糖果鸢尾花粉活力和柱头可授性研究
2017-09-25阮丽丽高亦珂杨占辉史言妍张启翔北京林业大学园林学院花卉种质创新与分子育种北京市重点实验室国家花卉工程技术研究中心城乡生态环境北京实验室北京100083
阮丽丽,高亦珂,刘 玮,杨占辉,史言妍,张启翔(北京林业大学 园林学院/花卉种质创新与分子育种北京市重点实验室/国家花卉工程技术研究中心/城乡生态环境北京实验室,北京 100083)
糖果鸢尾花粉活力和柱头可授性研究
阮丽丽,高亦珂,刘 玮,杨占辉,史言妍,张启翔
(北京林业大学 园林学院/花卉种质创新与分子育种北京市重点实验室/国家花卉工程技术研究中心/城乡生态环境北京实验室,北京 100083)
观测糖果鸢尾及其亲本射干(Irisdomestica)和野鸢尾(I.dichotoma)开花散粉时期花粉生活力和柱头可授性的变化规律,为提高人工授粉效率并为糖果鸢尾杂交育种提供依据。采用悬滴法测定花粉活力,用联苯胺-过氧化氢测定柱头可授性,结合田间花部性状随着时间变化的实际观测进行研究。刚散粉时,糖果鸢尾的花粉活力最大,随后逐渐下降。在花朵初放时柱头的可授性最弱,之后呈现先上升再下降的趋势。柱头萎蔫后,可授性低或丧失。花粉活力与柱头可授性强期没有重叠,且花粉比柱头先成熟,是糖果鸢尾避免自花传粉的适应机制;散粉1~2 h内收集花粉,并在柱头可授性最强时进行授粉,可以提高杂交成功率。
糖果鸢尾;花粉生活力;柱头可授性
糖果鸢尾(Pardancandanorrisii)是以野鸢尾(Irisdichotoma)和射干(Irisdomestica)为亲本,经不断杂交、回交等方式获得的后代群体的统称[1]。因其花被会在花朵闭合次日翻卷,连着子房就像糖果,故名Candy lily,中文译为糖果鸢尾。糖果鸢尾的亲本野鸢尾和射干均原产中国,具有耐旱、耐寒、耐瘠薄等优良抗逆性,是十分优良的育种材料[2]。但花色单一,限制了它们在园林中的应用。糖果鸢尾花色变异丰富,具有从白色到深紫色、黄色、橙色的花色,并且花量大于射干和野鸢尾,是新型商业品种,受到全世界育种家的瞩目。糖果鸢尾中如复色、重瓣、无斑点等特异新类型的获得,主要通过自交和回交的手段选育获得,因此研究糖果鸢尾及亲本花粉生活力和柱头可授性在开花后不同时间内的变化,将为提高糖果鸢尾的人工授粉效率和杂交育种提供理论依据。
1 材料与方法
1.1 材 料
2008年,野鸢尾采自北京鹫峰国家森林公园,射干购自东升乡种苗圃,栽培于昌平区国家花卉工程技术研究中心苗圃(N40°09′,E116°26′)。野鸢尾花白色或蓝紫色,外花被上部反折,花具棕褐色斑纹,花期为7-8月。射干花橙红色具紫褐色散生斑点,内外花被均无反折,花期7-8月。
2009年8月,将1株射干的花粉授于1株野鸢尾,得到1株F1植株,开紫色花,明显区别于亲本。2011年,将此株F1植株自花授粉188朵,获得120个杂交蒴果及大量F2代种子。2012-2015年,将野鸢尾、射干以及F1不断杂交,共获得201株F1(野鸢尾为母本,射干为父本)、91株F2、80株F1×S (F1作母本与射干回交)和66株Y×F1(野鸢尾作母本与F1杂交)。
1.2 花粉和柱头形态观察
