灰木莲种源幼苗叶片性状表型多样性分析
2017-11-09文珊娜仲崇禄姜清彬李清莹
文珊娜 仲崇禄 姜清彬 陈 羽 张 勇 李清莹
(中国林业科学研究院热带林业研究所,广州 510520)
灰木莲种源幼苗叶片性状表型多样性分析
文珊娜 仲崇禄*姜清彬 陈 羽 张 勇 李清莹
(中国林业科学研究院热带林业研究所,广州 510520)
以越南收集的12个灰木莲(ManglietiaconiferaDandy)种源为材料,对叶片形态性状(叶长、叶宽、叶面积、叶周长、长宽比、叶柄)以及微形态特征(气孔器密度、气孔器长、气孔器宽、气孔器面积、长宽比)进行测定,采用方差分析、变异系数、相关分析和主成分分析等方法进行分析。结果表明:灰木莲不同种源间的叶表型性状存在显著差异;种源中,LC2的叶形态性状的平均变异系数最大(22.09%),TQ2的变异系数最小(12.76%);叶表型性状中,叶面积的变异系数最大(28.60%),气孔器宽的变异系数最小(5.19%);相关性分析结果表明叶表型性状间存在显著或极显著的相关关系,而地理因素中经度与叶周长显著相关,经纬度与叶长宽比呈显著相关,海拔与叶表型性状间的相关性不显著(P<0.05);主成分分析表明前3个主成分的累计贡献率达到了89.29%,基本代表原始性状的全部信息。灰木莲12个种源经聚类分析在欧式距离5阈值处可分为4类。灰木莲种源间的叶表型性状存在着丰富的变异,纬度对灰木莲叶形态特征有明显影响,灰木莲种源间和性状间的变异程度存在着差异,本研究为灰木莲遗传改良提供理论依据。
灰木莲;种源;叶片;表型性状;气孔器
植物的表型性状多样性是遗传物质多样性的具体表现,是遗传多样性与环境多样性的综合表现,表型性状上的变异通常具有适应和进化上的意义[1]。植物叶片的表型特征常用于植物的分类,不同植物的叶片在叶形、叶片颜色、质地、叶缘等存在明显的差异[2],属内植物或同种植物不同种质资源的叶片表型性状会存在较大的差异[3~4]。植物叶表皮的气孔特征是植物种质资源多样性研究的一个重要指标,叶表皮微形态特征的多样性在分类学上也有着重要的意义,通过比较植物叶表皮气孔形态的研究还可以分析气孔形态与地理环境间的关系[5~7]。
灰木莲(ManglietiaconiferaDandy)为木兰科(Magnoliaceae)木莲属(Manglietia)常绿阔叶植物[8],原产于越南及印度尼西亚爪哇地区,树干通直,是优良的速生用材树种,在越南北部和我国华南地区均有广泛种植。灰木莲木材可做家具、建筑用材,也可制作胶合板等。自上世纪60年代以来,我国开始从越南引进灰木莲在广东、广西、福建、云南进行栽培种植,灰木莲生长表现良好。近年来林业科研工作者对灰木莲的研究也日益重视,对灰木莲人工林生态[9~12]、花的繁育特性[13~14]、木材材性[15~16]等方面有较多研究报道,但对灰木莲种质资源的表型性状多样性的研究少见报道。本文通过测量灰木莲12个种源幼苗的叶片性状,利用扫描电子显微镜对叶表皮结构进行观察比较,研究和分析灰木莲不同种源在叶表型性状间的差异,旨在了解灰木莲叶表型性状的变异程度及其与地理环境的关系,为灰木莲种源多样性研究、种源选育与评价、资源利用等奠定基础。
1 材料与方法
1.1 试验材料及试验地点
试验材料为12个种源的灰木莲苗(表1),灰木莲种子于2013年9月引自越南林业科学研究院,采种母树为自由授粉,于2013年10月播种,植株种植于装有黄心土与泥炭土(体积比为3∶1)的容器(10 cm×12 cm)内,期间根据苗木生长情况适时适量浇水和施肥,以保证苗木的正常生长,苗龄1.5年(苗高约70 cm,地径约9 mm)时用于试验。

