几个新疆主栽果树树种抗寒性初步研究
2017-11-09赵蕾杜研刘珩安鹭卢明艳张东亚
赵蕾,杜研,刘珩,安鹭,卢明艳,张东亚
(新疆林业科学院,新疆 乌鲁木齐 830063)
几个新疆主栽果树树种抗寒性初步研究
赵蕾,杜研,刘珩,安鹭,卢明艳,张东亚
(新疆林业科学院,新疆 乌鲁木齐 830063)
以新疆9个常见果树的1年生休眠枝条为试材,在低温胁迫下测定其电解质渗出率,用Logistic曲线拟合求出拐点温度即半致死温度作为各试材抗寒力评价的参考指标。结果表明:9个树种中西梅和砀山梨的抗寒性最强,黄连木最弱。抗寒性由强至弱为西梅>砀山梨>库尔勒香梨>枣>扎343 >新新2>温185>阿月浑子>黄连木。
果树;抗寒性;半致死温度;恢复生长法
新疆光照资源丰富,昼夜温差大,降水稀少而采用人工灌溉,是极为适合落叶果树的生态区,历来就有瓜果之乡的美誉。然而,新疆冬季严寒,部分地区最低温可达-40 ℃,寻找抗寒性强的树种在寒冷地区推广,具有深远的现实意义。
电解质渗出率法在鉴定植物抗寒性及杂种实生苗抗寒性早期预测等方面都有广泛的应用。在柑橘[1]、苹果[2]、葡萄[3]、李[4]等果树,多种粮食作物及热带作物枇杷[5]等方面都有许多成功运用的实例。本文采用电解质渗出率法拟合Logistic方程,对新疆常见的几个果树树种的抗寒性进行了初步研究,以期为我区果树抗寒栽培和抗寒育种提供理论参考。
1 材料与方法
1.1 试验材料
供试材料取自轮台国家果树资源圃,试材为阿月浑子、黄连木、西梅、枣、库尔勒香梨、砀山梨以及核桃的三个品种(温185、扎343和新新2)。
1.2 处理方法
1.2.1 低温处理 2015年11月从树冠外围中层选取粗度均匀一致的一年生休眠枝条,在-10 ℃冰箱中冷藏备用。2016年1月将冷藏保存的枝条依次用自来水、蒸馏水、重蒸馏水冲洗,擦干后,将各样树的供试枝条分成10组。留下其中1组,测定其电导率作为对照(-10 ℃)。其余9组用塑料袋包好,放入低温冰箱中分别进行-12、-15、-18、-21、-24、-27、-30、-33、-36 ℃的低温处理,降温速度为3 ℃·h-1,达到所要求的温度后,维持10 h。处理结束后以同样的速度升至室温。
1.2.2 恢复生长 冷冻处理后,每处理每供试单株留取等长枝段5~10枝。以蛭石为基质在光照培养箱中培养,湿度70%,日间25 ℃,夜间17 ℃。1个月后计算萌芽率。
1.3 研究方法
低温处理过的枝条在室温下放置1 h后,剪取0.3~0.5 cm的小段,称重1.0 g,放入大试管中,加20 mL双重蒸馏水浸泡10 h,用EC-215型电导仪测出初电导值(S1),然后将大试管用塑料薄膜封口,放入水浴锅中煮沸20 min,自然冷却,摇匀,并在室温下平衡10 min,测定终电导率值(S2)。对照为未进行冷冻处理的枝条。以上测定每处理重复6次。按以下公式计算出电解质渗出率:电解质渗出率(Y)=(S1/S2)×100
其中,Y—电导质渗出率,S1—初电导率,S2—终电导率。
1.4 数据分析
根据初电导率(S1)和终电导率(S2),求出电解质渗出率(y),并对其进行回归分析,将温度(t)和电解质渗出率(y)间的关系用Logistic方程y=k/(1+ae-bt)进行拟合,若拟合度(r)显著时,再计算半致死温度(LT50)。
2 结果与分析
2.1 不同低温处理后电解质渗出率的变化
