沂蒙山区与文昌中华蜜蜂形态鉴定及其分类研究
2017-10-21雪姜风涛娄德龙高景林李延璨马兰婷周延忠王桂芝
龚 雪姜风涛娄德龙高景林李延璨马兰婷周延忠王桂芝
(1山东农业大学动科院,泰安271018;2山东省蜂业良种繁育推广中心,泰安271000;3中国热带农业科学院环境与植物保护研究所,海口571101;4山东省沂源县畜牧局,沂源256100)
沂蒙山区与文昌中华蜜蜂形态鉴定及其分类研究
龚 雪1姜风涛2娄德龙2高景林3李延璨1马兰婷1周延忠4王桂芝1
(1山东农业大学动科院,泰安271018;2山东省蜂业良种繁育推广中心,泰安271000;3中国热带农业科学院环境与植物保护研究所,海口571101;4山东省沂源县畜牧局,沂源256100)
本研究对沂蒙山区(蒙阴、沂源)的中华蜜蜂资源做了调查取样,按照国际指标对蒙阴、沂源中华蜜蜂样本进行了42个形态特征的系统测量和分析,利用SPSS 22软件进行了主成分、判别、聚类、变异及与环境因子的相关等数据分析,并与海南文昌中华蜜蜂进行形态及分类比较。结果为:15群中华蜜蜂进行聚类且聚为四类,第一类包括 YY7、MY3、YY3、MY4、YY4、YY5,第二类有 YY1、YY8、YY2、YY6,第三类为 MY1、MY2,第四类为 HN1、HN2、HN3。判别分析支持上述结果;PCA得到的主成分1和主成分2在各地区间差异极显著。主成分1与纬度和海拔呈极显著正相关,与年气温呈极显著负相关,所测中华蜜蜂的体形随海拔升高和纬度的北移以及年均气温的降低而增大;主成分2与经度和年降雨量以及年气温呈极显著正相关,与海拔高度极显著负相关。
中华蜜蜂;形态特征;分类;沂蒙山区
东方蜜蜂(Apis cerana Febricius)分布于亚洲各地,中华蜜蜂是东方蜜蜂的指名亚种,杨冠煌等认为中华蜜蜂主要分为4个亚亚种,即东部中蜂、海南中蜂、阿坝中蜂和西藏中蜂[1],后期研究者将滇南中蜂列为我国第五个中华蜜蜂亚亚种。东部中蜂广泛分布在我国大陆东部丘陵和山区,由于生态条件的差异,东部中蜂又分为5个生态型:闽粤型、两湖型、云贵高原型、北方型和长白山型。北方型分布在北京、河北、河南、山东、山西、陕西的丘陵山区。按照上述分类(型)方法,山东中华蜜蜂属于北方型[2-5]。
近年来,山东省政府及蜂业主管部门对境内中华蜜蜂实施了一系列保护措施,建立了6个省、县市级保护区,中华蜜蜂的养殖数量正在稳步提高。2017年春,沂蒙山区申报国家级中华蜜蜂保护区。
本研究旨在对沂蒙山区的中华蜜蜂资源进行全面调查并进行形态特征的全面鉴定,同时与海南文昌中华蜜蜂进行形态特征比较,为开发和保护中华蜜蜂资源奠定理论基础。
1 材料与方法
1.1 研究材料及形态分析方法
2016年8月于山东省沂蒙山区(沂源、蒙阴)和海南文昌采集中华蜜蜂样本15群,其中临沂蒙阴4群,淄博市沂源8群,海南文昌3群。所采集地区的样本主要从活框蜂箱内、自然巢房(如树洞蜂巢)及半人工饲养的蜂巢内(如树桶蜂巢、墙洞蜂巢)进行蜂样采集。
形态测定方法按照国际通用的42项形态特征进行鉴定分析[7-12]来测定。所得数据除计算平均值和变异系数外,运用SPSS 22.0进行主成分、聚类及关联等分析。
2 结果与分析
2.1 三地区中华蜜蜂方差分析

表1 三个地区中华蜜蜂形态特征