表1 灰木莲种源信息表
试验地位于广东省广州市中国林业科学研究院热带林业研究所内苗圃,属南亚热带季风气候,年均温约为24℃,年均湿度约为80%,年均降水量约为2 200 mm,海拔在43 m。
1.2 灰木莲叶形态观测
每个种源随机选取生长一致的5株灰木莲,每株采摘一片位于第5~6位生长完整的叶片(自上而下),重复4次,每种源共计20片叶,用手持式激光叶面积仪(CI-203)测量叶片的面积、长、宽、周长及长宽比。
1.3 气孔形态观测
每个种源观测5株为一重复,共3次重复,每株选取一片成熟健康的叶片,取叶中部与主脉平行的位置切成0.8 cm×0.8 cm的小块,用戊二醛固定液固定于离心管中,经脱水干燥后,将叶片粘于金属台上并做标记,经离子溅射镀金后在JSM-6360L扫描电子显微镜下观察、拍照保存。每片叶随机选取10个视野统计气孔数目(400×),计算单位面积气孔数目,并测量气孔器大小(长、宽),气孔器面积=气孔器长×宽。
1.4 数据分析
采用SPSS 22.0分析软件对数据进行均值分析、Duncan’s多重比较分析、Pearson相关性分析和主成分分析,并进行UPGMA聚类分析;用变异系数(CV,%)来衡量叶表型性状的离散程度[4]。
2 结果与分析
2.1 叶片形态性状分析
由表2可知,灰木莲种源间在叶面积、叶长度、叶宽度、叶周长、长宽比及叶柄长等叶形态表型性状均存在着显著差异(P<0.05)。灰木莲12个种源的平均叶面积为58.25 cm2,叶长平均为19.45 cm,叶宽平均为5.15 cm,叶周长平均为45.94 cm,叶柄长平均为1.45 cm,灰木莲叶长宽比平均为3.79。叶面积指标中,不同种源间叶面积大小存在显著差异,种源TQ1最大为68.41 cm2,其次是种源TQ2为67.31 cm2,种源BK1、LC2和LC3叶面积相对较小,彼此间没有显著差异,其中,种源LC2的最小为46.46 cm2;叶长较长的种源有TQ1、TQ2和YB2,较小的为BK2、LC2和LC3,所有种源中叶长最大的为种源TQ1,最小的为种源BK2,分别为23.72和16.47 cm;灰木莲叶片宽度在12个种源间也表现出显著差异,最大的为种源LC4(5.54 cm),其次是种源LC1(5.52 cm),LC4和LC1的叶宽与种源TQ1、TQ2、TQ3无显著差异,与其余种源存在着显著差异,LC3的叶宽最小(4.68 cm),其与种源PT1、YB1、YB2、BK1、BK2和LC2之间没有显著差异;种源TQ1的叶片周长最大,其次是BK1和YB2,分别为49.96、49.88和49.76 cm,最小的为种源BK2,叶片周长为39.26 cm;叶柄长指标中,除种源LC4外,种源TQ1的叶柄为最长(1.88 cm),其次种源LC4为1.68 cm,叶柄长较小的种源有LC2和LC3,分别为1.20和1.24 cm。
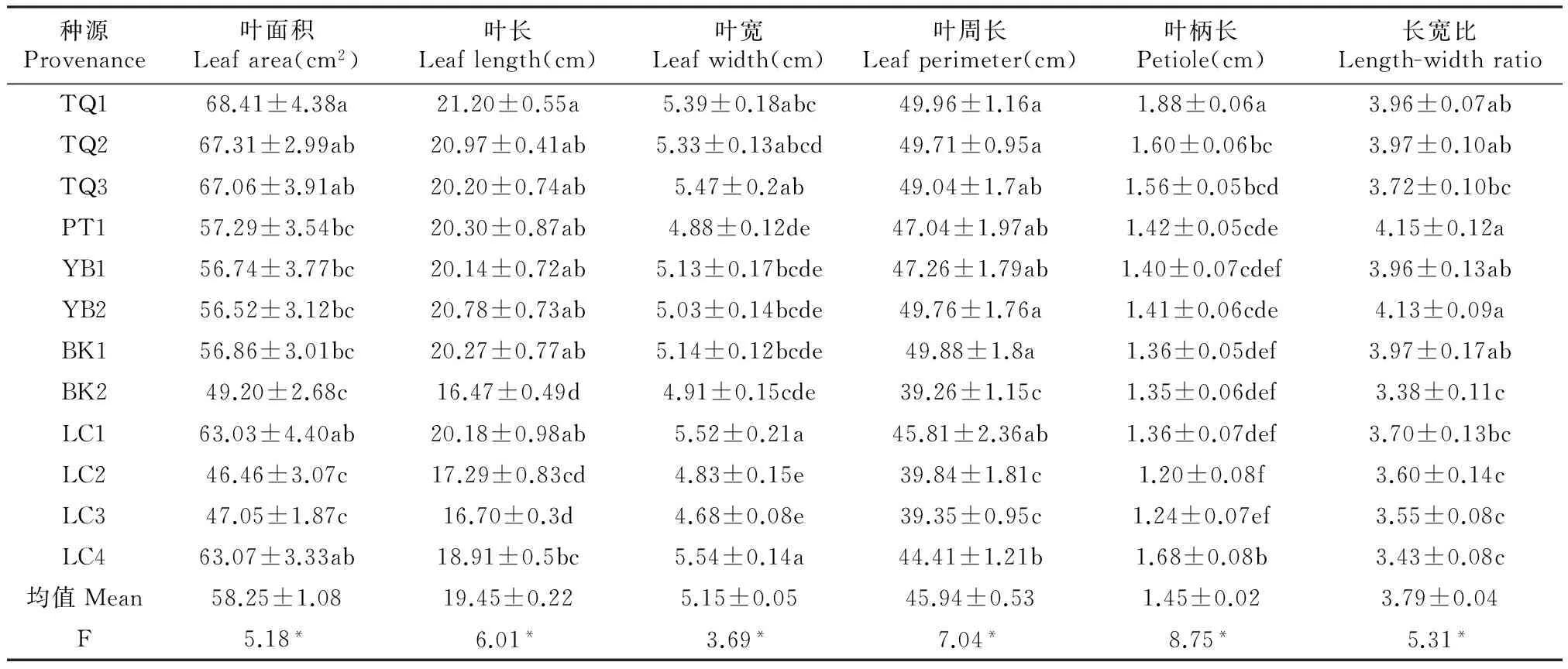
表2 灰木莲不同种源叶片形态表型性状分析
注:表格中数据为均值±标准误,同列不同字母表示种源间存在显著差异(P<0.05),*表示在0.05水平上存在显著差异。下同。
Note:The data in the table were mean±SE,the different small letters at the same column indicate significant difference at the 0.05 level,*means significant difference at the 0.05 level.The same as below.
