家蚕红色卵致死突变Fuyin-lre的性状特点与遗传分析
2017-10-23廖鹏飞丁善明陈安利李琼艳罗顺高朱水芬董占鹏
廖鹏飞,丁善明,陈安利,李琼艳,罗顺高,朱水芬,黄 平,董占鹏
(云南省农业科学院蚕桑蜜蜂研究所,云南 蒙自 661101)
家蚕红色卵致死突变Fuyin-lre的性状特点与遗传分析
廖鹏飞,丁善明*,陈安利,李琼艳,罗顺高,朱水芬,黄 平,董占鹏**
(云南省农业科学院蚕桑蜜蜂研究所,云南 蒙自 661101)
【目的】为了解发现于云南家蚕种质资源“芙印”系统红色卵致死突变Fuyin-lre的性状特点和遗传性。【方法】观察Fuyin-lre的卵形、卵色、胚胎发育及孵化情况,并与正常卵系统P50、突变红色卵标志系统re开展杂交、回交等遗传分析试验。【结果】Fuyin-lre系统出现全紫褐色卵、紫褐色卵与红色卵并存的2种蛾区,紫褐色卵能正常孵化发育,而红色卵稍透明,常会出现不规则的条状或块状的黑斑,胚胎发育至丙1时,停止发育不孵化,与已研究报道的红色卵突变re、ci、rec、b-4、ru等存在明显的差异而与rel相似;Fuyin-lre系统与P50杂交F1代均为紫褐色卵,F2代与BC1代出现紫褐色卵∶红色卵为3∶1的突变蛾区,所有紫褐色卵能正常孵化发育而红色卵不孵化;Fuyin-lre系统与re的F1、F2和BC1均会出现红色卵突变蛾区,依交配方式的不同,突变蛾区中有全红色卵、1/2红色卵和1/4红色卵等多种表现形式,其中红色卵存在正常孵化、孵化1/2或孵化3/4等差异。【结论】Fuyin-lre突变发生于re基因所在的第5染色体上,受隐性基因控制,纯合致死,遵循孟德尔遗传规律。
家蚕;红色卵突变;致死;性状特点;遗传分析
家蚕是重要的农业经济昆虫,经5000余年的驯化利用,中国与日本收集保存有基因突变, 染色体变异及特色遗传材料1000余份[1-2]。家蚕突变资源是创建特色育种素材[3-6]和选育特殊用途家蚕品种[7-11]的种质基础,又是鳞翅目模式昆虫和遗传学研究的重要材料[12-16]。【研究意义】通过研究家蚕红色卵致死突变Fuyin-lre的性状特点和遗传性,丰富红色卵理论知识,为红色卵致死突变的有效利用奠定基础。【前人研究进展】家蚕的卵色是卵壳、浆液膜和卵黄等颜色的复合性状,在家蚕的遗传学研究中占据重要地位[17-18]。卵色突变主要为卵壳色突变和浆液膜色突变,其中卵壳形成自母体,故卵壳色突变与卵形突变相似,均遵循家蚕卵壳性状的伪母性遗传规律[15-18]。