表1为三个地区的中华蜜蜂样本形态特征方差(ANOVA)分析,可以看出:在色型上,除了唇基、上唇以及小盾片的K/B两区在三个地区中差异不显著,小盾片Sc区的色度文昌中华蜜蜂与沂源和蒙阴中华蜜蜂均有显著差异(P<0.05),第2~4背板色型中,沂蒙山区中华蜜蜂间差异不显著(P>0.05),但与文昌中华蜜蜂有显著差异(P<0.05);在体尺数据上,第 3~4背板长、蜡镜长、基附节长等10项指标差异在三个地区间差异不显著(P>0.05),第四腹板长、蜡镜间距、前翅长和宽等12项指标都存在差异,沂源中华蜜蜂在吻长、第7腹板长等5个指标上表现出最长,与沂源和文昌中华蜜蜂差异显著(P<0.05);蒙阴中华蜜蜂前翅长与宽2项指标上表现最大,与沂源和文昌中华蜜蜂差异显著(P<0.05)。海南文昌中华蜜蜂在后翅钩数、翅脉角A4、L13表现最大,与沂蒙山区中华蜜蜂差异显著(P<0.05)。
2.2 主因子分析

图1 主因子分析散点图

图2 主成分分析碎石图
对沂源、蒙阴和文昌三个地区共15个群体中华蜜蜂样本的43项形态特征进行了详细的测定。
采样地三个地区的中华蜜蜂43项形态特征一同进行主因子分析,提出了三个特征值较高的因素。第一因素主要包含第二背板宽、前颌长、第七腹板宽度等,第二因素主要包含上唇色度、翅脉角G18、肘脉指数等指标,第三因素主要有前翅长等。
2.3 聚类分析
利用3个地区的15群中华蜜蜂三个主要因素进行聚类发现15群体共聚为四类,其中YY7、MY3、YY3、MY4、YY4 与 YY5 聚为第一类,YY1、YY8、YY2、YY6四个群体聚为第二类,MY1、MY2聚为第三类,海南文昌三个群体HN1、HN2、HN3聚为第四类(图3)。
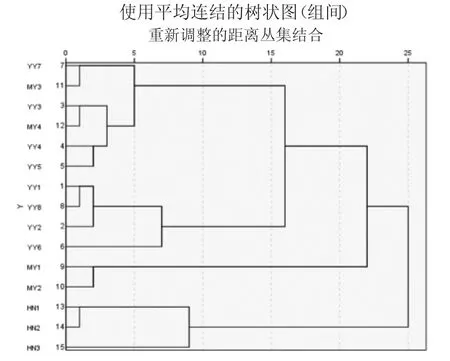
图3 沂源、蒙阴及文昌中华蜜蜂聚类分析图
2.4 判别分析

图4 主因子判别分析点图
将以上15个群体进行判别分析,结果为YY7与MY3、YY 与 MY4、YY1与 YY8、HN1与HN2遗传距离最近,YY7与HN3两地中华蜜蜂形态特征中心点相距最远。
而判别预测分析结果也与聚类结果基本验证了聚类分析结论的可靠性。
2.5 关联分析

图5 以主因子1为因变量与环境因子的回归分析图
采样地中华蜜蜂三个主成分与环境因子进行相关分析,主成分1与海拔、纬度呈极显著正相关(P<0.01),与平均气温和降雨量呈显著负相关(P<0.01);主成分2与温度相关;主成分3与年均降雨量相关,但两者与生态因子的相关不显著,故取因子1与纬度、经度、海拔、年均降雨量、年均气温等生态因子分析。以主成分为因变量,环境因子为自变量进行回归分析发现,所采集的三个地区中华蜜蜂主成分1与环境因子呈线性相关,与纬度、海拔、平均气温呈极显著正相关(相关系数分别为 r=0.963,r=0.826,r=0.672)(P<0.01);与经度、年均降雨量呈显著正相关(相关系数为r=0.598,r=0.568)(P<0.05)。
进一步将所有采样地中华蜜蜂形态特征与环境因子进行相关分析发现:经度与19项形态特征相关,与第二背板色度、第三背板色度、第四背板色度、第四背板长、第七腹板宽度、第四腹板长度、后翅钩数、翅脉角A4、翅脉角 K19、翅脉 L13、翅脉 N23、胫节长、基跗节长、基跗节宽、前颏、总长呈极显著正相关(P<0.01),相关系数分别为 r=0.943、r=0.883、r=0.652、r=0.651、r=0.681、r=0.689、r=0.784、r=0.672、r=0.721、r=0.679、r=0.641、r=0.691、r=0.746、r=0.819、r=0.717;与中唇舌、第五背板绒毛带宽度、翅脉角J16、翅脉角O26呈显著相关 (P<0.05),相关系数分别为 r=0.598、r=0.568、r=0.532、r=0.546。