9个供试果树休眠枝条经不同低温处理后,从电解质渗出率的变化趋势来看,随处理温度的降低,各参试材料的电解质渗出率逐渐升高,规律较明显。变化趋势基本上符合温度越低对细胞膜造成的损伤越大及细胞膜的透性增大将导致细胞内电解质渗出率也增大这一电导法测定抗寒力的基本理论。各试材低温处理后电解质渗出率见图1,从图1中所示电解质渗出率的曲线看出,在一系列冷冻低温处理后,9个树种休眠枝条的组织电解质渗出率与处理温度之间呈急剧或缓慢上升的“S”形曲线,虽然有个别数据在拐点之后出现电解质渗出率小幅下降的现象,但不影响总体的变化规律。不同的树种休眠枝条电解质渗出率急剧上升时的处理低温各不相同,如温185在处理低温-21 ℃之后枝条电解质渗出率开始急速上升,新新2、阿月浑子、黄连木的拐点都出现在-18 ℃左右,到-24 ℃往后逐渐平缓;扎343的拐点出现在-21 ℃,枣的拐点出现在-24 ℃,这两个树种拐点的结束温度都出现在-27 ℃;香梨、砀山梨和西梅电解质渗出率急速升高的区域出现在-27 ℃附近。拐点以前,电解质渗出率不断增加,表明降温对细胞膜伤害率的递增效应不断增大,到拐点时最大;之后,逐渐下降,表明降温对细胞膜伤害率的递增效应减小。拐点温度上表现出的差异可以用于衡量其耐受低温的能力,且这种差异表现为拐点温度与抗寒力之间呈负相关趋势,拐点温度越低,抗寒性越强[6]。
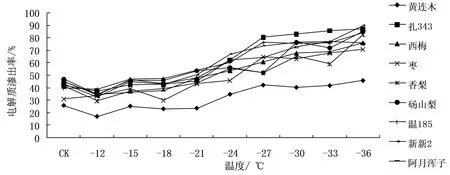
图1 低温处理后电解质渗出率的变化曲线
2.2 配合Logistic方程求低温半致死温度
供试果树休眠枝条在不同处理温度下电解质渗出率曲线基本符合Logistic变化规律,从表1的Logistic方程拟合统计结果可以看出,在低温胁迫下,相对电解质渗出率与温度之间的关系曲线,能较好地用Logistic曲线方程y=k/(1+ae-bx)进行拟合,相关系数介于0.883 2~0.960 8之间,进行F值(F0.01)检验表明,供试9个果树枝条的拟合值均大于相关系数临界值r0.01=0.841,呈极显著水平,说明在该抗寒研究中,不同低温处理电解质渗出率遵循Logistic方程的变化规律且与半致死温度呈线性关系,其拟合结果可靠,精确度较高。
表1 电解质渗出率回归方程及半致死温度(LT50)
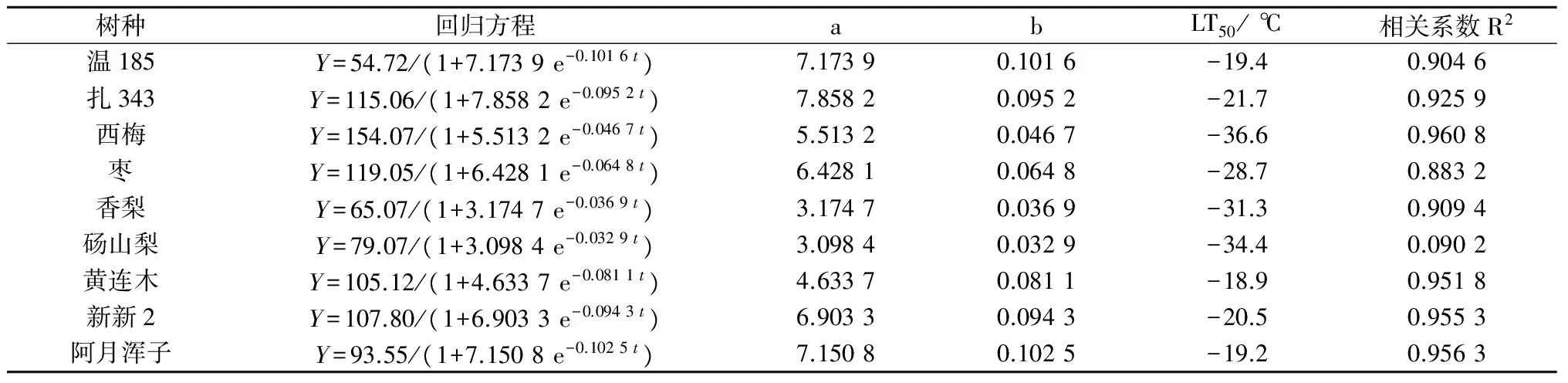
树种回归方程abLT50/℃相关系数R2温185Y=54.72/(1+7.1739e-0.1016t)7.17390.1016-19.40.9046扎343Y=115.06/(1+7.8582e-0.0952t)7.85820.0952-21.70.9259西梅Y=154.07/(1+5.5132e-0.0467t)5.51320.0467-36.60.9608枣Y=119.05/(1+6.4281e-0.0648t)6.42810.0648-28.70.8832香梨Y=65.07/(1+3.1747e-0.0369t)3.17470.0369-31.30.9094砀山梨Y=79.07/(1+3.0984e-0.0329t)3.09840.0329-34.40.0902黄连木Y=105.12/(1+4.6337e-0.0811t)4.63370.0811-18.90.9518新新2Y=107.80/(1+6.9033e-0.0943t)6.90330.0943-20.50.9553阿月浑子Y=93.55/(1+7.1508e-0.1025t)7.15080.1025-19.20.9563