2014年和2015年6月下旬-9月上旬,试验群体包括14株野鸢尾、6株射干、154株F1、91株F2、70株F1×S和66株Y×F1。在开花期,每隔0.5 h用相机拍摄群体的每株植株朵花花粉、柱头的形态变化并记录每朵花的开闭时间。
1.3 花粉活力测定
采用悬滴法测定花粉活力。预试验中,花粉培养液配方见表1。
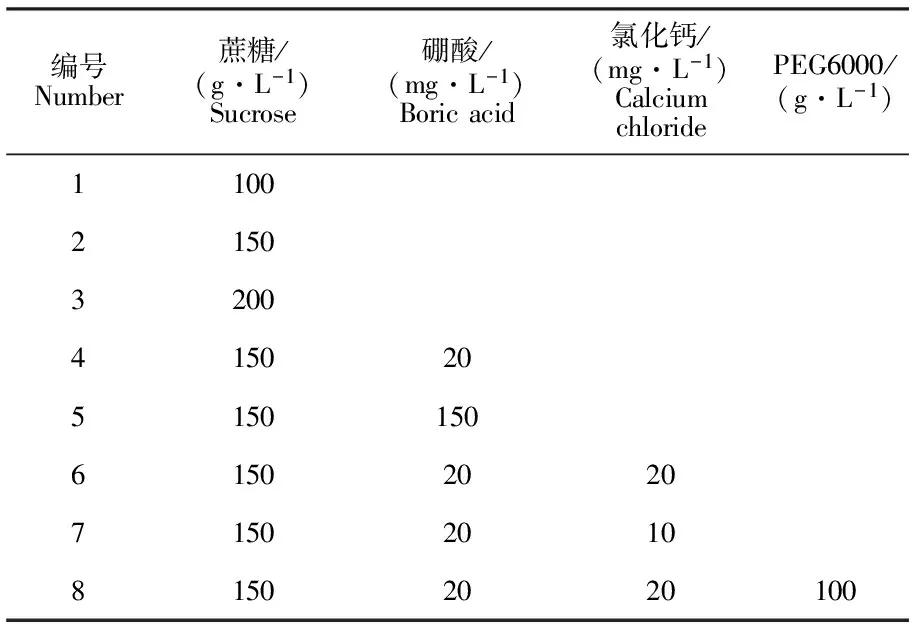
表1 不同的花粉培养液质量配方Table 1 Different formula of pollen culture medium
根据预试验结果得出最适宜的花粉培养液配方为150 g·L-1蔗糖+20 mg·L-1硼酸+20 mg·L-1氯化钙+100 g·L-1PEG6000。配好后进行高压灭菌,用离心管分装,存于4 ℃条件下待用。
花朵开放之前,选取发育程度相近的花蕾套袋隔离。并且在花朵开放后每隔1 h采集花粉,直到花朵闭合停止。将花粉离体于室温下培养24 h后在显微镜下检测并照相[3]。每样品重复3次,每重复观察3个视野。
1.4 柱头可授性测定
采用联苯胺-过氧化氢测定柱头可授性,参照李小远等[4]的方法进行,具体步骤为在花朵开放前选取发育程度相近的花蕾进行标记,并且在花朵开放后每隔1 h采集柱头,直到花朵闭合停止。将采集后的柱头置于凹槽载玻片上,滴入联苯胺-过氧化氢反应液,在显微镜下观察并照相。若反应液变蓝并有大量气泡产生,则柱头有较强可授性,若只有少量气泡产生,则柱头的可授性较低,若无气泡产生,则没有可授性。
2 结果与分析
2.1 柱头和花粉的形态变化
野鸢尾16:00开放,22:00闭合,单花花期长达6 h(表2)。花粉在开花0.5 h内开始大量散出,在自然状态下,直到花朵闭合,仍然有大量花粉保留在花药上;柱头在开花6 h后开始萎蔫变软、反卷。射干7:00开放,19:00闭合,单花花期为12 h(表2)。花粉在开花0.5 h内开始散出,并在1 h内大量散出,附在花药表面;2 h后,花粉散光,花药上没有花粉;柱头在开花10 h后开始萎蔫变软。
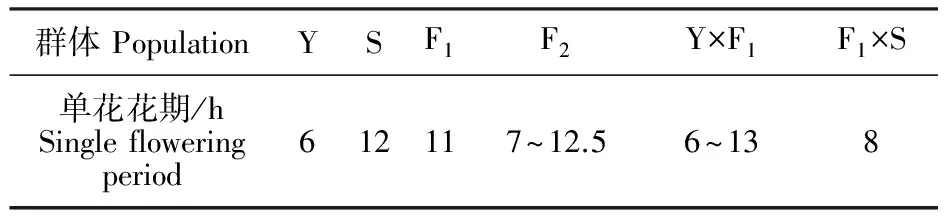
表2 糖果鸢尾及亲本单花花期Table 2 The single flowering period of Pardancanda norrisii, I.domestica and I.dichotoma
注:Y.野鸢尾;S.射干;F1.野鸢尾为母本,射干为父本;F2.F1自交;F1×S.F1作母本与射干回交;Y×F1.野鸢尾作母本与F1杂交。下表同。
Note:Y.I.dichotoma;S.I.domestica;F1.F1generation ofI.dichotomaas female parent andI.domesticaas male parent;F2.F2generation derived from F1selfing;F1×S.Backcross of F1as female parent withI.domesticaas male parent;Y×F1.Backcross of F1as male parent withI.dichotomaas female.The following table is the same.