叶片长宽比是反映叶形的一个重要指标,种源PT12的叶长宽比最大为4.15,与种源TQ1、TQ2、YB1、YB2和BK1之间没有显著差异,显著大于其他种源,种源BK2、LC4和LC3的叶长宽比相对较小,分别为3.38、3.43和3.55。可见,叶片形态表型性状叶面积、叶长、叶宽、叶周长、叶柄长和长宽比各指标中,种源TQ1和TQ2的各指标值均较大,而较小的为种源BK2、LC2和LC3。种源TQ1与BK2、LC2、LC3在叶片的形态性状存在显著差异。
对灰木莲12个种源叶的形态性状的变异程度进行分析(表3),叶面积、叶长、叶宽、叶周长、叶柄长及其长宽比的变异系数幅度分别为17.79%~31.25%、8.85%~21.81%、7.66%~16.80%、8.52%~23.06%、13.97%~29.05%、7.67%~18.96%,其平均变异系数依次为28.60%、17.60%、14.06%、17.71%、23.47%、14.79%,灰木莲叶形态性状的变异系数大小依次为:叶面积>叶柄长>叶周长>叶长>叶长宽比>叶宽,这表明灰木莲叶形态各性状在种源间有着一定程度的变异。种源LC1的叶面积和叶片长度变异幅度最大,分别为31.25%、21.81%,种源LC1的叶宽和周长的变异幅度最大,分别为16.80%、23.06%,种源LC2的叶柄长变异幅度最大(29.05%),种源BK1的叶长宽比的变异幅度为最大(18.96%),种源TQ2的叶长和叶周长的变异幅度最小,分别为8.85%、8.52%;种源LC3的叶面积和叶片宽度的变异幅度均为最小,分别为17.79%、7.66%,TQ1的叶柄长的变异幅度最小(13.97%)。
表3灰木莲不同种源叶片形态性状变异系数(%)
Table3CoefficientofvariationofdifferentprovenancesofM.conifera

种源Provenance叶面积Leafarea叶长Leaflength叶宽Leafwidth叶周长Leafperimeter叶柄长Petiole长宽比Length-widthratio均值MeanTQ128.6611.5315.0210.3913.977.6714.54TQ219.908.8510.678.5216.8811.7712.76TQ326.0716.3316.3015.5215.5812.5217.05PT127.6419.1610.9018.7016.1313.4717.67YB129.7115.9914.6716.9022.3714.5919.04YB224.7215.7412.4415.7820.439.7316.47BK123.6517.0610.2316.1417.6318.9617.28BK224.4013.4013.4813.1121.4714.6216.75LC131.2521.8116.8023.0622.9315.6221.91LC229.5321.5914.2020.3029.0517.8522.09LC317.799.917.6610.8225.379.8613.57LC423.6411.7311.6612.1922.0710.7615.34均值Mean28.6017.6014.0617.7123.4714.7919.37
12个灰木莲种源中,种源LC2和LC1叶表型性状的总体变异系数较大,其变异系数分别为22.09%、21.91%,这表明这2个种源的叶形态多样性比其余种源要丰富一些;而种源TQ2的变异系数最小,为12.76%,说明TQ2种源内个体的叶形态性状差异相对较小。
2.2 气孔形态分析
通过扫描电镜观察灰木莲不同种源的气孔显微形态,观察灰木莲叶片下表皮(图1)和上表皮(图2)可知:灰木莲的叶片下表皮分布有气孔器和表皮毛,上表皮没有表皮毛和气孔器的分布;下表皮细胞形状呈不规则形,细胞排列紧密;灰木莲的气孔器类型为平列型,气孔器呈椭圆形,不规则排列;上下表皮细胞均可见白色晶体物质的分布。灰木莲上表皮细胞多近似为四边形,垂周壁较平直或浅波状弯曲,其中,种源TQ1、TQ2、TQ3、BK2、LC1、LC2、YB1的上表皮垂周壁较平直,种源PT1、YB2的上表皮细胞垂周壁为不明显波状弯曲,BK1、LC3、LC4的上表皮细胞的垂周壁呈浅波状弯曲;灰木莲12个种源的下表皮细胞垂周壁呈浅波状弯曲。
