miRNA378*表达对柯萨奇B3病毒感染乳鼠心肌细胞凋亡、网腔钙结合蛋白、内质网应激伴侣蛋白表达的影响*
2017-10-14刘晓翠崔晓雪邱祥春魏成喜
赵 明, 刘晓翠, 崔晓雪, 邱祥春, 王 羽,3△, 魏成喜,3△
(1. 内蒙古民族大学第一临床医院, 2. 内蒙古民族大学, 3. 内蒙古民族大学医学院药物化学与药理学研究所, 通辽 028000)
miRNA378*表达对柯萨奇B3病毒感染乳鼠心肌细胞凋亡、网腔钙结合蛋白、内质网应激伴侣蛋白表达的影响*
赵 明1, 刘晓翠2, 崔晓雪2, 邱祥春1, 王 羽2,3△, 魏成喜2,3△
(1. 内蒙古民族大学第一临床医院, 2. 内蒙古民族大学, 3. 内蒙古民族大学医学院药物化学与药理学研究所, 通辽 028000)
目的研究沉默miRNA378*表达对柯萨奇B3病毒(CVB3)感染心肌细胞凋亡、内质网应激、网腔钙结合蛋白(calumenin)影响。方法原代培养乳鼠心肌细胞分为:对照组(正常细胞)、柯萨奇病毒感染组(正常细胞+柯萨奇B3病毒)、miRNA378*沉默对照组(正常细胞+柯萨奇B3病毒+转染miRNA378*空质粒)、miRNA378*沉默组(正常细胞+柯萨奇B3病毒+转染miRNA378*沉默质粒),各组细胞分别转染和感染处理后置37℃、CO2培养箱中培养3 d。检测细胞α-平滑肌肌动蛋白(α-SMA)、细胞凋亡率、网腔钙结合蛋白、葡萄糖调节蛋白78(GRP78)及内质网应激信号通路因子激活转录因子6(ATF6)、转录因子C/EBP同源蛋白(CHOP)的表达。结果通过检测ɑ-SMA蛋白,证实分离乳鼠细胞为心室肌细胞。TUNEL法检测不同组心室细胞凋亡情况发现,柯萨奇病毒感染组心室肌细胞凋亡明显,与柯萨奇病毒感染组心肌细胞相比较,miRNA378*沉默组心肌细胞凋亡细胞量明显减少。与柯萨奇病毒感染组比较, Calumenin表达减少(P<0.01),而GRP78、ATF6、CHOP表达增加(P<0.01)。结论CVB3病毒感染心肌细胞作用与miRNA378*,引发内质网应激并激活信号通路因子,心肌细胞凋亡增加。
心肌细胞;乳鼠;miRNA378*;柯萨奇B3病毒;内质网应激;网腔钙结合蛋白 ;凋亡
病毒性心肌炎(viralmyocarditis, VMC)是由病毒(主要由柯萨奇病毒)感染引起的心肌局限性或弥漫性的急性或慢性炎性病变[1],病变可累及心肌细胞实质和/或间质,导致心肌收缩力减低,射血分数下降甚至无效射血,最终导致心力衰竭[2]。研究发现心力衰竭患者心脏中存在心肌细胞凋亡, 致使存活的心肌工作细胞数量减少,心脏射血能力降低,导致心力衰竭[3]。心肌细胞凋亡途径众多,内质网应激反应通过其凋亡信号通路所介导心肌细胞凋亡是新近发现的凋亡途径[4]。 网腔钙结合蛋白(calumenin)是一种位于心肌细胞肌浆网中具有调节钙稳态作用的钙离子结合蛋白。microRNAs (miRNAs)是由内源基因编码的一类长约22个核苷酸的非编码单链分子RNA, 参与转录后基因表达调控,抑制靶mRNA转录后的表达[5]。文献报道:miRNA-378*能够抑制H9c2心肌细胞calumenin表达[6]。但miRNA-378*对于柯萨奇B组病毒感染的心肌细胞中的凋亡、calumenin、内质网应激及其凋亡信号通路是否具有作用以及是否通过抑制靶蛋白转录后的表达来影响心肌细胞凋亡的整个通路尚不明确。本实验拟上调CVB3病毒感染心肌细胞miRNA-378*表达,观察其对细胞凋亡、calumenin、ERS及其信号通路因子影响,进一步揭示病毒性心肌炎并发心力衰竭发病机制,为在临床工作提供新的理论基础。
1 材料与方法
1.1 动物药品与仪器