F1植株10:00开花,21:00闭合,花期为11 h(表2)。花粉在开花0.5 h内开始大量散出,6 h后花粉散完;柱头在9 h后开始萎蔫变软、反卷。F2植株7:30-14:00开花,18:00-23:00闭合,花期为7.0~12.5 h(表2)。开花1~2 h内花粉大量散出,6~9 h花粉散完,8~10 h后柱头开始萎蔫。Y×F1植株10:00开放,18:00闭合,开花时长为8 h(表2)。开花0.5 h内花粉大量散出,并且直到花朵闭合,大量花粉仍然保留在花药表面,与野鸢尾相似。F1×S植株8:30-15:00开放,19:30-23:00闭合,单花花期长达6.0~13.0 h(表2)。花粉在0.5~1 h内开始大量散出,4~6 h散完;柱头在8~10 h后开始萎蔫变软。
糖果鸢尾与亲本的花粉在0.5~1 h内都会大量散出,柱头开始萎蔫时间与花朵闭合时间基本重叠,存在2 h内的波动。
2.2 花粉活力变化
由表3可知,亲本野鸢尾和射干的花粉萌发率都很高,最大值均高于85%。糖果鸢尾群体中花粉萌发率存在较大的差异,F2的最大值为78.85%(图1),F1×S的最大值为32.50%,F1的最大值为29.37%,Y×F1的最大值为10.79%。因此,糖果鸢尾中花粉生活力由大到小的排序为F2>F1×S>F1>Y×F1。
野鸢尾的花粉萌发率在开花1~3 h内比较高,均高于50%;4 h后下降为25.14%;6 h后降为10.74%。且花朵在6 h后闭合,因此没有再测量花粉萌发率。射干的花粉萌发率在1~5 h内一直很高,最小值为58.74%,之后再逐渐降低。
F1的花粉萌发率在1 h时最高,为29.37%,之后逐渐下降,6 h后花粉失去生活力,并且观察视野中存在很多干扁、不规则的花粉。F2的花粉萌发率在1~3 h内都很高,均大于50%,9 h后花粉失去活性。F1×S的花粉生活力在1~3 h内较高,之后一直在1%左右,生活力低。Y×F1的花粉萌发率一直很低,最大值为10.79%,10 h后花粉失去活性。因此,糖果鸢尾的花粉生活力在开花1~3 h内较高,之后逐渐降低。

表3 糖果鸢尾及亲本的花粉活力Table 3 Pollen viability of Pardancanda norrisii, I.domestica and I.dichotoma %
注:“-” 表示未测量。
Note:“-” means no measurement.