单因素方差分析结果显示(表4),灰木莲不同种源间在叶片下表皮的气孔密度、气孔器大小(长、宽)、气孔长宽比等性状上存在着显著差异。灰木莲叶片气孔器密度平均为127.38个·mm-2;灰木莲的气孔器长平均为30.50 μm,其中,种源LC2的气孔器长最小,为29.16 μm,种源LC4的气孔器长最大,为31.96 μm;气孔器宽平均为22.12 μm,最大值为23.09 μm(LC4),最小值为21.50 μm(LC2)。气孔器的平均面积大小为529.18 μm2,其变异幅度477.59(LC2)~579.61 μm2(LC4);气孔的长宽比均值为1.38,其变幅为1.32~1.43,种源BK2气孔器的长宽比较小(1.32),YB2的长宽比较大(1.43)。种源TQ1、TQ2、TQ3与种源LC4在气孔器密度及其长、宽等形状均存在着显著差异;种源BK2在气孔器密度、气孔器长、气孔器面积及其长宽比等形状与LC4存在着显著差异。
对气孔器各性状进行变异系数分析可知(表4),各性状变异系数大小排列为:气孔器密度(27.8%)>气孔器面积(9.67%)>气孔器长(5.56%)>气孔器长宽比(5.20%)>气孔器宽(5.19%)。其中,气孔器长、气孔器宽、气孔器面积及其长宽比的变异系数都小于10%,可见气孔器大小的变异幅度较叶片大小的变异幅度小,受外界生态环境的影响小于叶片大小形态。

表4 灰木莲不同种源气孔器大小分析

图1 灰木莲12种源叶片下表皮形态(×400)Fig.1 Leaf abaxial epidermis morphology of 12 provenances of M.conifera(×400)

图2 灰木莲12种源叶片上表皮形态(×150)Fig.2 Leaf upper epidermis morphology of 12 provenances of M.conifera(×150)
2.3 叶表型性状的相关性分析
对灰木莲叶形态性状如叶面积、叶长、叶宽等和叶片表皮显微特征(气孔器密度、气孔器大小等)共11个性状指标进行了Pearson相关分析(表5)。结果表明:气孔器密度与气孔器长呈显著负相关;气孔器长、宽与叶面积、叶长、叶宽、叶柄长、叶周长、叶长宽比等6个性状无显著相关关系;气孔器长宽比与叶长宽比间存在显著相关关系;叶长与其余6个叶的形态性状均存在显著相关。
进一步分析叶表型性状与地理因子之间的相关关系,可知:经度与叶周长、气孔器长、气孔器宽、气孔器面积呈负相关,与叶周长和叶长宽比呈显著正相关;纬度与叶宽呈正相关,而与其他性状呈负相关,且与叶长宽比呈显著负相关;海拔与叶表型性状的相关性都不显著,且多为负相关关系。灰木莲与地理因子间的相关关系说明长期生长的生态环境对灰木莲的表型性状会产生一定的变异,对灰木莲的叶片性状产生一些影响。
2.4 叶表型性状主成分分析
利用主成分分析方法对灰木莲叶表型形态的11个性状进行分析可知(表6),第一主成分(y1)的贡献率占40.896%,第二主成分(y2)的贡献率占28.876%,第三主成分(y3)的贡献率占19.515%,前3个主成分的累计贡献率为89.287%,损失10.713%的信息,选取前3个主成分即能够基本反映所有变量信息,可将11个性状变量归类为3个新的变量(y1-y3)。叶长(x2)、叶周长(x4)和叶面积(x1)在第一主成分有较高的载荷,叶宽、叶长宽比及叶柄长的载荷量都大于0.6,信息的载荷量较大,这说明第一主成分基本反映了叶形态性状的信息,第一主成分是叶长、叶周长和叶面积的综合因子;气孔器面积、气孔器长和气孔器宽在第二主成分有较高的载荷,这说明第二主成分基本反映了气孔器大小形态的信息,第二主成分是气孔器面积和气孔器长宽的综合因子;气孔器长宽比、叶长宽比和叶柄长在第三主成分有较高的载荷量。所以,灰木莲种源间的表型差异主要表现在叶长、叶周长和叶面积,其次是气孔器面积和气孔器长宽。

表5 灰木莲叶表型性状相关性分析
注:*表示在0.05水平上显著相关;**表示在0.01水平上显著相关。
Note:*means significantly correlated at the 0.05 level;**means highly significant correlated at the 0.01 level.