而浆液膜色主要由浆液膜细胞所含色素决定,通常浆液膜色与成虫复眼色是一致的。正常型浆液膜含有黑色素,卵呈紫褐色或灰绿色,而突变型浆液膜或不含色素,或含红色素、褐色素等致蚕卵表现为白色、红色、褐色等性状,其遗传方式有孟德尔遗传、母性遗传等多种[18]。目前报道的红色卵突变有红色卵(re)、红褐卵(rec)、橙色卵(ci)、第4褐卵(b-4)、锈色卵(ru)等,能正常孵化发育而不具有胚胎致死性[18-21]。【本研究的切入点】2009年笔者在保存的家蚕种质资源“芙印”系统中发现一种红色卵致死突变,突变卵呈红色,且有胚胎致死性,命名为Fuyin-lre,简记为lre。Fuyin-lre与上述的re、rec、ci、b-4、ru的性状存在较大差异,而与日本家蚕种质资源库中记载的rel的性状表现相似,但rel性状描述较少,且未见其遗传分析的相关报道。【拟解决的关键问题】研究分析Fuyin-lre突变的性状特点,并弄清该突变性状的遗传性。
1 材料与方法
1.1 材料
红色卵致死突变系统:Fuyin-lre,由云南省农业科学院蚕桑蜜蜂研究所从家蚕种质资源“芙印”系统中发现并保存。该系统出现全紫褐色卵蛾区(简称正常卵蛾区)、紫褐色卵和红色卵并存的蛾区(简称突变蛾区)两种表现型式。紫褐色卵能正常孵化发育,而红色卵具有胚胎致死性,不能孵化。
正常卵色系统:P50(大造),卵色为灰紫色,引自苏州大学。
红色卵突变系统:re,引自西南大学家蚕基因库,该突变系统蚕卵全为红色,能正常孵化发育,不具有胚胎致死性。
1.2 方法
1.2.1 蚕卵性状调查 从Fuyin-lre突变系统的产卵开始观察卵形、卵色随时间的变化情况以及各卵色蚕卵经常规处理催青后的孵化表现,并用荧光体视显微镜 Leica M165 FC对正常卵和红色卵进行显微观察,比较正常卵与红色卵的性状差异。
1.2.2 胚胎发育调查 将Fuyin-lre突变系统的紫褐色卵(简称“正常卵”)与红色卵均按常规方法进行蚕种处理和催青,待正常卵孵化后,解剖红色卵,在解剖镜下观察胚胎终止发育时的外观形态。
1.2.3 遗传分析 将Fuyin-lre突变系统中发生红色卵突变蛾区的正常个体进行自交,并分别与正常卵系统P50、红色卵突变系统re进行杂交和回交,调查各交配后代F1、F2和BC1蛾区内和蛾区间的卵色分离情况,及突变蛾区内正常卵和红色卵的孵化情况,分析Fuyin-lre突变的遗传特点。
2 结果与分析
2.1 Fuyin-lre突变的蚕卵性状表现