A1.3 h,箭头表示花粉管 The arrow means pollen tube; A2.6 h; A3.9 h

B1.0 h; B2.2 h,箭头表示气泡 Arrow means bubbles; B3.4 h; B4.8 h
2.3 柱头可授性
由表4可知,野鸢尾的柱头可授性在开花5~6 h 内最强,射干在开花6~8 h内最强。F1在开花6~8 h内最强,F2在开花4~6 h内最强,F1×S在开花6~7 h内最强,Y×F1在开花1~3 h 内最强(图2)。糖果鸢尾及其亲本的柱头可授性均是开花之初最弱,之后先上升再下降。
3 讨 论
花粉萌发受很多因素影响,种间、品种间甚至同一品种不同取粉时间,其花粉萌发率也有所不同[5]。本研究中,射干花粉生活力总体上高于野鸢尾,且两亲本的花粉萌发率均高于糖果鸢尾群体。相似的结果在4种杜鹃花品种(‘大鸳鸯锦’‘胭脂蜜’‘琉球红’‘小青莲’)和4种鸢尾属植物(马蔺、长白鸢尾、紫花鸢尾和溪荪鸢尾)中也曾出现[6-7]。另外,亲本和糖果鸢尾群体的花粉萌发率都会随着时间的延续而逐渐下降,这与长白鸢尾、中亚鸢尾、马蔺等植物中观察的结果相同[8-10]。可见,种、品种或同一品种的取粉时间不同,花粉萌发率也会不同。因此,要想在育种中提高杂交结实率,选择合适群体的花粉以及合适的取粉时间都是非常重要的。
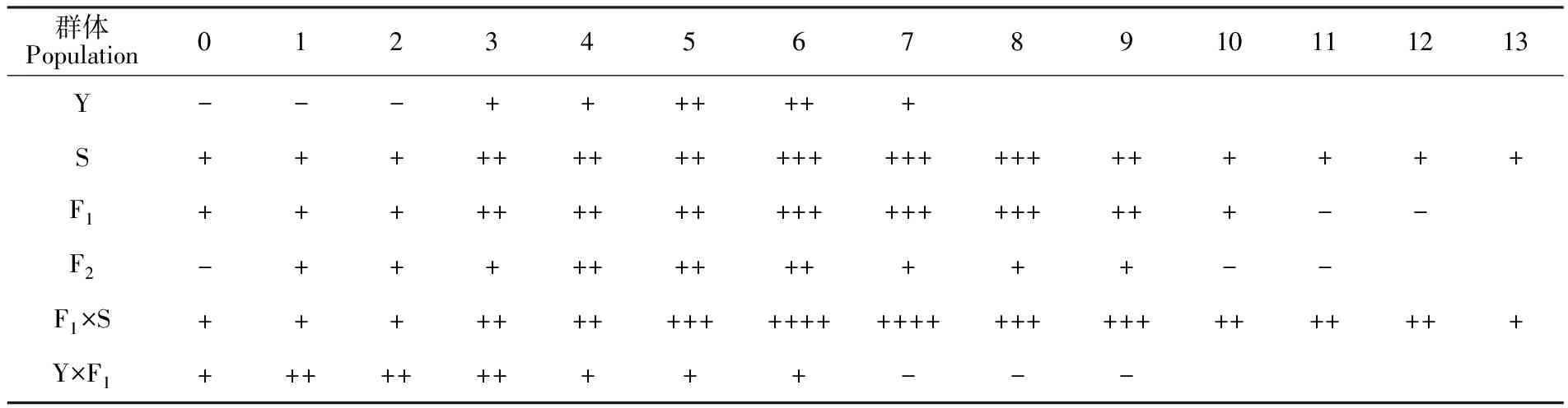
表4 糖果鸢尾及其亲本的柱头可授性Table 4 Stigma receptivity of Pardancanda norrisii, I.domestica and I.dichotoma
注:+表示柱头具有可授性;++表示柱头具较强可授性;+++/++++表示柱头具有最强可授性;-表示柱头不具有可授性。
Note:+. Stigmas have receptivity; ++. Stigmas have higher receptivity; +++/++++.Stigmas have the highest receptivity; -.No stigma has receptivity.