2.3 恢复生长对半致死温度的验证
由表2可以看出,当处理温度处于-21 ℃以上时,各供试树种都有较高的萌芽率;经过-24 ℃的低温处理后,温185、黄连木和新新2这3个树种没有萌芽;当温度降至-30 ℃,仅西梅、香梨和砀山梨还有萌芽率,其余树种枝条均已干枯;达到-33 ℃时,除西梅和砀山梨的枝条还有萌芽外,其余8个树种均不能正常萌芽。从枝条恢复生长试验来看,7个树种中西梅和砀山梨的抗寒力较强,而阿月浑子和黄连木的抗寒能力较弱,与半致死温度的计算结果基本一致。
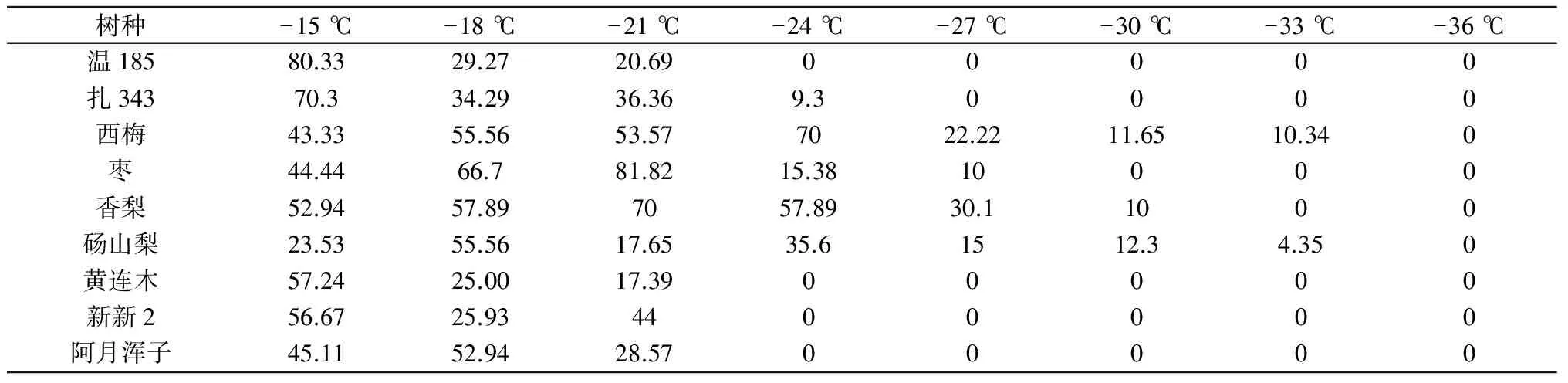
表2 不同温度处理下的萌芽率 %
3 讨论
3.1 供试9个树种的电解质渗出率曲线符合缓慢-迅速-缓慢的“S”形曲线,与大多数学者的研究结果一致[7-9]。
3.2 曹青爽[10]认为阿月浑子可耐-24~-25.2 ℃低温,与本实验结果基本吻合,在实际生产中阿月浑子适生区范围较窄,多受低温影响限制。黄连木因其较强的抗涝能力,常常作为阿月浑子的砧木被广泛应用,但从实验结果中可以看到,黄连木也存在不耐低温的问题。
3.3 虽然拟合Logistic方程计算半致死温度代表果树抗寒力的研究方法目前应用较为广泛,但植物的生理过程是错综复杂的,植物的抗性受多种因素影响,刘祖祺等[11]认为孤立地用某一指标很难反映植物的抗寒实质,也不利于揭示植物抗寒的本质。而张军科[12]等采用主成分分析法分析后发现半致死温度可以代表植物生理指标的绝大部分信息,从而验证了半致死温度可以作为快速鉴定的单因素指标。
4 结论
电解质渗出率及半致死温度的测定是影响和评价植物抗寒性的重要指标,本试验对9个新疆主栽果树1年生休眠枝条在不同低温下电解质渗出率和半致死温度进行了测定分析,9个果树品种抗寒性强弱为西梅>砀山梨>库尔勒香梨>枣>扎343 >新新2>温185>阿月浑子>黄连木,恢复生长法得出的萌芽率与半致死温度排序具有很好的一致性。尽管人工低温处理和室外自然低温有一定的差异,但9个供试果树休眠枝条是在相同的条件下进行测定,所以它们具有可比性,能够说明不同树种间抗寒性强弱。
[1] 伊华林,鲁忠芳,张志钰,等.鄂柑1号抗寒力测定与Logistic方程的应用[J].湖北农业科学,1996(3):46-48
[2] 高爱农,姜淑荣,赵锡温,等.苹果品种抗寒性测定方法的研究[J].果树科学,2000,17(1):17-21
[3] 牛锦凤,李国,王振平.鲜食葡萄品种抗寒性的比较研究[J].陕西农业科学,2005(6):35-36
[4] 赵蕾,廖康,王瑾,等.野生欧洲李抗寒特性研究初探[J].中国农学通报,2009,25(06):97-100
[5] 盛宝龙,吴伟民.枇杷用电导法测定其花果的抗冻性[J].东南园艺,1996(3):1-5
[6] 徐康,夏宜平,徐碧玉,等.以电导法配合Logistic方程确定茶梅‘小玫瑰’的抗寒性[J].园艺学报,2005,32(1):148-150
[7] 丁灿.电解质渗漏法在植物抗寒性鉴定中的应用[J].思茅师范高等专科学校学报,2001,17(3):8-10
[8] 陈建白.电导法在植物抗寒研究中的应用[J].云南热作科技,1999,22(1):26-28
