根据广义形态性状和nrDNA ITS序列试论长颖固沙草的归并
2017-11-09刘玉萍周勇辉张晓宇
刘玉萍 吕 婷 周勇辉 张晓宇 苏 旭
(青海师范大学生命与地理科学学院,青藏高原药用动植物资源重点实验室,青海省自然地理与环境过程重点实验室,青藏高原环境与资源教育部重点实验室,西宁 810008)
根据广义形态性状和nrDNAITS序列试论长颖固沙草的归并
刘玉萍 吕 婷 周勇辉 张晓宇 苏 旭*
(青海师范大学生命与地理科学学院,青藏高原药用动植物资源重点实验室,青海省自然地理与环境过程重点实验室,青藏高原环境与资源教育部重点实验室,西宁 810008)
固沙草属是禾本科中一个具有重要经济价值的高山特有属,共记载6个物种,它们均具有较强的抗逆特性,是农业良种繁育、畜牧业牧草改良利用的重要资源;并且,固沙草属物种形态差异不显著,种间界限较模糊。本研究通过广义形态性状的观测、统计和分析,同时结合核基因组ITS片段的研究,系统探讨了长颖固沙草、固沙草和鸡爪草3个近缘物种的主要形态性状和分子序列的差异,旨在评判长颖固沙草的系统分类归属。研究表明,在广义形态性状上长颖固沙草与固沙草差异甚小,难以进行区分,但与鸡爪草差异明显,是典型的种间关系;从基于ITS序列构建的系统发育树看,3个物种中只形成两个比较明显的遗传分支,一个分支是由长颖固沙草和与固沙草所测个体构成,另一分支仅由鸡爪草所测个体组成,并且分支的靴带支持率高达100%。故本研究认为,长颖固沙草与固沙草应是同一个物种,长颖固沙草应作为固沙草的异名处理。
禾本科;固沙草属;长颖固沙草;形态性状;ITS序列
长颖固沙草[1]是在2009年《Annales Botanici Fennici》上发表的禾本科(Poaceae)、三齿稃亚族(Tridentinae)、固沙草属(Orinus)一新种,即OrinuslongiglumisL.B.Cai & X.Su,主要分布于中国西藏南部的乃东县、朗县和加查县,生于海拔3 040~3 650 m的山坡、草原、河滩、湖岸及林缘。它自发表以来,一直没有得到国内外禾本科分类学学者的普遍接受和认可,进而导致其形态变异至今无人做过细致考证,那么该种究竟是否是一个独立的物种?其与属内其余物种的亲缘关系如何?这些科学问题迄今无人问津。事实上,长颖固沙草与属内近缘物种间性状差异甚小,种间界限不明确,这也往往造成分类学家对其野外辨别困难或室内鉴定张冠李戴。在过去的5年内,我们调查了长颖固沙草在青藏高原的分布,并收集了大量群体标本、DNA材料和种子,发现长颖固沙草的那些性状即使在该种同一居群中也都存在相当大的变异,且其与近缘物种固沙草间也没有明显差异存在,可能隶属于同一个物种。据此,本研究在广义形态性状特征对比基础上,结合核基因组ITS片段的研究,对该种进行分类处理。