柱头可授性是花朵成熟过程中的一个重要时期,其在很大程度上影响自花传粉率、开花后不同阶段的传粉率、各种传粉者的相对重要性、雄性和雌性功能之间的相互干扰、不同基因型花粉之间的竞争以及配子体选择的机会等。植物的柱头可授性受单花期的长短、开花后的时间、1 d内不同时辰以及柱头分泌物的有无等影响[11]。冷蒿的柱头在开花后第2~3天的活力最高,并能保持6 d左右[12]。木地肤的柱头在第8~12天时可授性最强[13]。大花惠兰在20~40 d内的可授性最强,45 d后其柱头可授性逐渐降低[14]。本研究中,野鸢尾、射干以及糖果鸢尾由于单花花期不同,柱头开始具有最强可授性的时间和长度也不相同,但各个群体保持最强柱头可授性的时间相差不大,都在1~2 h内。
另外,柱头的可授性与其形态特征之间也有密切的对应关系[15]。多枝柽柳的柱头在呈现白色并有明显水渍状时,具有可授性;在出现萎蔫状态且柱头顶部变为浅黄色时,可授性减弱;当柱头萎缩变干且顶部变为黄色时,不具有可授性[16]。刺五加雌花和两性花的柱头只有在花柱明显伸长、柱头外翻、乳突细胞呈猫耳朵状后才具可授性[17]。本研究中,当野鸢尾、射干以及糖果鸢尾的柱头萎蔫变软的时候,柱头的可授性减弱或者丧失。
植物杂交育种主要依赖于花粉和柱头的相互协作、共同完成受精作用,因此授粉前亲本花粉育性和雌蕊发育状况对结实率有一定的影响,高活力的花粉和大量发育正常的雌蕊是保证较高结实率的重要前提条件。杨占辉等[18]和阮丽丽等[19]发现糖果鸢尾可以通过进一步自交和回交获得变异丰富的植株。毕晓颖等[20]研究表明,野鸢尾和射干间不存在杂交不亲和性,并且两者能够杂交获得真杂种;在本研究中,F1的花粉活力高时比其柱头可授性最强时早1~2 h,F2是早1~3 h,射干的花粉活力很强,是很好的父本材料,但收集不及时,花粉会被风吹散。因此,要想获得变异丰富的糖果鸢尾群体,可在花后1~2 h内收集作为父本植株上的花粉,F1的植株可在花后6~8 h内进行授粉,F2植株于4~6 h内进行授粉。此外,糖果鸢尾植株的花粉具有高活力的时间和柱头可授性强期没有重叠,且花粉比柱头先成熟,符合避免自花传粉的适应机制。
Reference:
[1] CHIMPHAMBA B B.Intergeneric hybridization betweenIrisdichotoma,pall andBelamcandachinensis,Leman[J].Cytologia,1973,38:539-547.
[2] 郭 瑛.北京地区三种鸢尾属植物的生物研究[D].北京:北京林业大学,2006.
GUO Y.Biological study of three specieces ofIrisin Beijing[D].Beijing:Beijing Forestry University,2006(in Chinese with English abstract).
[3] DAFNI A .Pollinati on Ecology[M].New York :Oxford University Press,1992:1-57.
[4] 李小远,马玉磊,刘 阳,等.小报春(Primulaforbesii)不同授粉组合亲和性研究[J].西北植物学报,2012,32(12):2426-2431.
LI X Y,MA Y L,LIU Y,etal.Compatibility of different pollination combinations inPrimulaforbesi[J].ActaBotanyBoreali-occidentalisSinica,2012,32(12):2426-2431(in Chinese with English abstract).
[5] 许建兰,马瑞娟,宋宏峰,等.不同杏品种开花生物学特性和花粉萌发率研究[J].江苏农业科学,2005,33(6):83-85.
XU J L,MA R J,SONG H F,etal.Study on the biological characteristics and pollen germination rate of different apricot varieties[J].JiangsuAgriculturalSciences,2005,33(6):83-85(in Chinese).
[6] 童 俊,周 媛,董艳芳,等.四种杜鹃花粉生活力和柱头可授性研究[J].湖北农业科学,2015,54(17):4232-4236.
TONG J,ZHOU Y,DONG Y F,etal.Study on pollen vitality and stigma receptivity of four rhododendron cultivars[J].HubeiAgriculturalSciences,2015,54(17):4232-4236(in Chinese with English abstract).
[7] 许玉凤,韩 静,海 风,等.鸢尾属植物花粉活力和柱头可授性的研究[J].北方园艺,2010(16):129-131.
XU Y F,HAN J,HAI F,etal.Study on stigma receptivity and pollen viability ofIris[J].NorthernHorticulture,2010(16):129-131(in Chinese with English abstract).