Fuyin-lre突变系统的蚕卵在刚产出时,卵形、卵色无明显差异,均为短椭圆形、淡黄色卵。卵龄约30 h时开始出现色差,部分蚕卵颜色无明显变化,部分卵色加深成浅赤豆色;产卵后约48 h,前期卵色无明显变化的蚕卵颜色也开始加深成深黄色,而浅赤豆色蚕卵已变为赤豆色;72 h卵龄时,深黄色蚕卵渐成浅赤豆色,而赤豆色蚕卵颜色进一步加深成淡茶褐色;产卵72 h后,蚕卵颜色变化较慢,随着时间的推移,浅赤豆色蚕卵逐渐转为红色,淡茶褐色蚕卵逐渐转为紫褐色。观察发现,Fuyin-lre突变系统分化出全紫褐色的正常卵蛾区(图1A,封三)和紫褐色卵与红色卵并存的突变卵蛾区(图1B,封三),紫褐色卵能够正常孵化(图1C、1D,封三),而红色卵全部为死卵(图1D,封三)。
经荧光体视显微镜观察发现,紫褐色卵不透明,而红色卵呈现半透明状,透明度随红色卵颜色的加深而降低,在卵粒的不同部位常会出现不规则的条状或块状的黑色斑块(图2,封三)。
2.2 Fuyin-lre突变的胚胎发育
解剖Fuyin-lre突变蚕卵显示,红色卵胚胎与卵壳粘黏较为严重,解剖的难度较大,不易得到完整胚胎,且黏附物较多,可见大致的轮廓(图3,封三)。红色卵胚胎体躯较长,头尾褶增大,能隐约看见头褶凹陷和腹部、尾部环节,第1~2环节正中未见纵沟,形态与丙1期胚胎相似,推测红色卵胚胎可能发育至丙1期即停止发育。
2.3 Fuyin-lre突变的遗传分析
2.3.1 Fuyin-lre突变的遗传分离 从Fuyin-lre突变系统内选择突变蛾区的正常卵进行单蛾饲育,同蛾区自交,每次自交后代均出现正常卵蛾区和突变卵蛾区2种表现型式,经多代自交后,随机选择4个发生红色卵突变的蛾区调查正常卵粒数与红色卵粒数,比例符合3∶1的分离规律,其中正常卵的孵化率在90 %以上,而红色卵的孵化率为0(表1)。调查4个蛾区的自交后代正常卵蛾区与突变卵蛾区的分离比均符合5∶4的分离规律(表2)。以上结果表明Fuyin-lre突变系统的正常卵存在显性纯合和杂合2种基因型,红色卵致死性状由隐性遗传基因控制,遵循孟德尔遗传规律。
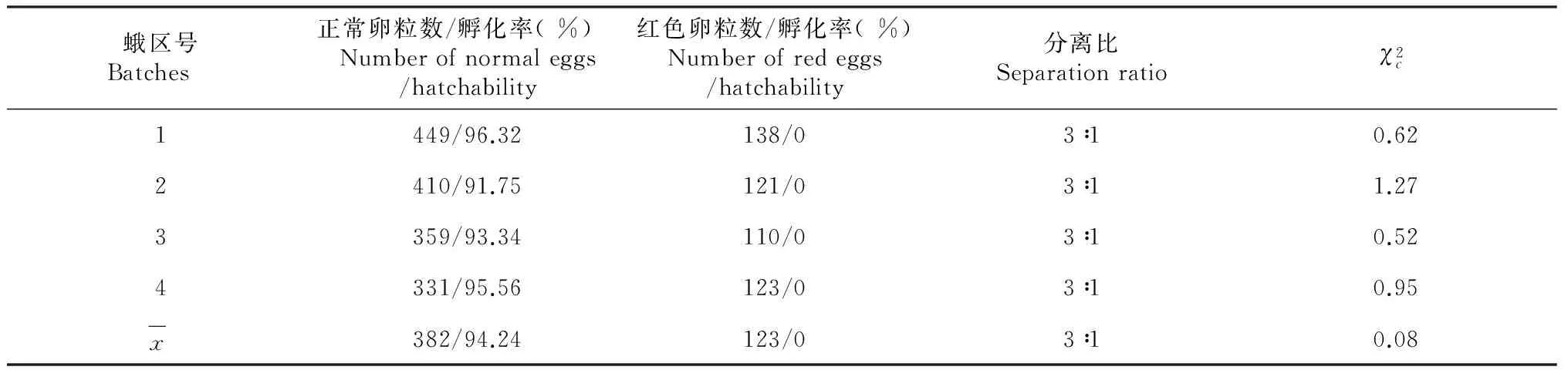
表1 Fuyin-lre突变蛾区内正常卵与红色卵的分离和孵化Table 1 The separation and hatchability of normal eggs and red eggs in batches of the mutant Fuyin-lre
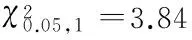