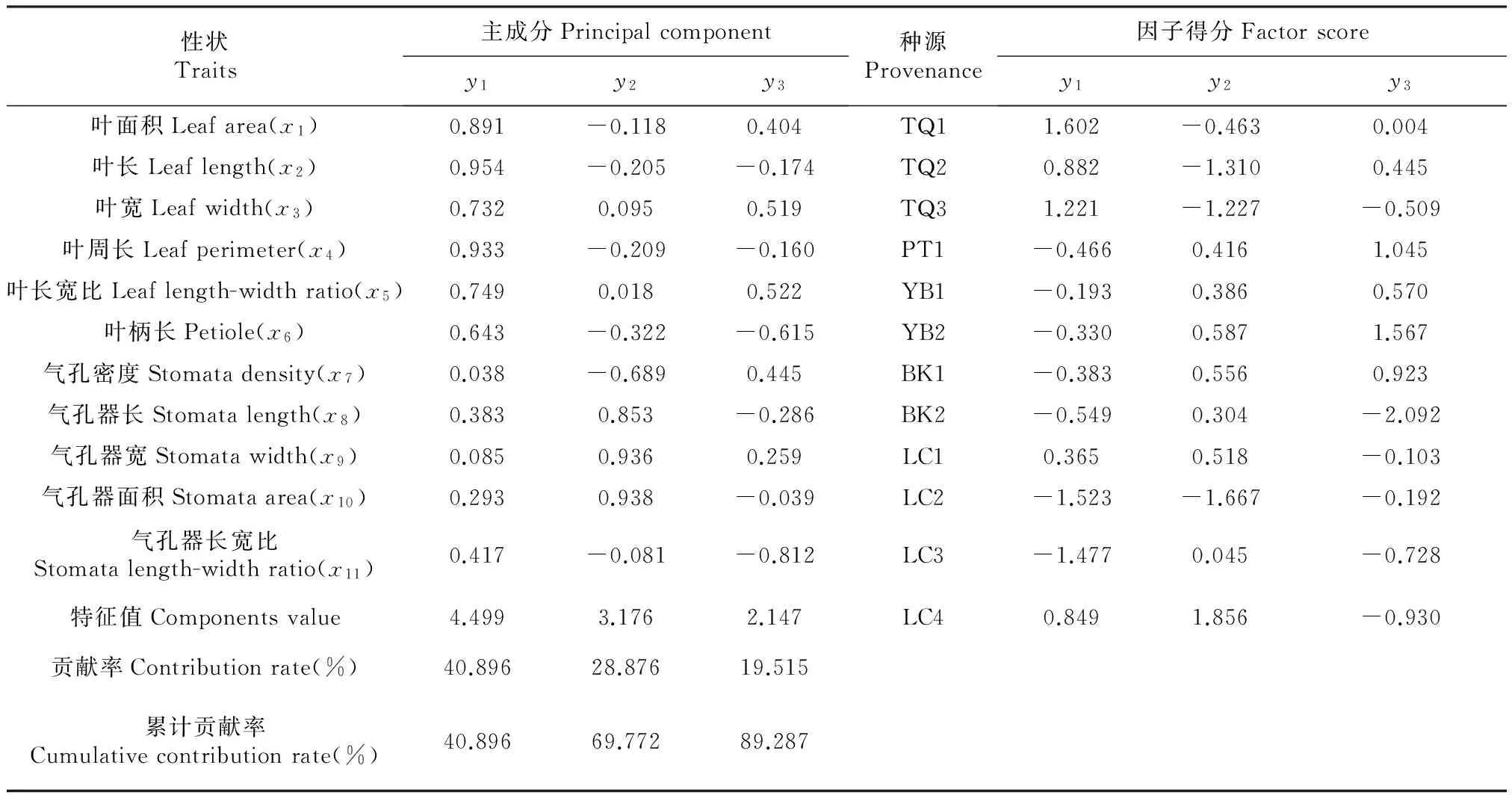
表6 灰木莲叶表型11个性状主成分分析结果
种源TQ1、TQ2、YB2在第一主成分中有较高的正因子得分,而BK2、LC2和LC3在第一主成分中有较高的负因子得分,说明TQ1、TQ2、YB2在第一主成分中的值大于平均值,而BK2、LC2和LC3的值要小于平均值,表明种源TQ1、TQ2、YB2的叶片明显大于BK2、LC2和LC3。同理,种源TQ2、TQ3和LC2在第二主成分有较高的负因子得分,LC4和BK2有较高的正因子得分,说明种源LC4和BK2的气孔器明显大于TQ2、TQ3和LC2。种源YB2和PT1第三主成分中有较大的正因子得分,而BK2和LC4有较大的负因子得分,种源YB2和PT1具有较大的叶长宽比及气孔器长宽比。
2.5 不同种源叶表型性状聚类结果分析
以上述灰木莲的11个叶表型性状作为变量,采用Ward方法对灰木莲12个种源进行聚类分析(图3),以欧式距离为5时,灰木莲12个种源可划分为4大类,即:第一类包括PT1、YB1、BK1、YB2等4个种源,这一大类种源的叶片大小和气孔分布密度均为中等,气孔器的大小相对较大;第二大类包括LC1和LC4两个种源,这一类群具有大叶片和较大的叶长宽比值,气孔分布密度相对小,具有较大的气孔器;第三大类包括TQ1、TQ2和TQ3这三个种源,均来自同一省份,叶片相对较大,气孔密度也相对较大;第四大类包括BK2、LC3和LC2共3个种源,叶片相对较小,叶柄长相对较短,气孔分布密度大。
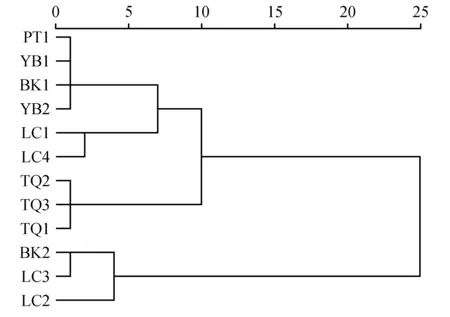
图3 灰木莲叶表型性状聚类分析结果Fig.3 The cluster analysis result of leaf phenotypic traits of M.conifera
3 讨论
植物的变异与自身的遗传特性及生长环境密切相关,一般情况下,树种分布范围越大则变异大,分布的范围小则变异也越小[17]。灰木莲的叶表型性状在12个不同种源间产生了显著的差异,以叶片长和宽为例,种源TQ1的叶片平均长和宽都大于其他种源,种源TQ1的叶片在所有种源中是最大的。种源LC2的叶片表型变异程度在种源中为最大,TQ2的变异程度为最小,同一种源在不同性状间的变异程度也不一样,说明灰木莲种源间和种源内个体间在叶片表型性状变异程度存在不均衡现象,性状具有丰富的变异,这是灰木莲种源选择的基础。灰木莲苗期叶片表型性状也存在着不同程度的变异,其中叶面积和气孔器密度的变异程度相对较大,气孔器宽的变异幅度最小,说明在相同生境下同一性状受到的影响是不一样的,或是不同性状在同一生境下的反应是不一样的,这可能是由于与灰木莲本身的遗传因素有关,也与环境因子的影响相关[18]。
灰木莲叶表型性状间具有不同的相关性,气孔器长宽比与叶片大小形态有着显著或极显著差异,叶片的大小一定程度上能够影响着气孔器的形状。通过种源苗木叶片表型性状与产地气候因子的相关分析揭示,产地经纬度与大部分性状的相关性不显著,叶片的长宽比与经纬度显著相关,叶周长与经度显著相关,经纬度对灰木莲不同种源的叶表型变异具有一定的影响。木荷(Schimasuperba)不同种源苗木的的表型性状表现为纬向变异模式,而温度是造成这一变异模式的主要环境作用因子[19],温度对灰木莲的生长也有较大地影响,在平均最低气温较低(-5℃以下)的地区会出现冻害或冻死[20~21],根据叶片表型性状可以帮助选择抗寒的优良种质资源,如种源BK2的叶长宽比最小,而PT1的最大,BK2可能在平均气温较低的地区的适应性更强。有研究报道,植物的气孔器特征会随着纬度或海拔的变化而发生不同的变化[22~23],本次研究中不同种源叶表皮气孔形态与采集地的经纬度、海拔等地理因子没有表现出明显的相关性,因此,判断灰木莲植株的叶表皮气孔形态与地理因子间的相关性还有待进一步地研究。