1 材料和方法
1.1 对比类群
要考证长颖固沙草的形态变异及其与近缘物种的性状差异,首先一个重要的程序是要找出与该种至少在外部形态上相似的相近类群。由于原始描述[1]认为长颖固沙草与固沙草(O.thoroldii(Stapf ex Hemsl.) Bor)接近,而固沙草与鸡爪草(O.anomalaKeng ex Keng f.et L.Liou)关系极为密切,故选择固沙草和鸡爪草作为长颖固沙草的参照和对比类群。
1.2 实验材料
选取长颖固沙草、固沙草和鸡爪草3个物种、31个居群作为研究材料,其涵盖研究对象主要的地理分布范围。实验材料主要用于形态学性状的观察、测量和统计分析,叶表皮微形态性状的研究以及分子序列的分析。此外,每个个体的新鲜健康叶片采集后迅速用硅胶干燥处理以便日后的DNA提取。其各份材料的名称及来源详见表1。
1.3 研究方法
1.3.1 形态特征分析
对3个物种、31个居群分别进行植株高度、根状茎是否明显、旗叶长度、旗叶宽度、叶片被毛状况、叶鞘被毛状况、圆锥花序长度、圆锥花序分枝数、植株分节数、小穗数、小穗长度、每小穗小花数、小穗颜色、颖体长度、颖脉数、颖片被毛状况、第一外稃长度、外稃被毛状况、第一内稃长度、内稃被毛状况等21个性状的观察和测量。每个代表居群取3~5株观察值的平均数,再将每个物种不同居群的观察资料平均,以对各个物种进行相互比较;同时,对分类价值较大的外部形态数量性状矩阵应用SPSS软件进行主成分分析(Principal component analysis,PCA)。
1.3.2 叶表皮微形态观察
材料处理和操作方法,请参阅苏旭等[2]方法。长细胞测其绝对长度,每个代表物种取3个视野,每个视野取10个细胞,取平均值;气孔器大小测其长宽比,每个代表种取3个视野,每个视野取5个气孔器,取平均值。叶表皮特征用Metcalfe[3]的微形态术语进行描述。
1.3.3DNA提取、PCR扩增、序列比对及系统发育树构建
硅胶干燥的叶片采用改进的CTAB法[4]提取总DNA。以总DNA作为模板,引物ITS1和ITS4[5]对ITS区域(包括ITS1、5.8S和ITS2)进行PCR扩增,扩增产物用于测序反应,应用ABI3730自动测序仪进行测序;然后,应用Clustal X 1.81[6]和MEGA 5[7]对ITS序列进行比对并加以手工校对;最后,应用PAUP*4.0b10[8]和MrBayes 3.1.2[9]分别构建分子系统发育树,并检验它们之间的遗传差异,同时给出重复1 000次各分支的自展支持率BV(bootstrap value)值和贝叶斯系统树(Bayesian inference,BI)的后验概率PP值。