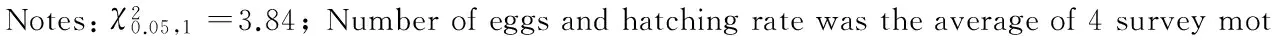

表2 Fuyin-lre突变自交后代蛾区卵色分离Table 2 The egg color separation of the mutant Fuyin-lre in batch after selfing
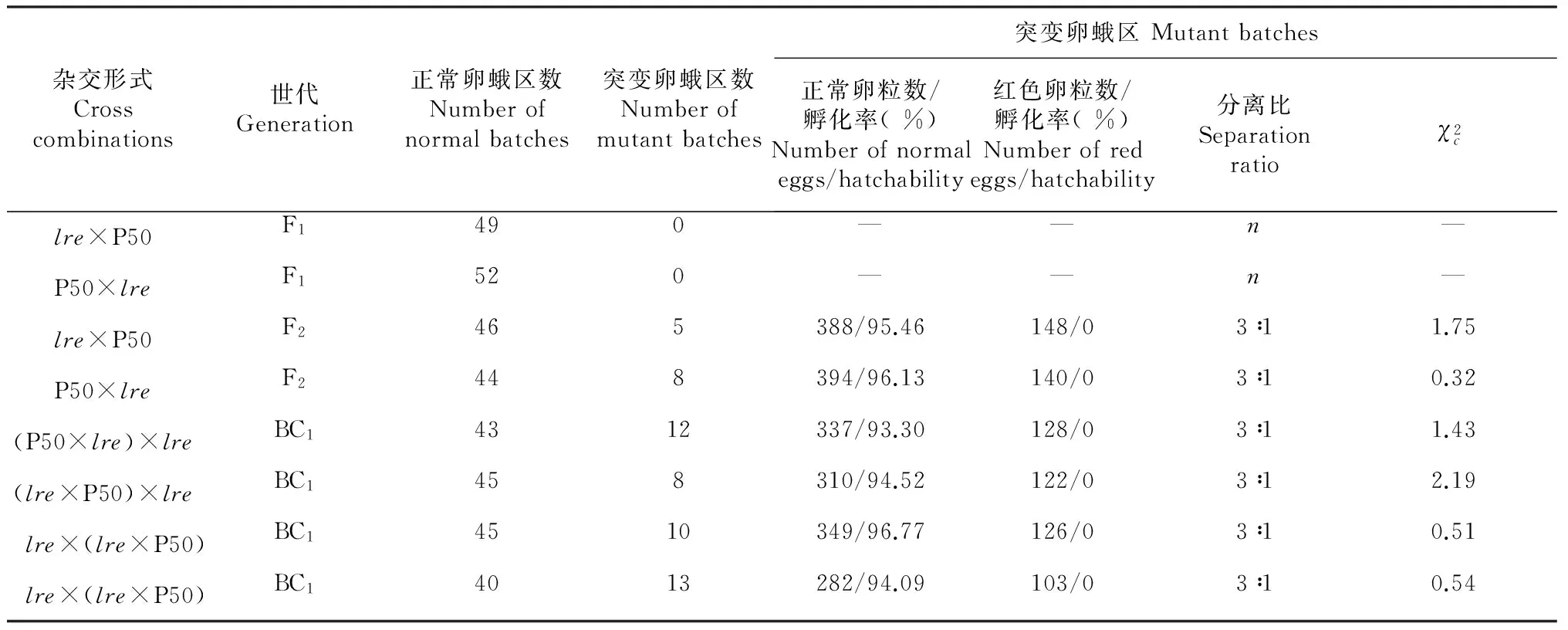
表3 Fuyin-lre突变与P50正反交后代正常卵与红色卵的分离Table 3 The separation of normal eggs and red eggs of the reciprocal cross progenies of Fuyin-lre and P50
注: “—”表示无数据可调查统计;“n”表示无性状分离,下同。
Notes: ‘—’represented no data available statistics;‘N’ represented no character separation.The same as below.
2.3.2 Fuyin-lre突变与正常卵P50的杂交试验 为了进一步验证Fuyin-lre突变的遗传特性,将突变系中发生红色卵突变蛾区的正常个体(因突变个体致死不孵化,不能用于交配试验)与正常卵系P50杂交,结果显示其正反交F1代均为紫褐色卵且能正常孵化发育,这与红色卵致死突变受隐性基因控制的结论相符。为减少饲育蛾区数,正反交F1混合育,正反交的F2代中均发生正常卵蛾区和突变卵蛾区两种型式,调查突变卵蛾区发现,正常卵与红色卵的分离比符合3∶1的分离规律,且只有正常卵能够孵化发育,而红色卵不孵化,表现出红色卵的强致死作用(表3)。结果表明,红色卵致死突变由隐性基因控制,在隐性纯合时表现为红色卵致死。
正反交F1代与Fuyin-lre突变系统中突变蛾区的正常个体进行回交,调查BC1代红色卵的分离情况。结果显示,BC1代均发生正常卵蛾区和突变卵蛾区的分离,在突变卵蛾区中,正常卵与红色卵的分离比为3∶1,正常卵能正常孵化发育,而红色卵致死不孵化(表3)。结果进一步证明红色卵致死性状由隐性基因控制,在隐性纯合时表现为红色卵致死。
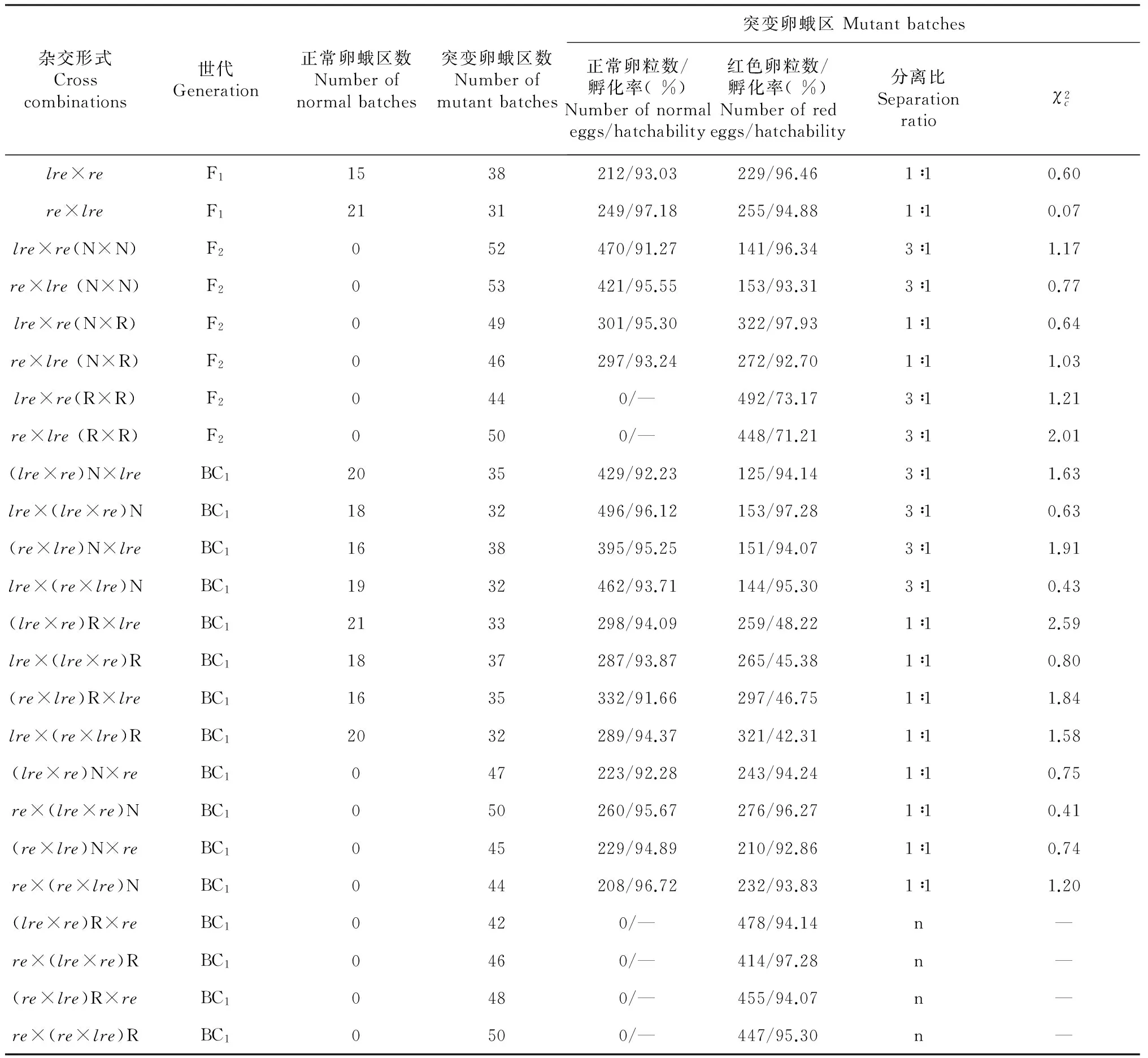