1~3 d Balb/c新生乳鼠购置吉林大学基础医学院动物中心, 许可证号:scxk(吉)2011-0004; Calumenin antibody购于Bioss公司, GRP78 antibody,PERK antibody,P-PERK antibody,eIF2ɑ antibody,ATF4 antibody,CHOP antibody,购于Boster公司,内参抗体β-actin购于wanleibio公司,慢病毒转染miRNA-378*沉默质粒由沈阳万类生物科技公司合成;柯萨奇B3病毒,Woodruf亲心肌株由吉林大学农牧学院提供。
1.2 乳鼠心肌细胞的分离与培养
将乳鼠心脏放入培养皿中,将清洗后的心脏组织剪碎至1~3 mm3小块。加入0.1%Ⅱ型胶原酶以及0.1%胰蛋白酶混合液,37℃消化25 min。离心7 min去上清。用PBS重悬沉淀,离心5 min,用培养基重悬细胞,过200目筛网去除大块组织。离心去上清后加入1 ml的培养基重悬细胞,放置到培养板中。2 h差速贴壁法可去除大量的成纤维细胞,将剩余较纯的心肌细胞放置在培养板。24 h后按实验内容将细胞接种于6孔板中,细胞浓度为5×104cells/ml,置于37℃,5% CO2的培养箱内培养过夜,24 h可进行转染。
1.3 免疫组织化学方法鉴定乳鼠培养心肌细胞
各组乳鼠培养心肌细胞进行免疫组化鉴定: 0.1%TritonX-100孵育;3%过氧化氢孵育;血清封闭; 一抗孵育;二抗孵育;辣根酶标记;DAB显色;苏木素复染;镜检。
1.4 细胞接种与培养
细胞培养于10%胎牛血清的DMEM完全培养基中,置于37℃,5% CO2的培养箱内培养。隔天换液处理,接种密度为70%为宜,便于病毒感染。
1.5 慢病毒转染
(1)准备病毒:取出4℃保存的病毒,使用前轻轻摇匀。(2)感染目的细胞:病毒准备好之后,从培养箱中拿出细胞开始实验。(3)使用移液器吸取准确体积(病毒数与细胞数比值为100∶ 1)的病毒液加入准备好的培养基中。(4)在培养基中加入ploybrene(6 μg/ml)来提高病毒的感染效率。(5)吸去细胞中剩的培养基,在目的细胞和对照组细胞中分别加入计算好的病毒液。(6)混匀后放于37℃,5% CO2的培养箱内孵育过夜。(7)24 h后将含有慢病毒的培养液更换成正常培养液(含有10%血清的DMEM培养基),37℃、5% CO2继续培养。
1.6 感染CVB3的乳鼠心肌细胞及实验分组
乳鼠心肌细胞按照细胞浓度为5×104cells/ml接种于6孔板,并将其置于37℃,5% CO2的培养箱内培养,心肌细胞随机分成4组,每组3孔,对照组(正常细胞)、柯萨奇病毒感染组(正常细胞+柯萨奇B3病毒)、miRNA378*沉默对照组(正常细胞+柯萨奇B3病毒+转染miRNA378*空质粒)、miRNA378*沉默组(正常细胞+柯萨奇B3病毒+转染miRNA378*沉默质粒)。其中两组细胞分别转染miRNA378*空质粒和miRNA378*沉默质粒,处理24 h后,取3组细胞(1组正常细胞和2组转染后细胞)与100 PFU的CVB3共同孵育1 h,弃上清,加入正常培养液作为CVB感染组;设立细胞对照组(仅加入培养液,不进行病毒感染),将4组心肌细胞在37℃,置于CO2培养箱中3 d。
1.7 TUNEL法检测各组心肌细胞凋亡
各组心肌细胞经过TUNEL法标准实验步骤:透化;PBS漂洗;封闭;滴加TUNEL反应液50 μl;保湿、避光、37℃孵育60 min;滴加Converter-POD 50 μl;保湿、37℃孵育30 min;DAB显色:加DAB底物50 μl;苏木素复染;显微镜下观察染色效果,400×镜下拍照。
1.8Westernblot检测各组心肌细胞calumenin蛋白、内质网应激伴侣蛋白GRP78及ATF6、CHOP表达