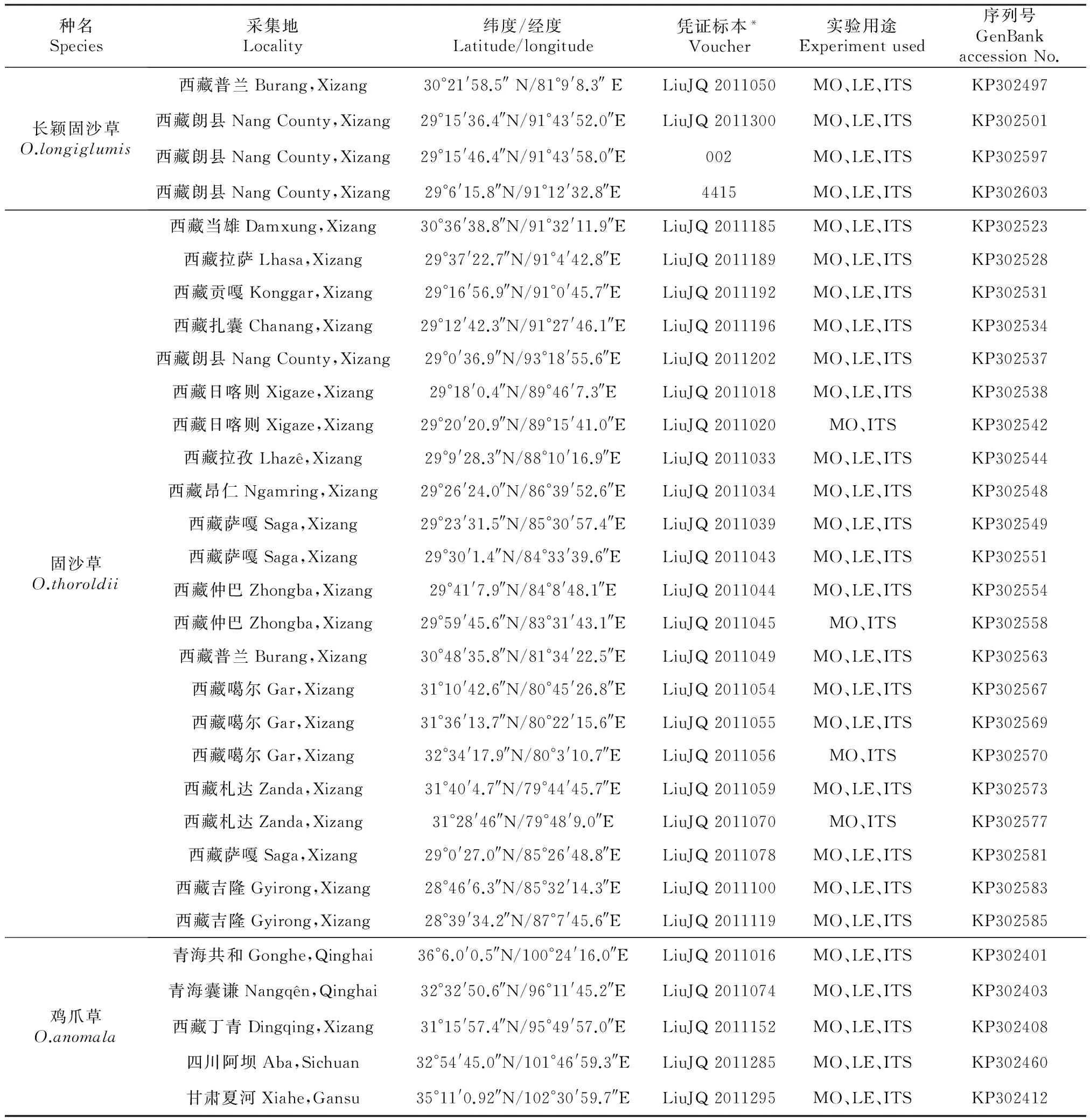
表1 材料名称及来源
注:*凭证标本存放于中国科学院西北高原生物研究所青藏高原生物标本馆(QTPMB);MO.形态学观察;LE.叶表皮观察;ITS. nrDNA ITS测序分析
Note:A*Vouchers are conserved in QTPMB; MO. Used for morphological observation; LE.Used for leaf epidermal observation; ITS.Used for nuclear rDNA ITS sequencing
2 结果与分析
2.1 外部形态性状差异分析
通过对固沙草属3个物种、31个居群、21个性状的观察、统计和分析发现,有些性状如植株分节数、旗叶长度、旗叶宽度、小穗数、内稃长度在3个物种间表现重叠,差异皆不明显;而其它绝大多数性状在类群间差异明显,至少能把对应比较的两个物种外态间隔大小显示出来,如对于小穗长度、小穗颜色、每小穗小花数、颖体和外稃被毛等性状,长颖固沙草和固沙草相似或接近,而鸡爪草却与其差异明显,说明鸡爪草与前两个物种的外态差距显著;然而,长颖固沙草除颖体长度比固沙草的稍长外,其它特征均与固沙草的相近或相同,说明此两个物种的外态差距极小,是难于界定的两个类群。同时,本研究还对植株高度、旗叶长宽比、叶片和叶鞘被毛状况、圆锥花序长度、小穗颜色、小穗长度、每小穗小花数、颖体长度、颖体被毛状况、外稃长度、外稃被毛状况等12个具有较大分类价值形态性状特征构成的矩阵进行主成分分析。根据特征值大小获得的前3个主成分的变异量分别为43.530%、23.117%和9.435%(累计贡献比率达76.082%),其中前两个主成分代表所研究12个性状中66.647%的信息,为研究类群提供主要的信息,因而依据前两个主成分做二维散点图,结果长颖固沙草和固沙草的所有个体呈现交错分布,未能区分出这两个先前学者鉴定区分的物种;然而,鸡爪草与长颖固沙草和固沙草的所有个体区分明显,表明鸡爪草与其显著不同,应隶属于不同的类群(图1)。
2.2 叶表皮微形态性状差异分析