植物能够通过气孔吸收大气中的污染物如SO2、Cl2及重金属,叶片的结构特征对大气污染物的吸收和滞尘能力有着直接的影响[24~25],有研究显示沈阳地区绿化树种的叶面积与大气污染物中的Cl、Pb、Cr显著正相关,气孔密度与Hg、S有着极显著负相关[25]。有研究结果显示,灰木莲在高浓度SO2和氟化物大气污染环境下的抗性为中等[26~27],还有着较好杀菌的效果[28~29]。灰木莲叶面积较大的种源(TQ1、TQ2和TQ3)相对于叶面积小的种源(LC2、LC3和BK2)可能能吸收更多的大气污染物中的Cl、Pb、Cr等物质,气孔密度低的种源如BK1、LC4、LC1可能对Hg和SO2的抗性强于气孔密度高的种源(TQ3、TQ1、TQ2),种源TQ1、TQ2、TQ3可能对大气污染物中的Cl、Pb、Cr等物质的抗性强于Hg和SO2。了解灰木莲不同种源的叶片结构特征,有助于灰木莲在华南地区的城市森林景观树种配置中的应用,进一步筛选抗污染性能较好的材料。
1.牟洪香,侯新村,刘巧哲.木本能源植物文冠果的表型多样性研究[J].林业科学研究,2007,20(3):350-355.
Mu H X,Hou X C,Liu Q Z.Study on the phenotype diversity of woody energy plantXanthocerassorbifolia[J].Forest Research,2007,20(3):350-355.
2.Beghin T,Cope J S,Remagnino P,et al.Shape and texture based plant leaf classification[M].//Blanc-Talon J,Bone D,Philips W,et al.Advanced concepts for intelligent vision systems.Berlin Heidelberg:Springer,2010,64:345-353.
3.贾春红,张利,魏晓,等.川西地区毛叶木姜子的表型多样性研究[J].林业科学研究,2015,28(6):844-850.
Jia H C,Zhang L,Wei X,et al.Phenotypic polymorphism ofLitseamollisHemsl in West Sichuan province[J].Forest Research,2015,28(6):844-850.
4.朱利利,吴疆翀,彭兴民,等.印楝属植物表型变异分析[J].林业科学研究,2016,29(2):162-166.
Zhu L L,Wu J C,Peng X M,et al.Phenotypic difference among species and a variation type ofAzadirachta[J].Forest Research,2016,29(2):162-166.
5.唐自慧,逄云莉,何兴金,等.鹿药属植物叶表皮特征及其系统学意义[J].武汉植物学研究,2007,25(6):550-557.
Tang Z H,Pang Y L,He XJ,et al.Characters of leaf epidermis and their systematic significance inSmilacina[J].Journal of Wuhan Botanical Research,2007,25(6):550-557.
6.刘静,王磊,邢怡,等.黑龙江苹果亚科植物叶表皮形态结构的研究[J].电子显微学报,2014,33(1):69-76.
Liu J,Wang l,Xing Y,et al.Morphology structure of leaf epidermis of Maloideae in Heilongjiang[J].Journal of Chinese Electron Microscopy Society,2014,33(1):69-76.