[9] 田如男,薛建辉.深山含笑和乐昌含笑的抗寒性测定[J].南京林业大学学报,2004,28(6):55-57
[10] 曹青爽.阿月浑子的生长习性及利用价值[J].中国林副特产,2007,10,5(90):95-96
[11] 刘祖琪.植物抗性生理学[M].北京:中国农业出版社,1994
[12] 张军科,桑春果,李嘉瑞,等.杏品种资源抗寒性主成分分析[J].西北农业大学学报,1999,27(6)79-84
ColdResistanceofSeveralMainTreeFruitSpeciesinXinjiangUygurAutonomousRegion
Zhao Lei, Du Yan, Liu Heng, An Lu, Lu Mingyan, Zhang Dongya
(Xinjiang Forestry Academy,Urumqi 830063,China)
Taking annual dormant branches of several common fruit trees in Xinjiang Uygur Autonomous Region as experiment materials,the electrolyte leakage rate was measured under low temperature stress,the Logistic curve fitting was used to find the inflection point temperature and the half lethal temperature as the reference index for the evaluation of cold resistance. Result shows that among nine species plum and Dangshan pear have the strongest cold resistance ,Pistaciachinensishas the weakest cold resistance. Cold resistance from strong to weak is plum>Dangshan pear>Korla fragrant pear>Jujube>Juglansregia‘Zha 343’>Juglansregia‘Xinxin 2’>Juglansregia‘Wen 185’>Pistaciavera>Pistaciachinensisbunge.
fruit tree; cold resistance; semi-lethal temperature(LT50); restoring growth method
1005-5215(2017)10-0024-03
2017-09-11
新疆维吾尔自治区公益性科研院所基本科研业务经费资助项目(KY2017029)
赵蕾(1981-),女,新疆人,硕士,助理研究员,从事果树栽培与育种研究,Email:zhaolei1118@qq.com
张东亚(1964-),男,河南人,硕士,研究员,从事林木育种研究,Email:zdywah998@126.com
S66
A
10.13601/j.issn.1005-5215.2017.10.008