表4 Fuyin-lre与re正反交后代正常卵与红色卵的分离Table 4 The separation of normal eggs and red eggs of the reciprocal cross progenies of Fuyin-lre and re
2.3.3 Fuyin-lre突变与红色卵突变re的杂交试验 家蚕红色卵突变re受隐性遗传基因控制,隐性纯合时即产生红色卵,红色卵能够正常孵化发育,不具有胚胎致死性。为了研究红色卵突变re与Fuyin-lre突变间的遗传关系,笔者进行了Fuyin-lre与re的杂交试验(表4)。结果显示,正反交的F1代均有正常卵蛾区和突变卵蛾区2种表现型,其中突变卵蛾区正常卵与红色卵的比例均为1∶1,2种卵色的蚕卵均能够正常孵化。
将F1代出现红色卵的蛾区,依卵色分别收蚁饲养(即将红色卵和正常卵剥离单独收蚁饲养),按照正常卵×正常卵(N×N),正常卵×红色卵(N×R),红色卵×红色卵(R×R)的交配方式制种,结果显示N×N的所有蛾区中均有红色卵出现,正常卵与红色卵的比例为3∶1;N×R的所有蛾区中也均有红色卵出现,正常卵与红色卵的比例为1∶1;R×R的所有蛾区均为纯红色卵,没有紫褐色的正常卵出现。对各种表型的蛾区进行浸酸催青,发现正常卵与红色卵比例为3∶1和1∶1的蛾区的所有蚕卵均能够正常孵化,而全红色卵圈中的红色卵孵化率为71 %~74 %,经适合性测试发现全红色卵圈中孵化的卵粒数与未孵化的卵粒数符合3∶1的分离规律。
将F1代发生红色卵的蛾区依卵色(正常卵和红色卵)单独收蚁饲养至发蛾后,分别与Fuyin-lre突变中突变蛾区的正常个体正反回交(BC1)。结果发现F1代与Fuyin-lre中正常个体回交得到的BC1代均出现正常蛾区与突变蛾区2种表现型,其中F1代正常卵[(lre×re)N 或(re×lre)N]回交Fuyin-lre突变蛾区中正常卵得到的BC1代突变蛾区的正常卵与红色卵符合3∶1的分离规律,且正常卵和红色卵均能正常孵化,而F1代红色卵[(lre×re)R 或(re×lre)R]回交Fuyin-lre突变蛾区中正常卵得到的BC1代突变蛾区的正常卵与红色卵的分离比则为1∶1,正常卵能正常孵化,而红色卵的孵化率则介于40 %~50 %,孵化卵粒数与未孵化卵粒数的比例符合1∶1的分离规律。
将F1代发生红色卵的蛾区依卵色(正常卵和红色卵)单独收蚁饲养至发蛾后,分别与re突变进行正反回交(BC1)。结果显示BC1代的所有蛾区均有红色卵出现,其中F1代正常卵[(lre×re)N 或(re×lre)N]回交re突变系得到的BC1代的正常卵与红色卵的分离比为1∶1,正常卵与红色卵均正常孵化;F1代红色卵[(lre×re)R 或(re×lre)R]回交re红色卵突变系得到的BC1代均为红色卵,且能正常孵化。