7.谢果珍,舒少华,王沫.不同居群栝楼叶表皮气孔形态研究[J].时珍国医国药,2008,19(12):2956-2960.
Xie G Z,Shu S H,Wang M.Study on the stomatal morphology of different geographical populations ofTrichosantheskirilowiiMaxim[J].Lishizhen Medicine and Materia Medica Research,2008,19(12):2956-2960.
8.Pham H H.Manglietia conifera[M].//An illustrated flora of Vietnam,Vol.I.Ho Chi Minh.Vietnam:Young Publishing House,1999:230.
9.李俊贞,秦武明,覃毓,等.灰木莲人工林生物量和生产力的研究[J].福建林业科技,2011,38(1):1-5.
Li J Z,Qin W M,Qin Y,et al.Studies on the Biomass and Productivity ofManglietiaglauca[J].Journal of Fujian Forestry Science and Technology,2011,38(1):1-5.
10.覃祚玉,何斌,韦录,等.灰木莲人工林碳贮量及其分配特征[J].林业科技开发,2013,27(5):33-36.
Qin Z,He B,Wei L,et al.Carbon storage and distribution ofManglietiaglaucaplantation[J].China Forestry Science and Technology,2013,27(5):33-36.
11.黄承标,韦善华,秦武明,等.不同年龄灰木莲人工林对林内小气候的影响[J].西南大学学报:自然科学版,2012,34(11):95-99.
Huamg C B,Wei S H,Qin W M,et al.Influences ofManglietiaglaucaplantations of different ages on the microclimate in them[J].Journal of Southwest Agricultural University,2012,34(11):95-99.
12.戴军,何斌,肖继谋,等.灰木莲人工林营养元素分配及其积累特征[J].林业科技开发,2012,26(4):39-42.
Dai J,He B,Xiao J M,et al.Accumulation and distribution characteristics of nutrient elements onManglietiaglaucaplantation[J].China Forestry Science and Technology,2012,26(4):39-42.
13.招礼军,韦善华,朱栗琼,等.灰木莲的开花特性及繁育系统的研究[J].西部林业科学,2015,44(2):24-28,35.
Zhao L J,Wei S H,Zhu L Q,et al.Flowering characteristics and breeding system ofManglietiaglauca[J].Journal of West China Forestry Science,2015,(2):24-28.
14.姜清彬,文珊娜,仲崇禄,等.灰木莲开花结实生物学观察[J].西南农业学报,2016,29(9):2229-2233.
Jiang Q B,Wen S N,Zhong C L,et al.Observation of flowering biology and fruit set inManglietiaconiferaDandy[J].Southwest China Journal of Agricultural Sciences,2016,29(9):2229-2233.
15.韦善华,唐天,符韵林,等.灰木莲树皮率、心材率及木材密度研究[J].西北林学院学报,2011,26(3):152-155.
Wei S H,Tang T,Fu Y L,et al.Bark Percentage,heartwood percentage and density forManglietiaglauca[J].Journal of Northwest Forestry University,2011,26(3):152-155.
16.陈松武,梁炜文,许彩娟,等.人工林灰木莲木材物理力学性质研究[J].安徽农业科学,2012,40(16):8993-8996.
Chen S W,Liamg W W,Xu C J,et al.Study on the physical and mechanical properties ofManglietiaglauca[J].Woods Journal of Anhui Agricultural Sciences,2012,40(16):8993-8996.
17.姜荣波,刘军,姜景民,等.红楠主要表型和苗期性状地理种源变异[J].东北林业大学学报,2011,39(5):9-11,23.
Jiang R B,Liu J,Jiang J M,et al.Geographical variation in main phenotypic traits and seedling traits ofMachilusthunbergii[J].Journal of Northeast Forestry University,2011,39(5):9-11,23.
18.解新明,云锦凤,卢小良,等.蒙古冰草表型数量性状的变异与生境间的相关性[J].生态学杂志,2003,22(4):31-36.
Xie X M,Yun J F,Lu X L,et al.Correlation analysis between the variation of phenotypic quantitative characters and the habitat ofAgropyronmongolicum[J].Chinese Journal of Ecology,2003,22(4):31-36.
19.张萍,金国庆,周志春,等.木荷苗木性状的种源变异和地理模式[J].林业科学研究,2004,17(2):192-198.
Zhang P,Jin G Q,Zhou Z C,et al.Provenance difference and geographic variation pattern for seedling trait ofSchimasuperba[J].Forest Research,2004,17(2):192-198.
19.徐大平,丘佐旺.南方主要珍贵树种栽培技术[M].广州:广东科技出版社,2013:57-70.
Xu D P,Qiu Z W.Cultivation techniques of valuable tree species in South China[M].Guanzhou:Guangdong Science and Technology Publishing House,2013:57-70.
21.杨耀海,王代艳.灰木莲大苗培育及山地造林对比试验[J].林业建设,2010(3):15-18.
Yang Y H,Wang D Y. Contrast test on cultivation of nursery stock and hill afforestation forManglietiaglauca[J].Forestry Construction,2010(3):15-18.
22.周浩.不同地理条件下紫椴亚显微结构比较研究[D].哈尔滨:东北林业大学,2008.