通过对固沙草属3个物种叶片表皮结构的观察,发现它们皆由长细胞、短细胞、气孔器细胞、刺毛、乳突和微毛组成,并且这些结构成分在叶面的分布、着生情况与他人[3,10~12]报道的虎尾草族(Chlorideae)或画眉草族(Eragrostideae)三齿稃亚族(Tridentinae)的叶表皮特征基本一致,所不同的是3种植物在长细胞形态、气孔器大小及副卫细胞形状、短细胞形态以及刺毛位置上存在各自的差异,完全能够作为区分类群的参考辅助依据(表2,图2)。同时,从表2所列的叶片下表皮微形态性状特征,可以看出长颖固沙草和固沙草理应是两个特征相近的类群,即除了气孔器大小及副卫细胞形状、乳突和微毛这3个性状外,其余绝大多数特征都体现为长颖固沙草与固沙草类同甚至一致,而它们却与鸡爪草呈现出显著差异的特点,实属叶表皮解剖特征明显间断不同的两大类群。
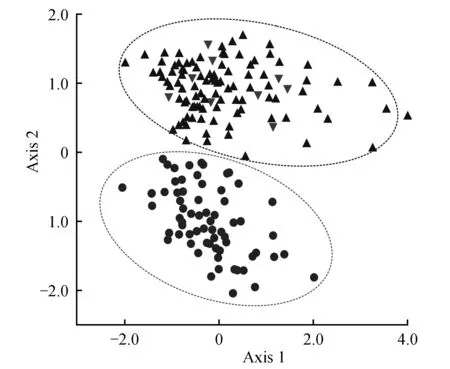
图1 主要外部形态性状主成分的散点图Fig.1 2D PCA scatter plot of principal component in main external morphological characters
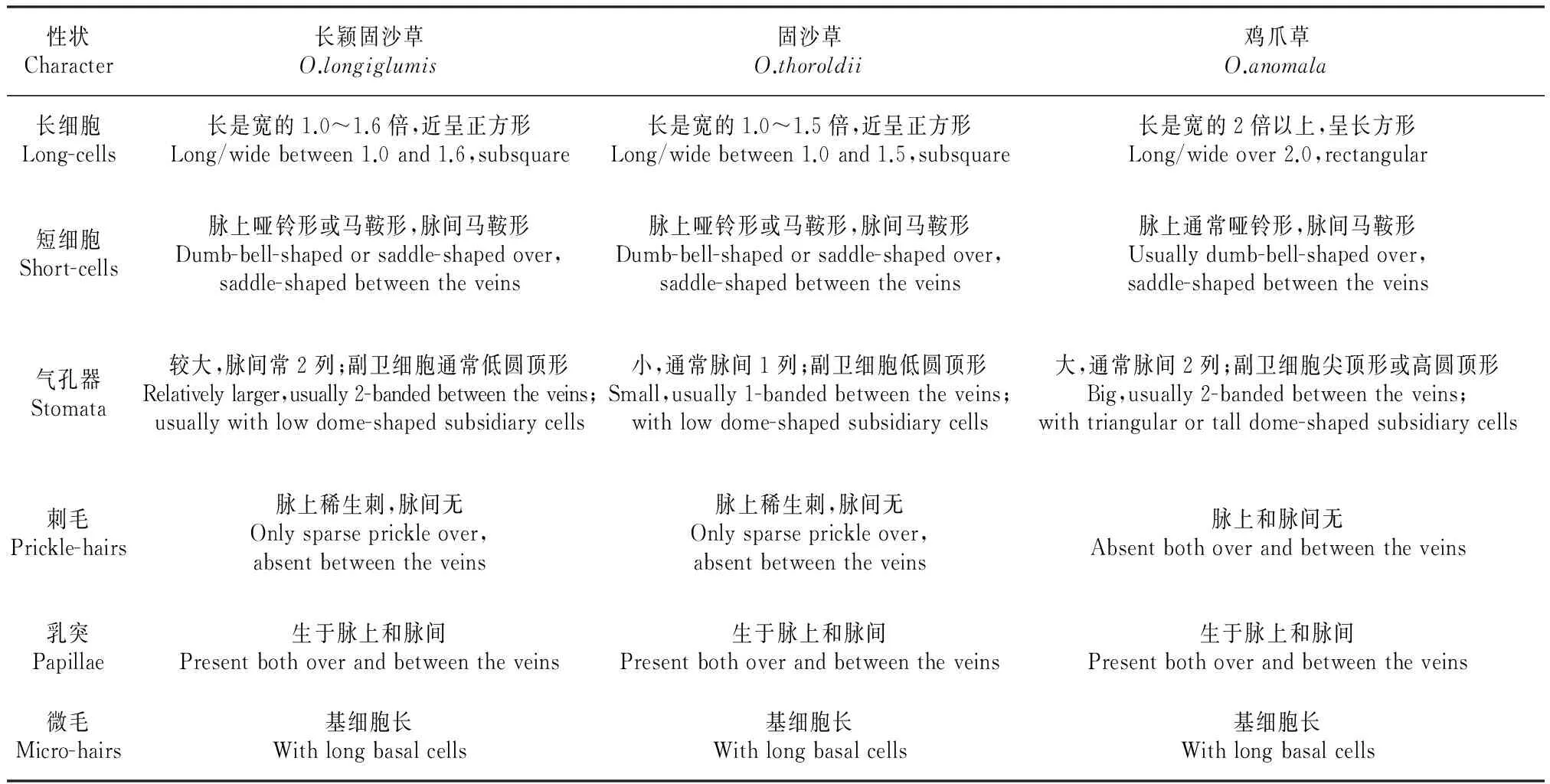
性状Character长颖固沙草O.longiglumis固沙草O.thoroldii鸡爪草O.anomala长细胞Long-cells长是宽的1.0~1.6倍,近呈正方形Long/widebetween1.0and1.6,subsquare长是宽的1.0~1.5倍,近呈正方形Long/widebetween1.0and1.5,subsquare长是宽的2倍以上,呈长方形Long/wideover2.0,rectangular短细胞Short-cells脉上哑铃形或马鞍形,脉间马鞍形Dumb-bell-shapedorsaddle-shapedover,saddle-shapedbetweentheveins脉上哑铃形或马鞍形,脉间马鞍形Dumb-bell-shapedorsaddle-shapedover,saddle-shapedbetweentheveins脉上通常哑铃形,脉间马鞍形Usuallydumb-bell-shapedover,saddle-shapedbetweentheveins气孔器Stomata较大,脉间常2列;副卫细胞通常低圆顶形Relativelylarger,usually2-bandedbetweentheveins;usuallywithlowdome-shapedsubsidiarycells小,通常脉间1列;副卫细胞低圆顶形Small,usually1-bandedbetweentheveins;withlowdome-shapedsubsidiarycells大,通常脉间2列;副卫细胞尖顶形或高圆顶形Big,usually2-bandedbetweentheveins;withtriangularortalldome-shapedsubsidiarycells刺毛Prickle-hairs脉上稀生刺,脉间无Onlysparseprickleover,absentbetweentheveins脉上稀生刺,脉间无Onlysparseprickleover,absentbetweentheveins脉上和脉间无Absentbothoverandbetweentheveins乳突Papillae生于脉上和脉间Presentbothoverandbetweentheveins生于脉上和脉间Presentbothoverandbetweentheveins生于脉上和脉间Presentbothoverandbetweentheveins微毛Micro-hairs基细胞长Withlongbasalcells基细胞长Withlongbasalcells基细胞长Withlongbasalcells

图2 固沙草属3个物种的叶片下表皮光学显微照片 A~C. 长颖固沙草;D~F. 固沙草;G~J. 鸡爪草Fig.2 Light microscopic micrographs of the lower epidermises of leaf blades in three Orinus species A-C. O.longiglumis; D-F. O.thoroldii; G-J. O.anomala

图3 基于ITS序列数据构建的最大简约树和Bayesian 50%多数规则一致性树 分支上数字分别代表最简约树(MP)的自展支持率和Bayesian后验概率。Fig.3 The most parsimonious tree based and fifty percent of Bayesian majority rule consensus tree based on ITS sequences Numbers above the branches indicate bootstrap support and the Bayesian posterior probabilities respectively.