各组乳鼠心肌细胞经下列实验步骤:蛋白质抽提;蛋白质定量;取15 μl的样品进行SDS-PAGE;转印至PVDF膜;封闭;孵育一抗;孵育二抗;ECL底物发光;图像保存。普通抗体的封闭液、一抗孵育液、二抗孵育液为5%脱脂奶粉溶液。检测各组心肌细胞calumenin蛋白(1∶1 000)、葡萄糖调节蛋白78(glucose regulated protein 78, GRP78)(1∶1 000)及内质网应激信号通路因子激活转录因子6(activation transcription factor 6, ATF6)(1∶1 000)、转录因子C/EBP同源蛋白(transcription factors c/ebp homologue protein,CHOP)(1∶1 000)表达。
1.9 统计学处理
2 结果
2.1 乳鼠心肌细胞鉴定
培养的乳鼠心肌细胞经免疫组化实验,检测出心肌细胞特有的ɑ-SMA蛋白,确定为心室肌细胞(图1)

Fig.1Mouse myocardium celle(×400) A: Primary culture observed under microscope; B: Immunocytochemistry of culturing myocardial cell
2.2 心肌细胞凋亡的比较
用TUNEL 法检测对照组心肌细胞仅有少量凋亡的细胞(凋亡率6.83%±0.71%),柯萨奇病毒感染组心肌细胞凋亡明显,凋亡率60.37%±7.20% (P<0.01), 与柯萨奇病毒感染组心肌细胞相比较,miRNA378*沉默组心肌细胞凋亡细胞量明显减少,凋亡率为46.6%±8.3%(P<0.01,图2)。
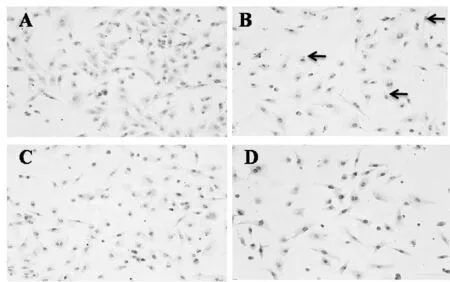
Fig.2Myocardial apoptosis in each group(×200) A: Control group; B: Coxsackie virus B3 infecton group; C: miRNA378*silence control group; D: miRNA378*silence group
2.3心肌细胞calumenin蛋白、内质网应激伴侣蛋白GRP78及ATF6、CHOP表达
与对照组相比较,柯萨奇病毒感染组心肌细胞calumenin蛋白表达减少(P<0.01),GRP78、ATF6、CHOP表达增加(P<0.01);与柯萨奇病毒感染组相比较,miRNA378*沉默组calumenin蛋白表达显著减少(P<0.01); miRNA378*沉默组GRP78、ATF6、CHOP表达显著增加(P<0.01,图3,表1)。

Fig.3Expression change of Calumenin, GRP78 ATF6 and CHOP A: Control group; B: Coxsackie virus B3 infecton group; C: miRNA378*silence control group; D: miRNA378*silence group; GRP78: Glucose regulated protein 78; ATF-6: Activation transcription factor 6; CHOP: Transcription factors c/ebp homologue protein


GroupCalumeninGRP78 ATF6 CHOP A2265±192880±79753±82673±42B1335±119∗∗1693±141∗∗1355±121∗∗1288±137∗∗C1541±1341625±1331091±117716±126D613±42##2502±315##2611±221##2698±244##
A: Control group; B: Coxsackie virus B3 infecton group; C:miRNA378*silence control group; D: miRNA378*silence group; GRP78: Glucose regulated protein 78; ATF6: Activation transcription factor 6; CHOP: Transcription factors c/ebp homologue protein