Zhou H.Comparative study on ultrastructure ofTiliaamurensisin different geograghical conditions[D].Harbin:Northeast Forestry University,2008.
23.马书荣,阎秀峰,陈柏林,等.不同海拔裂叶沙参和泡沙参气孔形态的对比研究[J].东北林业大学学报,1999,27(6):94-97.
Ma S R,Yan X F,Chen B L,et al.Comparative study on stomatal morphology ofAdenophoralobophyllaandA.potaniniiin different altitude[J].Journal of Northeast Forestry University,1999,27(6):94-97.
24.李汉卿,谢文焕,傅纯彦,等.环境污染与植物[M].哈尔滨:黑龙江科学技术出版社,1998:210-229.
Li H Q,Xie W H,Fu C Y,et al.Environmental pollution and plants[M].Harbin:Heilongjiang Press of Science and Technology,1998:210-229.
25.魏忠平,刘红民,陈罡,等.沈阳地区11种绿化树种的叶结构特征及其对大气污染物的积累特性[J].沈阳农业大学学报,2015,46(6):678-683.
Wei Z P,Liu H M,Chen G,et al.Leaf structure and accumulating capacity of air pollutants of 11 virescence tree species in Shenyang[J].Journal of Shenyang Agricultural University,2015,46(6):678-683.
26.孔国辉,陆耀东,刘世忠,等.大气污染对38种木本植物的伤害特征[J].热带亚热带植物学报,2003,11(4):319-328.
Kong G H,Lu Y D,Liu S Z,et al.Injury symptoms of 38 woody species exposed to air pollutants[J].Journal of Tropical And Subtropical Botany,2003,11(4):319-328.
27.刘世忠,薛克娜,孔国辉,等.大气污染对35种园林植物生长的影响[J].热带亚热带植物学报,2003,11(4):329-335.
Liu S Z,Xue K N,Kong G H,et al.Effects of air pollution on the growth of 35 garden plants[J].Journal of Tropical And Subtropical Botany,2003,11(4):329-335.
28.谭家得.珠三角陶瓷工业污染区耐污染树种选择研究[D].北京:中国林业科学研究院,2009:46-56.
Tan J D.Study on selection of Pollution-tolerant species in the ceramic industrial polluted area,Pearl River Delta Region[D].Beijing:China Academy of Forestry,2009:46-56.
29.胡羡聪,吴小英,温海祥,等.珠三角城市森林景观树种杀菌效应及其应用[J].中国城市林业,2005,3(6):46-49.
Hu X C,Wu X Y,Wen H Y,et al.Effects on bacteria control of urban ornamental trees and application in Pearl River Delta Region[J].Journal of Chinese Urban Forestry,2005,3(6):46-49.
Special Fund for Forest Scientific Research in the Pubil Welfare(201304113-01)
introduction:WEN Shan-Na(1989—),female,Ph.D candidate,mainly engaging in forest tree genetics and breeding.
date:2016-11-07
PhenotypicDiversityAnalysisofSeedlingLeafTraitsofManglietiaconiferaDandy
WEN Shan-Na ZHONG Chong-Lu*JIANG Qing-Bin CHEN Yu ZHANG Yong LI Qing-Ying
(Research Institute of Tropical Forestry,Chinese Academy of Forestry,Guangzhou 510520)
It studied the phenotypic variation in the leaves ofManglietiaconiferawith 12 provenances seedlings from Vietnam, and measured the leaf morphological traits(length, width, area, perimeter, length-width ratio, petiole length) and micro morphological characteristics of stomata(density, length, width, area, length-width ratio) by the variance analysis, variable coefficient(CV), correlation analysis and principal components analysis(PCA). There were significant differences in leaf phenotypic traits amongM.coniferaprovenances. The average CV of leaf morphological traits of provenance LC2 was the maximum(22.09%), and TQ2 was the minimum(12.76%), leaf area recorded the largest CV(28.60%) while stomata width recorded the smallest CV(5.19%). The result of correlation analysis showed that there was significant correlation between leaf phenotypic traits(P<0.05), leaf perimeter was significantly correlated with longitude, and leaf length-width ratio were significantly correlated with longitude and latitude(P<0.05), leaf phenotypic traits were not significantly correlated with longitude and altitude)(P<0.05). By PCA, the cumulative contribution rate of the first 3 principal components was 89.29%, basically represent all the information of the original character. By cluster analysis, the 12 provenances could be clustered into 4 groups at the Euclidean distance of 5. There is abundant variation of leaf phenotypic traits inM.coniferaprovenances, longitude and latitude have obvious influence on leaf morphological characteristics ofM.conifera, and there is difference of variation degree among provenances and leaf phenotypic traits ofM.conifera. Our study provides theoretical basis for genetic improvement ofM.conifera.
ManglietiaconiferaDandy;provenance;leaf;phenotypic traits;stomata
林业公益性行业科研专项“灰木莲种源家系选育”(201304113-01)
文珊娜(1989—),女,博士研究生,主要从事林木遗传育种研究。
* 通信作者:E-mail:zclritf@21cn.com
2016-11-07
* Corresponding author:E-mail:zclritf@21cn.com
Q949.747.1
A
10.7525/j.issn.1673-5102.2017.02.018