2.3 ITS序列分析
ITS序列比对后全长624 bp;当空位作缺失处理时,全长序列可以提供的总变异位点和信息位点均是33个,占总位点的5.29%,G+C平均含量为56.3%。采用启发式搜索和贝叶斯推论法建立所有个体之间的系统关系树,并给出重复1 000次各分支的自展支持率BV(Bootstrap value)值和贝叶斯系统树(Bayesian inference,BI)的后验概率PP值。比较简约法和贝叶斯法的分析结果,发现两种方法构建的系统发育关系相同,其均能较为清晰地反映物种间的系统发育关系,只是支持率略有不同(图3);所研究的长颖固沙草(O.longiglumis)和固沙草(O.thoroldii)所测个体形成一个独立的分支,另一分支是由外类群鸡爪草(O.anomala)所测个体组成,BV值和PP值分别高达100%和85%,并且长颖固沙草和固沙草两个物种的ITS序列完全相同,这种系统发育树的拓扑结构与依据广义形态性状所获得的研究结果一致。
3 讨论
Grant[13]认为,种内外部形态变异一直困扰着物种界定和分类。我们研究了先前学者确认的3个固沙草属物种,即长颖固沙草、固沙草和鸡爪草的各代表居群的形态变异,发现先前用于界定长颖固沙草、固沙草和鸡爪草的许多外部形态性状如小穗长度、每小穗小花数目、外稃和秆被毛状况、内稃与外稃及颖片与相邻小花的相对长度等即使在同一居群的不同个体间也存在较大变异,这在某种程度上来说,先前学者用于界定长颖固沙草、固沙草和鸡爪草等类群的这些性状不稳定、不全面,它们不足以形成任何阶元等级划分。原始文献[1]报道新种长颖固沙草与近缘类群固沙草的差异是“较长的圆锥花序、颖片和花药,以及内稃短于外稃”,但通过我们进一步的野外考察和形态比较来看,仅鸡爪草与固沙草或长颖固沙草的种间差异明显,而长颖固沙草与固沙草的种间差异却不明显。先前学者在后两个种中指出的花序、颖片、外稃和内稃等性状方面存在差异,而今我们大量调查的结果证实它们之间花序、颖片和花药长度明显重叠,内稃和外稃长度并没有确定的数量关系,可见原始文献在后两物种间重复使用的花序、颖片、外稃、内稃、花药等性状差异不稳定、不全面,确认的两个物种不能成立。同时,PCA分析也表明,长颖固沙草、固沙草的花序、颖片、外稃、内稃及花药长度,小穗颜色、颖片和外稃被毛状况等性状特征在居群间的连续性很难用于类群间界定,说明长颖固沙草和固沙草之间没有明显的种间间断,不足以构成种的界限,很可能就是一个物种的同物异名。
从叶表皮微形态解剖证据来看,长颖固沙草和固沙草均与鸡爪草具有明显的种间间断,这与先前基于外部形态性状划分的结果相一致,表明该两个物种在叶表皮微形态特征上并无种间差异,而是种内的一个变化幅度,从而从侧面支持了本研究对两个物种植物形态比较分析的结果。当然,两种植物在外部形态和叶表皮上也有个别特征还不是完全一致,我们认为这很有可能是由于环境饰变造成的不稳定分异,但其完全不足以形成任何等级阶元划分。再者,就群体遗传学研究结果而言,参试固沙草属物种为一单系类群,并在2种方法中均得到较高支持的BV值和PP值(BV=100%,PP=100%),并且先前确认的长颖固沙草和固沙草居群处于系统发育树的同一个分支上,明显有别于位于另一个分支上的鸡爪草居群,这表明长颖固沙草和固沙草具有极高的相似性,而明显有别于另一个物种—鸡爪草,所以长颖固沙草和固沙草之间是不足以构成任何种间划界的。鉴于此,综合广义形态学和nrITS分子序列信息研究结果,本研究特将长颖固沙草处理为固沙草的异名:
固沙草(禾本科图说)
Orinusthoroldii(Stapf) Bor in Kew Bull. 6:454. 1951[1952];Keng in Fl. III. Pl. Prim. Sin. Gram. 284:229. 1959;Bor in Grass. Burm. Ceyl. Ind. Pak. 519. 1960;Anonymous in Icon. Cormoph. Sin. 5:134. 1976;-Diplachne thoroldii Stapf ex Hemsl. in J. Linn. Soc. Bot. 30:121. 1894. —Cleistogenesthoroldii(Stapf) Roshev. in Fl. Kirgh. 2:107. 1950. —Kengiathoroldii(Stapf) H. Yu & N. X. Zhao in Ann. Bot. Fenn. 42:47. 2005. Type:China. Tibet[Xizang],15,800 ft,Thorold 120(Holotype,K!).—OrinusarenicolaHitchc. in J. Wash. Acad. Sci. 23:136. 1933. Type:[Pakistan],Western Himalayas,Kashmir,Ladak,Tsaka,alt. 4 700 m,1931-07-18,Koelz 2365(Holotype,US-1535771[image!]).