纬度与21项形态特征相关,与第二背板、第三背板、第四背板、第七腹板宽度、第四腹板长度、翅脉角A4、翅脉角 K19、翅脉角 L13、翅脉角 N23、胫节长、基跗节宽、前颏、吻总长呈极显著正相关(P<0.01),相关系数分别为 r=0.951、r=0.890、r=0.664、r=0.667、r=0.691、r=0.712、r=0.795、r=0.678、r=0.732、r=0.667、r=0.730、r=0.825、r=0.729。纬度与中唇舌、第五背板绒毛带宽度、后翅钩数、翅脉角B4、翅脉角J16、翅脉角O26、基跗节长呈显著相关(P<0.05),相关系数为 r=0.567、r=0.635、r=0.516、r=0.528、r=0.564、r=0.631、r=0.616。
海拔与18项形态特征相关,与第二背板、第三背板、第四背板、第七腹板宽度、第四腹板长度、蜡镜长、前翅长度、翅脉角A4、翅脉角K19、翅脉N23、胫节长、后颏、前颏、总长呈极显著正相关(P<0.01),相关系数为r=0.837、r=0.683、r=0.652、r=0.701、r=0.769、r=0.686、r=0.648、r=0.679、r=0.705、r=0.699。海拔与第四背板、第四腹板长度、蜡镜长、前翅长、基跗节长、基跗节宽、中唇舌呈显著相关(P<0.05),相关系数为 r=0.522、r=0.519、r=0.605、r=0.562、r=0.543、r=0.577、r=0.618。
降雨量与9项形态特征相关,与翅脉角A4呈极显著正相关(P<0.01,相关系数为r=0.714),与第二背板、第四腹板长度、翅脉角D7、翅脉角K19、肘脉长b、肘脉指数、前颏、中唇舌、吻总长呈显著相关(P<0.05),相关系数为 r=0.551、r=0.542、r=0.572、r=0.528、r=0.564、r=0.631、r=0.616。
平均气温与7项形态特征相关,与第三背板、基跗节宽、后颏呈极显著正相关(P<0.01,相关系数为r=0.678、r=0.856、r=0.796),与第二背板、第七腹板宽度、后翅钩数、翅脉角L13、基跗节长、基跗节宽、中唇舌呈显著相关(P<0.05),相关系数为 r=0.640、r=0.633、r=0.5、r=0.528、r=0.564、r=0.631、r=0.616。
同质性检验显示三个地区主因子在各地区间方差分析说明主因子1(F=0.604,P>0.05),主因子2(F=3.790,P<0.05)和因子 3(F=4.721,P<0.05)在各地区间差异极显著结果是符合方差齐性假设(表3)。

表3 主因子同质性检测

图6 主因子1-3在不同地区间的平均值
主因子在各地区间的多重比较显示主区间均有差异(图 6)。
主因子1在三个地区间差异显著(P<0.05),主因子2在沂源和蒙阴之间差异不显著(P>0.05),而与文昌差异显著(P<0.05),在主因子3的平均值的图上显示沂源与其他两个地区间有显著差异(P<0.05)。
3 讨论
沂源、文昌、蒙阴3个采样地的中华蜜蜂一同进行主成分因子散点分析,第一因素里除有个别地区表现极大值和较小值以外,其它地区蜜蜂资源形态大小特征和分布比较均衡。
聚类分析结果显示沂源、蒙阴、文昌3地区基本按照地理分布的远近聚类,如沂源与蒙阴两地中华蜜蜂遗传距离较近,而海南文昌与山东地区的中华蜜蜂却有一定的遗传距离,沂源与蒙阴地理位置以及气候相似,所以两地中华蜜蜂资源形态相似,并且因两地区中华蜜蜂的形态特征趋于接近,因此聚类结果显示沂源2和蒙阴3较早发生聚合。值得注意的是,海南文昌蜜蜂在三个采样地区的中华蜜蜂中的遗传距离最远,而且其形态特征有很大保守性,显示了海南文昌与其他地区具有显著的差异,且判别分析证明了上述分类结果具有的可信性。
