosomal 18S,and mitochondriamatR genes[J].Botanical Bulletin Academia Sinica,2003,44:217-222.
2.傅立国.中国植物红皮书:稀有濒危植物:第1册[M].北京.科学出版社,1992:208-209.
Fu L K.China Plant Red Data Book.Rare and Endangered Plants:Vol.1[M].New York:Science Press,1992:208-209.
3.Dunn A.Dipentodon.A new genus of uncertain systematic position[J].Bulletin of Miscellaneous Information(Royal Botanic Gardens,Kew),1911,7:310-313.
4.Sprague T A.Samydaceae[J].Journal of Botany,British and Foreign,1925,63:9-13.
5.Fischer C E C.Contributions to the flora of Burma:XVⅢ[J].Bulletin of Miscellaneous Information(Royal Botanic Gardens,Kew),1940,7:288-294.
6.Metcalfe C R,Chalk L.Anatomy of the dicotyledons:leaves,stem and wood in relation to taxonomy with notes one economic uses[M].Oxford:Clarendon Press,1950,1:1120-1127.
7.Merrill E D.Dipentodontaceae.Plants collected by Captain F Kindon-Ward on the Vernay-Cutting expedition[J].Brittonia,1941,4:69-73.
8.Hutchinson J.The Families of flowering plants:Vol.1[M].Dicotyledons.2nd ed.Oxford,1959:329.
9.Hutchinson J.The families of flowering plants arranged according to a new system based on their probable phylogeny:3rd ed[M].London,1973:415.
10.Dahlgren R M T.A revised system of classification of the angiosperms[J].Botanical Journal of the Linnean Society,1980,80:91-124.
11.Nee M,Takhtajan A.Diversity and classification of flowering plants[J].Brittonia,1998,50(2):191.
12.Cronquist A.An integrated system of classification of flowering plants[M].New York:Columbia University Press,1981:681.
13.Peng H.Lectotypification of the NameDipentodonsinicus(Dipentodontaceae)[J].Taxon,2001,50(4):1177-1178.
14.刘建生,诚静容.十齿花属分类地位的研究[J].武汉植物学研究,1991,9(1):29-39.
Liu J S,Cheng J R.On the systematic position of genusDipentodonDunn[J].Journal of Wuhan Botanical Research,1991,9(1):29-39.
15.Angiosperm Phylogeny Group[APG Ⅰ].An ordinal classification for the families of flowering plants[J].Annals of the Missouri Botanical Garden,1998,85:531-553.
16.Angiosperm Phylogeny Group[APG Ⅱ].An update of the angiosperm phylogeny group classification for the Orders and Families of Flowering Plants[J].Botanical Journal of the Linnean Society,2003,141:399-436.
17.Zhang L B,Simmons M P.Phylogeny and delimitation of the Celastrales inferred from nuclear and plastid Genes[J].Systematic Botany,2006,31(1):122-137.
18.Angiosperm Phylogeny Group[APG Ⅲ].An update of the angiosperm phylogeny group classification for the orders and families of flowering plants[J].Botanical Journal of the Linnean Society,2009,161:105-121.
19.Angiosperm Phylogeny Group[APG Ⅳ].An update of the angiosperm phylogeny group classification for the orders and families of flowering plants[J].Botanical Journal of the Linnean Society,2016,181:1-20.
20.傅德志,傅立国,左家哺,等.中国被子植物物种多样性[M].//宋延龄,杨亲二,黄永青.物种多样性研究与保护.杭州:浙江科学技术出版社,1998,3:48-78.
Fu D Z,Fu L G,Zuo J P,et al.Status and conservation of angiosperm diversity in China[M],//Song Y L,Yang Q E,Huang Y Q.Research and conservation of species diversity.Hangzhou:Zhejiang Science and Technology Publishing House,1998,3:48-78.
21.汪松,解焱.中国物种红色名录.第1卷:红色名录[M].北京:高等教育出版社,2004:351.
Wang S,Xie Y.China species red list Vol.1:Red List[M].Beijing:Higher Education Press,2004:351.
22.Thome R F.Eastern Asia as a living museum for archaic angiosperms and other seed plants[J].Taiwania,1999,44:413-422.
23.袁庆军.十齿花谱系地理学和保护遗传学[D].昆明:中国科学院植物研究所,2007.
Yuan Q J.A Phylogeographic and conservation genetic study ofDipentodonsinicus[D].Kunming:Institute of Botany,Chinese Academy of Sciences,2007.
24.Krajewski C.Phylogeny and diversity[J].Science,1991,254:918-919.
25.Yuan Q J,Zhang Z Y,Peng H,et al.Chloroplast phylogeography ofDipentodon(Dipentodontaceae) in Southwest China and Northern Vietnam[J].Molecular Ecology,2008,17:1054-1065.
26.林长松,左经会,廖雯稀.稀有植物十齿花群落物种多样性研究[J].植物研究,2008,28(3):354-358.
Lin C S,Zuo J H,Liao W X.Species diversity of rare plantDipentodonsinicuscommunities in Yushe National Forest Park[J].Bulletin of Botanical Research,2008,28(3):354-358.
27.苏文苹,杜凡,杨宇明,等.昭通北部地区稀有植物十齿花群落特征[J].福建林业科技,2015,42(3):54-59.