OrinuslongiglumisL. B. Cai & X. Su in Ann. Bot. Fennici 46:143. 2009,syn.nov. TYPE:China. Xizang:Nêdong,Jieba,29°15′N,91°43′E,alt. 3 500 m,2005-07-28,L. B. Cai & C. R. Luosang 002(Holotype & Isotype,HNWP!).
中国特有,产青海、新疆和西藏,生于海拔3 000~4 700 m的山坡、草原、河滩、湖岸及林缘。
1.Su X,Cai L B.Orinuslongiglumis(Poaceae:Chloridoideae),a new species from Xizang(Tibet),China[J].Annales Botanici Fennici,2009,46(2):143-147.
2.苏旭,刘玉萍,李兆孟,等.基于形态和叶表皮微形态特征试论西藏固沙草的归并[J].植物研究,2015,35(3):321-326.
Su X,Liu Y P,Li Z M,et al.Reduction onOrinustibeticusbased on the external morphology and micromorphological characteristics of leaf epidermis[J].Bulletin of Botanical Research,2015,35(3):321-326.
3.Metcalfe C R.Anatomy of the monocotyledons vol.1 Gramineae[M].London:Oxford Clarendon Press,1960:1-713.
4.Doyle J J,Doyle J L.A rapid DNA isolation procedure for small quantities of fresh leaf tissue[J].Phytochemical Bulletin,1987,19:11-15.
5.White T J,Bruns T,Lee S,et al.Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics[M].//Innis M,Gelfand D,Sninsky J,et al.PCR protocols:a guide to methods and applications.San Diego,CA:Academic Press,1990:315-322.
6.Thompson J D,Gibson T J,Plewniak F,et al.The CLUSTAL_X windows interface:flexible strategies for multiple sequence alignment aided by quality analysis tools[J].Nucleic Acids Research,1997,25(24):4876-4882.
7.Tamura K,Peterson D,Peterson N,et al.MEGA5:molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance,and maximum parsimony methods[J].Molecular Biology and Evolution,2011,28(10):2731-2739.
8.Swofford D L.PAUP:phylogenetic analysis using parsimony(and other methods),version 4.0[M].Sunderland,MA:Sinauer Associates,2002.
9.Ronquist F,Huelsenbeck J P.MrBayes 3:Bayesian phylogenetic inference under mixed models[J].Bioinformatics,2003,19(12):1572-1574.
10.蔡联炳,郭延平.禾本科植物叶片表皮结构细胞主要类型的演化与系统分类和发育途径的探讨[J].西北植物学报,1995,15(4):323-335.