以上试验结果表明,Fuyin-lre突变系统控制红色卵性状的基因位点与re相同,控制胚胎致死性状的基因位于re所在的第5染色体上,且为隐性遗传。
3 讨 论
卵壳色形成自母体,Fuyin-lre红色卵虽然不孵化未能观察到其卵壳色,但可能与紫褐色卵壳一致为乳白色,因此Fuyin-lre红色卵的卵色可能与re一样,均因浆液膜为红色所致;经遗传分析Fuyin-lre的红色卵性状和致死性状均受隐性基因控制,其中控制红色卵性状的基因位点与红色卵突变re相同,而控制致死性状的基因位于红色卵突变re所在的第5染色体上。本文的杂交、回交试验结果尚不能推断出控制致死性状基因在第5染色体上的确切位置。因此,Fuyin-lre突变是否像re和rel一样是同一位点的突变,或者实质上是rel突变,或者存在红色卵和致死2种遗传距离较短的连锁突变还有待于通过两点测验、三点测验或分子生物学方法确定。
因Fuyin-lre突变属纯合致死,故仅能以杂合体形式保存。为实现Fuyin-lre的有效保存,降低突变基因丢失的风险,通常需要选取紫褐色卵和红色卵并存的蛾区(显性纯合占比1/3,杂合占比2/3)进行继代保存,出现红色卵突变体后代( 蛾区)的概率为4/9,因此一般情况下需要扩大繁育制种量到56蛾以上,使突变蛾区尽可能达到家蚕种质资源继代保存所需的数量要求。
家蚕是鳞翅目昆虫的代表,同时也是突变基因研究成果最为丰富的生物之一,通过对家蚕突变基因的研究,可以为进一步研究其他鳞翅目昆虫提供遗传基因材料。家蚕红色卵致死突变Fuyin-lre的发现,如同溃卵突变 (Grcol)[22]、“明”死卵突变(1-em)[15]、家蚕新小卵突变(sm-n)[14]等,可以丰富中国家蚕突变基因资源,同时也为研究家蚕卵致死突变的规律提供良好的材料。本研究通过对红色卵致死突变性状调查和遗传分析,为进一步研究其致死的发生机理奠定了遗传学基础,在深入研究基因的信息及其致死分子机制、明确红色卵形成和胚胎致死之间的本质关系、揭示红色卵致死这一独特现象产生的原因后,红色卵致死突变有望成为开发田间害虫种群遗传控制技术的靶标基因素材。
4 结 论