关联分析显示了3个地区的中华蜜蜂有关形态特征的主成分与环境因子相关,与体型小有关的主因子1随海拔的升高和纬度的北移而增大,且随年均气温和年均降雨量的升高而减小,而体形大小与经度和年均降雨量没有显著相关关系。随着海拔的升高和纬度的北移年气温会降低,中华蜜蜂为了御寒,需要不断的补充食物,以满足生命需求而导致个体发育较低,采蜜能力强,飞行能力强,体形较大。在国际上Friedrich Ruttner和Stefen Fuch等研究北纬360西方蜜蜂的自然地理形态趋化[12,13],及Verma等研究印度东方蜜蜂的地理形态趋化性上得出过相同的结论,并与HePburn研究非洲也门不同海拔条件下的非洲蜜蜂形态得出的变异结论一致[13-15],与体色有关的第二因子的数据随海拔的升高而减小,体色变深,这与谭垦、王桂芝等分析的中华蜜蜂的体色与海拔的相关结论相符[3-5,7-12]。通过沂源、文昌、蒙阴三个地区的数据分析显示,中华蜜蜂的体色随经度的东移和年降雨量的增多以及年均气温的增大而增大,体色随海拔的升高和年均气温的降低,推测体色的改变是蜜蜂对自然环境的一种适应,体色深有利于吸收阳光起到保持体温的作用。
近年来,蜜蜂形态测定引入了很多新的测定方法,如翅脉几何形态测量法[16]。光学技术及测定软件的联合开发与应用为蜜蜂形态测定提供了有力条件,埃及的SPT半自动的测定技术(Scan Phototechnique)[17]更加省时准确。
4 结论
1.小盾片Sc区的色度文昌中华蜜蜂最浅(7.578±0.567),沂源中华蜜蜂在吻长(5.134±0.050)、第 7 腹板长(2.321±0.032)等5个指标上表现出最长,蒙阴中华蜜蜂前翅长(8.489±0.099)与宽(2.925±0.043)2项指标上表现最大,文昌中蜂在后翅钩数(19.022±0.920)、翅脉角 A4(32.393±0.150)、L13(21.025±2.182)表现最大。
2.42个形态特征共提出了三个特征值较高的因素。
3.将沂源、蒙阴、文昌中华蜜蜂进行聚类,结果YY7、MY3、YY3、MY4、YY4 与 YY5 为第一类,而 YY1、YY8、YY2、YY6为第二类,MY1和 MY2聚为第三类,HN1、HN2、HN3聚为第四类,最终15群聚为四类。而且沂源与蒙阴部分地区中华蜜蜂具有亲缘性。
4.主成分1与海拔、纬度呈极显著正相关(P<0.01),与年均气温呈极显著负相关(P<0.01),与年平均降雨量呈显著负相关(P<0.05);主成分2和主成分3与生态因子相关不显著(P>0.05)。
[1]杨冠煌.中华蜜蜂[M].北京:中国农业出版社,2000:1-56.
[2]龚一飞,张其康.蜜蜂分类与进化[M].福州:福建科技出版社,2000:18-22.
[3]王桂芝,石巍.山西东方蜜蜂形态类群的研究[J].江西农业大学学报,2008,30(3):411-417.
[4]王桂芝,石巍.河南东方蜜蜂形态多样性研究[J].江西农业大学学报,2008,30(6):989-996.
[5]王桂芝,石巍.黄河中下游东方蜜蜂种质资源工蜂色型多样性研究[J].江西农业大学学报,2009,31(5):818-825.
[6]Ruttner F.Biogeography and taxonomy of honeybees[M].Berlin:Spfnger-Vedag,1988:68-284.
[7]谭垦,张炫.云南省东方蜜蜂形态学研究[J].蜜蜂杂志,2001(6):3-4.