**P<0.01vsA group;##P<0.01vsB group
3 讨论
心肌细胞calumenin与RyR2通道及钙泵(SERCA2a) 相互作用来调节心肌细胞钙释放、回摄与储存,共同维持钙循环稳态[7]。文献报道,病毒性心肌炎时,心肌细胞calumenin蛋白不但表达明显减少,而且与SERCA2a结合能力减弱,从而使心肌细胞钙循环稳态失衡,心脏功能降低[8]。同时,病毒性心肌炎又可以导致心肌细胞发生内质网应激(ERS),并激活ERS凋亡信号通路引起心肌细胞凋亡,引发心力衰竭[9]。研究发现calumenin可以缓解心肌细胞ERS并减少ERS介导的心肌细胞凋亡数量[10],重度心力衰竭患者miRNA-378*表达显著下降,随着患者心功能好转,miRNA-378*表达也明显增加[11]。
miRNA-378 和miRNA-378*是一个基因出来的产物,叫正义链和反义链 。microrna前体有发卡结构,在成熟过程中发卡结构被剪掉,形成miRNA-378 和miRNA-378*,也叫左臂和右臂。通常正义链有功能,反义链无功能被降解。但是近年来,人们发现反义链有时候有功能,可以协助正义链功能,也可以拮抗正义链,或者和正义链功能毫无关系。本研究根据上述资料和本课题组研究结果,提出如下科学假说:柯萨奇B3病毒通过干预心肌细胞miRNA-378*表达来影响calumenin表达,进而引起ERS并通过ERS凋亡信号通路:ATF6→CHOP介导心肌细胞凋亡。
本实验在成功培养原代培养乳鼠的心室肌细胞的基础上,采用干扰技术下调miRNA-378*的表达,并与CVB3病毒感染处理后的心室肌细胞。TUNEL检测发现感染CVB3病毒的心室肌细胞凋亡率提高,而下调miRNA-378*后,心室肌细胞的凋亡率有所下降。证明下调miRNA-378*的表达可缓解CVB3病毒所致的心室肌细胞凋亡。内质网应激是诱发凋亡的机制之一。本研究进一步探讨miRNA-378*的作用是否与内质网应激相关,检测内质网应激分子伴侣GRP78及效应蛋白ATF6,CHOP,发现GRP78,ATF6,CHOP在CVB3病毒感染的心室肌细胞中均上调,而下调miRNA-378*后,3个蛋白表达量增加。说明miRNA-378*缓解心室肌细胞凋亡不通过内质网应激通路。最后,calumenin蛋白的表达也进行了检测,下调miRNA-378*的心室肌细胞可提高因感染CVB3病毒而导致的表达减少。本研究结果部分揭示miRNA-378*可缓解病毒性心肌炎所致的凋亡,而与内质网应激无关,具体作用还需以后的实验中进一步阐明。
[1] 刘 磊, 王洪军, 赵 明, 等. 病毒性心肌炎所致小鼠心力衰竭心肌组织内质网应激相关的凋亡关系的研究[J]. 中国应用生理学杂志, 2014, 30(4): 465-468.
[2] 牛晓峰, 赵雅君, 赵 明, 等. 黄芪总黄酮通过抑制内质网应激对小鼠病毒性心肌炎保护作用的实验研究[J]. 临床心血管病杂志, 2015, 31(2): 129-132.
[3] Boon RA, Dimmeler S. MicroRNAs in myocardial infarction[J].NatRevCardiol, 2015, 12(3): 135-142.
[4] 周晓敏, 陶谢鑫, 赵 明, 等. 黄芪总黄酮与丹参酮ⅡA磺酸钠对病毒性心肌炎小鼠心肌细胞内质网伴侣蛋白及L-型钙通道表达的作用[J]. 临床心血管病杂志, 2015, 31(3): 254-256.
[5] 黄 侠, 邱祥春, 赵 明. 网腔钙结合蛋白的研究进展[J]. 生理科学进展, 2015, 46(6): 458-460.