Fuyin-lre系统存在全紫褐色卵蛾区、紫褐色卵和红色卵并存的蛾区两种表现型式,其中存在红色卵的蛾区占比为4/9,紫褐色卵和红色卵并存蛾区中红色卵的占比为1/3;红色卵卵形与正常的紫褐色卵卵形一致,均为短椭圆形;刚产出的蚕卵,均为淡黄色,但随着时间的推移,色差逐渐显现、扩大而形成红色与紫褐色的明显差异;紫褐色卵能正常孵化发育,而红色卵稍透明,常伴随不规则的条状或块状的黑斑,发育至丙1期时停止发育不孵化;Fuyin-lre突变发生于re基因所在的第5染色体上,受隐性基因控制,纯合致死,遵循孟德尔遗传规律。
[1]代方银.家蚕突变基因的遗传与近等位基因系研究[D].重庆:西南大学博士学位论文,2008:1-155.
[2]鲁 成,代方银,向仲怀.家蚕基因库突变系统的研究[J].中国农业科学,2003,36(8):968-975.
[3]张剑飞,杜周和,刘彬斌,等.家蚕斑纹茧色特色育种素材创制及利用[J].西南农业学报,2016,29(6):1481-1487.
[4]秦 俭,易文仲.家蚕突变基因分析和基因资源的建立及应用研究[J ].蚕业科学,1996,22(1):13-19.
[4]司马杨虎,孙德斌.家蚕特色种质资源的创新开发及其应用前景[J].广西蚕业,2000,37(3):17-21.
[5]廖鹏飞,陈安利,朱水芬,等.一种将性连锁平衡致死系限性卵色和致死基因导入现行家蚕品种的方法[J].蚕业科学,2014,40(6): 1011-1016.
[6]颜海燕,杨明观,钟伯雄.利用温敏致死性状专养雄蚕的研究进展[J].蚕桑通报,2002,33(1):4-7.
[7]肖金树,张友洪,周安莲,等.限性黄茧品种“蜀·黄×川·白”的育成[J].蚕业科学,2008,34(4):745-751.
[8]刘 敏,吴克军,田梅惠,等.春用家蚕皮斑双限性品种”云蚕10号的选育” [J].西南农业学报,2013,26(5):2147-2152.
[9]大规良树.日本育成不同新用途的蚕品种[J].蚕丝科学与技术 (日),1991,30( 2):30-34.
[10]刘俊凤,杜周和,张剑飞,等.家蚕限性种质资源主要经济性状的稳定性测定及其在育种亲本选择中的应用[J].西南农业学报,2008,21(3):842-848.
[11]王 磊,黄德辉,孙家羿,等.基因突变在家蚕育种应用上的研究进展[J].安徽农业科学,2007,35(34):11123-11125.
[12]秦 俭,何宁佳,向仲怀.家蚕模式化研究进展[J].蚕业科学,2010,36(4): 645-649.
[13]张凤林,娄齐年,聂 磊,等.家蚕突变型灰黑蛾杂交性状调查与遗传分析[J].山东农业科学,2009(6):99-101.