Su W P,Du F,Yang Y M,et al.Study on community characteristics of rareDipentodonsinicusin Northern Zhaotong[J].Journal of Fujian Forestry Science and Technology,2015,42(3),54-59.
28.叶冠,彭华,范明松,等.十齿花化学成分研究[J].中草药, 2008,39(6):808-810.
Ye G,Peng H,Fan M S,et al.Chemical constituents ofDipentodonsinicus[J].Chinese Traditional and Herbal Drugs,2008,39(6):808-810.
29.秦向东,刘吉开.十齿花化学成分研究[J].天然产物研究与开发,2014,26(5):671-674.
Qin X D,Liu J K.Chemical constituents ofDipentodonsinicus[J].Natural Product Research and Development,2014,26(5):671-674.
30.薛媛,朱忠荣,王涛.十萼花组织培养与快速繁殖[J].植物生理学通讯,2005,41(3):349.
Xue Y,Zhu Z R,Wang T.Tissue culture and rapid propagation ofDipentodonsinicus[J].Plant Physiology Communications,2005,41(3):349.
31.李正理.植物组织制片学[M].北京:北京大学出版社,1996:72-73.
Li Z L.Technology of plant tissue section[M].Beijing:Beijing University Publishing Company,1996.
32.胡适宜.被子植物生殖生物学[M].北京:高等教育出版社,2005.
Hu S Y.Reproductive biology of angiosperms[M].Beijing:Higher Education Press,2005.
33.Davis G L.Systematic embryology of the angiosperms[M].New York:John Wiley & Sons Inc,1966.
34.Johri B M,Ambegaokar K B,Srivastava P S.Comparative embryology of angiosperms:2 Vols[M].New York:Springer-Verlag,1992.
35.Maheshwari P.An introduction to the embryology of angiosperms[M].London:McGraw-Hill.Mathew,1950.
36.Tobe H.The embryology of angiosperms:its broader application to the systematic and evolutionary study[J].Botanical Magazine Tokyo,1989,102:351-367.
37.Li L,Liang H X,Peng H,et al.Sporogenesis and gametogenesis inSladeniaand their systematic implication[J].Botanical Journal of the Linnean Society,2003,143:305-314.
38.Christenhusz M J M ,Fay M F,Clarkson J J,et al.Petenaeaceae,a new angiosperm family in Huerteales with a distant relationship toGerrardina(Gerrardinaceae)[J].Botanical Journal of the Linnean Society,2010,164:16-25.
39.康华钦,刘文哲.瘿椒树大小孢子发生及雌雄配子体发育解剖学研究[J].西北植物学报,2008,28(5):0868-0875.
Kang H Q,Liu W Z.Anatomical studies on megasporogenesis,microsporogenesis and development of female and male gametophyte inTapisciasinensisOliver[J].Acta Botanica Boreali-Occidentalia Sinica,2008,28(5):0868-0875.
The National Natural Science Foundation of China(NSFC31260045;NSFC31460040);The Scientific Research Foundation in Southwest Forestry University(2015);College Students Innovative Practice Training Program(2015) in Kunming institute of Botany,Chinese Academy of Sciences
introduction:ZHANG Zi-Yuan(1994—),female,undergraduate,major at plant conservation.
date:2016-12-19
AntherDevelopmentandMicrosporogenesisinDipentodonwithSystematicImplications
ZHANG Zi-Yuan1LIU Xiao-Xiao1LI Lu1*KONG Dong-Rui2PENG Hua3
(1.Yunnan Academy of Biodiversity,Southwest Forestry University,Kunming 650024;2.School of Life Sciences,Ludong University,Yantai 264025;3.Key Laboratory of Biodiversity and Biogeography of East Asian,Kunming Institute of Botany,Chinese Academy of Sciences,Kunming 650201)
The monotypic genusDipentodonDunn is endemic to Eastern Asia and has been rare and endangered due to the habitat loss. Its systematic position has been controversial. The investigation in a natural population showed that the average ratio of seed-setting inD.sinicuswas very low with a number of 4.31% for three successive years. The anther development and microsporogenesis ofD.sinicuswere observed by means of conventional paraffin sections. The anther is tetrasporangiate, and its anther wall comprises four or five layers: an epidermis, an endothecium, one or two middle layers and a tapetum. The development of anther wall belongs to basic type. The tapetum is glandular type, and wall thickening occurs in endothecium. Cytokinesis of microsporocyte meiotic division is simultaneous, and the tetrads are tetrachedral, occasionally isobilateral. The mature pollen grains are two-celled with three colpates. The high percentage of pollen abortion was observed in process of male gametogenesis. Comparative embryological features supported the molecular view thatDipentodonshould be included into Dipentodentaceae among order Huerteales. The abnormality in the pollen development inDipentodonmight be one of the main causes resulting to its extremely low seed-setting.
anther development;Dipentodon;microsporogenesis;systematics;rare and endangered species
国家自然科学基金(NSFC31260045;NSFC31460040);西南林业大学校级启动基金(111439);2015年度“中国科学院昆明植物研究所大学生创新实践训练计划”项目
张梓袁(1994—),女,本科生,主要从事植物保护方面的研究。
* 通信作者:E-mail:lilusaraca@foxmail.com
2016-12-19
* Corresponding author:E-mail:lilusaraca@foxmail.com
Q944.58
A
10.7525/j.issn.1673-5102.2017.04.003