[6] Mallat Y, Tritsch E, Ladouce R,etal. Proteome modulation in H9c2 cardiac cells by microRNAs miR-378 and miR-378*[J].MolCellProteomics, 2014, 13(1): 18-29.
[7] 赵雅君, 王伊林, 赵 明, 等. 阿霉素致心肌细胞凋亡时网腔钙结合蛋白的变化[J]. 临床心血管病杂志, 2015, 31(8): 895-898.
[8] Zhou X, Xin Q, Wang Y,etal. Total flavonoids of astragalus plays a cardioprotective role in viral myocarditis[J].ActaCardiolSin, 2016, 32(1): 81-88.
[9] 王洪军, 于 淼, 赵 明, 等. 黄芪总黄酮对柯萨奇B3病毒感染心肌细胞内质网应激及促凋亡信号因子作用实验研究[J]. 临床心血管病杂志, 2016, 32(1): 70-73.
[10]Lee JH, Kwon EJ, Kim DH,etal. Calumenin has a role in the alleviation of ER stress in neonatal rat cardiomyocytes[J].BiochemBiophysResCommun, 2013, 439(3): 327-332.
[11]Ooi JY, Bernardo BC, McMullen JR. The therapeutic potential of miRNAs regulated in settings of physiological cardiac hypertrophy[J].FutureMedChem, 2014, 6(2): 205-222.
ThecorrelationresearchonmiRNA378*andcalumenin,endoplasmicreticulumstress,apoptosisinsucklingmousemyocardialcellsinfectedwithcoxsackievirusB3
ZHAO Ming1, LIU Xiao-cui2, CUI Xiao-xue2, QIU Xiang-chun1, WANG Yu2,3△, WEI Cheng-xi2,3△
(1. The Inner-Mongolia National University First Clinical Hospital, 2. The Inner-Mongolia National University, 3. The Inner-Mongolia National University Medical Chemistry and Pharmacology Institute, Tongliao 028000, China)
Objective: To investigate the effects of silencing miRNA378*on apoptosis, endoplasmic reticulum stress and calumenin of cardiomyocyte with coxsackie virus B3 (CVB3) infection.MethodsPrimary cultured suckling mouse myocardium were divided into control group (normal cell), coxsackie virus infection group (normal cell and coxsackie virus B3), miRNA378*control group (normal cell +coxsackie virus B3+miRNA378*empty plasmid), miRNA378*silencing plasmid group(normal cells + coxsackie virus B3 + miRNA378*silencing plasmid). Four groups of cells were transfected, infected and treated in CO2incubator at 37℃. The α-SMA protein, cell apoptosis rate, calumenin, glucose regulated protein 78 (GRP78) , activation transcription factor 6(ATF6) and transcription factors c/ebp homologue protein (CHOP) in endoplasmic reticulum were analyzed.ResultsBy detecting α-SMA protein, the isolated suckling mouse ventricular myocardium were confirmed. TUNEL detection of different groups of ventricular cell apoptosis found that coxsackie virus group of ventricular myocytes apoptosis was significant. Compared with the coxsackie virus infection group of myocardial cells, miRNA378*silencing plasmid expression of cardiomyocyte apoptosis cells significantly reduced(P<0.01). The expressions of GRP78, ATF6 and CHOP were increased compared with those infected by Coxsackie virus infection (P<0.01), while the expressions of calumenin were decreased (P<0.01).ConclusionCVB3 infected myocardial cells effected miRNA378*expression. It can trigger endoplasmic reticulum stress and activates signaling pathway factor and increase myocardial cell apoptosis.
cardiomyocytes; suckling mice; miRNA378*; coxsackie virus B3; endoplasmic reticulum stress; calumenin; apoptosis
R331.3;R-332
A
1000-6834(2017)04-304-04
内蒙古自治区自然科学基金(2015MS08153)
2016-06-07
2016-11-21
△
Tel: 0475-8314234; E-mail: shen348@126.com; weichengxi1224@163.com
10.12047/j.cjap.5461.2017.074