[14]代方银,王先燕,胡 海,等.家蚕突变新小卵(sm-n)的特异性状及其遗传[J].蚕业科学,2006,32(4):459-463.
[15]陈安利,赵巧玲,张国政,等.家蚕卵突变体“明”死卵(1-em)的卵壳构造和遗传分析[J].蚕业科学,2009,35(1): 139-143.
[16]范晓东,代方银,童晓玲,等.家蚕保存系统红色卵的遗传分析[J].昆虫学报,2006,49(4): 543-549.
[17]吕鸿声.蚕种学原理[M].上海:上海科学技术出版社,2011:179-184.
[18]向仲怀.家蚕遗传育种学[M].北京:中国农业出版社,1994:273-289.
[19]鲁 成,代方银,向仲怀.家蚕橙色卵(ci)基因的连锁检索与定位研究[J].蚕业科学,1999,25(2):74-76.
[20]鲁 成,王枫,向仲怀,等.家蚕锈色卵的遗传学研究[J].蚕学通讯,1988,7(4):1-8.
[21]向仲怀.蚕丝生物学[M].北京:中国林业出版社,2005:107-132.
[22]Nadel M R,Goldsmith M R,Goplemd J,et a1.A secretory defect of chorion formation in the Grcolmutant ofBombyxmori[J].Dev.Biol.,1980,75(1):41-58.
CharacterFeaturesandGeneticAnalysisofLethalRedEggMutantFuyin-lreofSilkworm,Bombyxmori
LIAO Peng-fei,DING Shan-ming*,CHEN An-li,LI Qiong-yan,LUO Shun-gao,ZHU Shui-fen,HUANG Ping, DONG Zhan-peng**
(The Sericultural & Apicultural Research Institute,Yunnan Academy of Agricultural Sciences,Yunnan Mengzi 661101,China)
【Objective】The present study was conducted to investigate the character features and hereditary of red egg mutant Fuyin-lrewhich was found in silkworm (Bombyxmori) germplasm resource Fuyin.【Method】We observed the egg shape,egg colour,embryo development and incubation of Fuyin-lre,and hybridizing with normal egg system P50,red egg mutant system to analyze the hereditary law.【Result】There were two kind batchs of Fuyin-lresystem,all of the eggs were purple brown eggs in one batch,purple brown eggs and red eggs coexisted in the other batch.The purple brown eggs could develop normally,but the red eggs slightly transparence,often appeared irregular strips or blocks of dark spots,and the embryos of the red eggs stopped develop atbing-1 embryo.It was obviously different from the reported red egg mutant systemsre,ci,rec,b-4,ru,but similar torel.Hybrid offspring (F1) of Fuyin-lreand P50 were purple brown eggs.The batchs which included purple brown eggs and red eggs (purple brown eggs∶red eggs =3∶1) appeared in F2and BC1.All the brown eggs hatched normally,but the red eggs could not hatch.The batchs included red eggs appeared in hybrid offspring F1,F2and BC1of Fuyin-lreandre.The red eggs in mutant batchs had various types,including all red eggs,1/2 red eggs and 1/4 red eggs,and so on.Hatching rate of red eggs included normal hatching,hatching 1/2 or hatching 3/4 types.【Conclusion】 The Fuyin-lremutant occurred in the fifth linkage group,followed recessive inheritance,and conformed to the mendelian inheritance.
Bombyxmori; Red egg mutant; Lethal;Character features;Genetic analysis
1001-4829(2017)4-0969-06
10.16213/j.cnki.scjas.2017.4.041
2016-07-05
国家自然科学基金(31460640);云南省现代农业蚕桑产业技术体系建设项目(2015KJTX006);云南省重点新产品开发计划项目(2013BB007、2015BB008);国家现代农业产业技术体系专项(CARS-22)
廖鹏飞(1973-),男,四川中江人,副研究员,主要从事家蚕种质资源保存与创新利用,E-mail:liaolpf@126.com,*为共同第一作者:丁善明,E-mail:dingsm_yn@sohu.com,**为通讯作者,董占鹏,E-mail:scsdong@163.com。
S851.34+7.31
A
(责任编辑 王家银)