Cai L B,Guo Y P.Studies on constituent cells of leaf epidermis,systematics and phylogenetic path of the family Poaceae[J].Acta Botanica Boreali-Occidentalia Sinica,1995,15(4):323-335.
11.蔡联炳,郭延平.禾本科植物叶片表皮结构细胞主要类型的演化与系统分类和发育途径的探讨(续)[J].西北植物学报,1996,16(1):65-72.
Cai L B,Guo Y P.Studies on constituent cells of leaf epidermis,systematics and phylogenetic path of the family Poaceae(Continued)[J].Acta Botanica Boreali-Occidentalia Sinica,1996,16(1):65-72.
12.蔡联炳,张燕,张同林.根据叶表皮特征试论国产三齿稃亚族的属间关系[J].西北植物学报,2004,24(9):1691-1696.
Cai L B,Zhang Y,Zhang T L.Relationships among genera ofTridentinaefrom China based on some leaf epidermal characteristics[J].Acta Botanica Boreali-Occidentalia Sinica,2004,24(9):1691-1696.
13.Grant V.Plant speciation[M].2nd ed.New York:Columbia University Press,1981.
Under the auspices of the National Natural Science Foundation of China(No.31260052);The National Natural Science Foundation of Qinghai Province(2016-ZJ-937Q,2014-ZJ-947Q);The Key Laboratory of Medicinal Plant and Animal Resourcesofthe Qinghai-Tibetan Plateau(2015-Z-Y06);The Young and Middle-Aged Research Foundation of Qinghai Normal University(2015-19)
introduction:LIU Yu-Ping(1980—),female,doctor,associate professor,major in plant systematics and evolution.
date:2016-11-07
ReductiononOrinuslongiglumiswithGeneralizedMorphologicalCharactersandnrDNAITSSequence
LIU Yu-Ping LÜ Ting ZHOU Yong-Hui ZHANG Xiao-Yu SU Xu*
(Key Laboratory of Medicinal Plant and Animal Resources in the Qinghai-Tibetan Plateau,Key Laboratory of Physical Geography and Environmental Process in Qinghai Province,Key Laboratory of Education Ministry of Environments and Resources in the Qinghai-Tibetan Plateau,School of Geography and Life Science,Qinghai Normal University,Xi’ning 810008)
Orinusis an alpine genus with important economic values in the grass family(Poaceae). There are six species in it and all of them have stronger adverse-resistant characteristics, and they are the important resources for improving and applying agricultural breeding and animal husbandry. Meanwhile, the discriminative characters are obscure among these species. We compared and analyzed the main morphological characters and molecular sequence differences among nearest relatedOrinuslongiglumis,O.thoroldiiandO.anomalaby generalized morphological observation and nuclear ITS region research. There is very slight disparity betweenO.longiglumisandO.thoroldiiin generalized morphological characters, the classification of the two species is very difficult, the differences betweenO.longiglumisandO.anomalaare distinct, and the two taxa are two different species. We further used the ITS sequence variation to tentatively delimit the previously acknowledged species and found that there is no distinct boundary between species and only two distinct lineages were recognized for all three species. A branch is composed ofO.longiglumisandO.thoroldii, while another is only made up ofO.anomala. The bootstrap support rates of two branches reach as high as 100%. Therefore,O.longiglumisandO.thoroldiishould belong to the same species, andO.longiglumiscan be reduced to synonymy ofO.thoroldii.
Poaceae;Orinus;Orinuslongiglumis;morphological character;ITS sequence
国家自然科学基金项目(31260052);青海省自然科学基金项目(2016-ZJ-937Q,2014-ZJ-947Q);青藏高原药用动植物资源重点实验室项目(2015-Z-Y06);青海师范大学中青年科研基金项目(2015-19)
刘玉萍(1980—),女,博士,副教授,主要从事系统与进化植物学研究。
* 通信作者:E-mail:xusu8527972@126.com
2016-11-07
* Corresponding author:E-mail:xusu8527972@126.com
Q949.71
A
10.7525/j.issn.1673-5102.2017.02.003