[8]Tan K,Stefan F,Nikolaus K,et al.Morphological characterization of Apis cerana in the Yunnan Province of China [J].Apidologie,2003,34(6):553-561.
[9]Radloff S E,Hepburn H R,Stefan F,et al.Multivariate morphometfic analysis of Apis cerana populations of oceanic Asia[J].Apidologie,2005,36(3):127-140.
[10]Tan K,Marina D M,Stefan F.Morphologocal and molecular studies of Ap cerana Fabr.(Apidae,Hymenoptera)in China[J].Systematics and Biodiversity,2006,4(4):473-482.
[11]Tan K.Environmentally-induced developmental efects on morphometrie characters of workers in Apis cerana colonies[J].Apidology,2007,36(3):289-295.
[12]Tan K,Hepburn H R,Radlof S E.Multivariate morphometric analysis of the Apis cerana of China [J].Apidologie,2008,39(3):343-353.
[13]Franck P,Gamery L,Loiseau A,et al.Genetic diversity of the honeybee in Africa:Miemsatellite and mitochondrialdata [J].Heredity,2001,86(4):420-430.
[14]Radlof S E,Hepburn H R,Tan K.Multivariate morphometric analysis of Apis cerana of southern mainland Asia[J].Apidologie,2004,35(1):475-492.
[15]Franck P,Gamery L,Celebrano G,et al.Hybrid origins of honeybees from Italy(A.me Wera Ligustica)and Sicily(A.m.sicula)[J].Molecular Ecology,2000,9(7):907-921.
[16]郑秀娟,周姝婧,徐新建,等.翅脉几何形态测量法划分东方蜜蜂种群的方法研究 [C].中国养蜂学会蜜蜂饲养管理专业委员会第17次学术会议论文集,2012.
[17]张祖芸,余玉生,杨若鹏,等.微小生物形态测定仪在蜜蜂形态学中的应用[J].中国蜂业,2012,63(10-12):23-26.
Identification and classification of Apis cerana cerana in Yimeng mountain area and Wenchang
Gong Xue1,Jiang Fengtao2,Lou Delong2,Gao Jinglin3,Li Yancan1,Ma Lanting1,
Zhou Yanzhong4,Wang Guizhi1
(1 Shandong Agricutrural University College of Animal Science and Veterinary Medicine,Taian 271018,China;2 Shandong Apis Breeding and Promotion Center,Taian 271000,China;3 Chinese Academy of Chopical Agricutrural Science Evironment and Plant Protection Institute,Haikou 571101,China;4 Shandong Yiyuan Animal Husbandry Bureau,Yiyuan 256100,China)
In this study,Apis cerana cerana (ACC)in Yimeng mountain area (Mengyin and Yiyuan)was investigated and sampled.According to the international indexes,the system measurement and analysis of 42 morphological characteristics of the ACC samples were carried out,which principal component is studied by SPSS 22,so did discriminant,clustering,mutation,and environmental factors related data analysis.And the results were compared with the ACC of Hainan Wenchang in morphology and classification.The results revealed that 15 groups of ACC were clustered in four categories,including YY7,MY3,YY3,MY4,YY4 and YY5.The second type is YY1,YY8,YY2,YY6,and MY1 and MY2.The fourth category is HN1,HN2,and HN3.Discriminant analysis supports the above results.PCA suggested that the main component 1 and the main component 2 in different regions showed significant difference.Principal component 1 was very significant positive correlated with latitude and altitude,which was in significant negative correlation with annual temperature.The bee's body showed the tendencies of increasing,as altitude increasing,latitude going north and the reducing of average annual temperature.The principal component 2 was positively correlated with longitude and annual rainfall and annual temperature,while being significantly negatively correlated with altitude.
Apis cerana cerana;morphological characteristic;classification;Yimeng mountain area
山东省蜂产业技术体系育种与资源保护(SDAIT-24-01);蜜蜂种质资源保护及优质新品系的培育(2016LZGC039);国家蜂产业技术体系泰安综合试验站项目(CARS-45-SYZ9)
龚雪(1991-),女,硕士研究生,主要从事蜜蜂遗传资源保护,E-mail:790143495@qq.com
王桂芝(1966-),女,博士,主要从事蜜蜂育种与遗传资源保护,E-mail:wgz@sdau.edu.cn