[8] 穆 丹,王 玲,卓丽环.长白鸢尾开花的生物学特性及花粉生活力[J].东北林业大学学报,2006,34(4):57-58.
MU D,WANG L,ZHUO L H.Biological charateristics of flowering and pollen viability ofIrismandshurica[J].JournalofNortheastForestryUniversity,2006,34(4):57-58(in Chinese with English abstract).
[9] 董玉芝,昝少平,李 宁,等.中亚鸢尾的花粉生活力及其授粉[J].东北林业大学学报,2003,31(6):78-79.
DONG Y ZH,ZAN SH P,LI N,etal.The pollen living ability and pollination ofIrisbloudowii[J].JournalofNortheastForestryUniversity,2003,31(6):78-79(in Chinese with English abstract).
[10] 王育青,马小春,周延林,等.马蔺开花及传粉的生物学特性[J].中国草地学报,2010,32(3):45-50.
WANG Y Q,MA X CH,ZHOU Y L,etal.Biological characteristics of flowering and pollination ofIrislacteavar.chinensis[J].ChineseJournalofGrassland,2010,32(3):45-50(in Chinese with English abstract).
[11] DAFNI A .Pollination Ecology [M] .New York :Oxford University Press,1992:59-89.
[12] 张雅荣,宛 涛,蔡 萍,等.冷蒿的开花动态与花粉活力及柱头可授性研究[J].中国草地学报,2012,34(1):108-112.
ZHANG Y R,WAN T,CAI P,etal.Observation on the flowering dynamic,pollen viability and stigma receptivity inArtemisiafrigidaWilld[J].ChineseJournalofGrassland,2012,34(1):108-112(in Chinese with English abstract).
[13] 郭红超,严 成,魏 岩.木地肤的开花动态与花粉活力及柱头可授性研究[J].草业学报,2014,23(4):87-93.
GUO H CH,YAN CH,WEI Y.Study on the flowering dynamic,pollen viability and stigma receptivitydynamic,pollen viability and stigma receptivity ofKochiaprostrate[J].ActaPrataculturaeSinica,2014,23(4):87-93(in Chinese with English abstract).
[14] 褚 怡,范义荣,张韶伊,等.大花蕙兰与国兰花粉活力及柱头可授性分析[J].浙江农林大学学报,2013,30(6):950-954.
CHU Y,FAN Y R,ZHANG SH Y,etal.Pollen vitality and stigma receptivity ofCymbidiumhybridumand Chinese orchid[J].JournalofZhejiangA&FUniversity,2013,30(6):950-954(in Chinese with English abstract).
[15] 刘林德,张 萍,张 丽,等.锦带花的花粉活力、柱头可授性及传粉者的观察[J].西北植物学报,2004,24(8):1431-1434.
LIU L D,ZHANG P,ZHANG L,etal.Pollen viability,stigma receptivity and pollinators ofWeigelaflorida[J].ActaBotanyBoreali-occidentailsSinica,2004,24(8):1431-1434(in Chinese with English abstract).
[16] 姬慧娟,尹林克,严 成,等.多枝柽柳的开花动态及花粉活力和柱头可授性研究[J].西北农林科技大学学报(自然科学版),2009,37(5):114-118.
JI H J,YI L K,YAN CH,etal.Study on floral dynamics ,pollen viability and stigma receptivity ofTamarixramosissima[J].JournalofNorthwestA&FUniversity(NaturalScienceEdition),2009,37(5):114-118(in Chinese with English abstract).
[17] 刘林德,张洪军,祝 宁,等.刺五加花粉活力和柱头可授性的研究[J].植物研究,2001,21(3):375-379,483.
LIU L D,ZHANG H J,ZHU N,etal.Pollen viability and stigma receptivity ofEleutherococcussenticosus(Araltiaceae)[J].BulletinBotanicalResearch,2001,21(3):375-379,483(in Chinese with English abstract).
[18] 杨占辉,高亦珂,刘菁芸,等.野鸢尾和射干种间杂交F2代主要性状变异分析[J].中国农业大学学报,2014,19(1):67-73.
YANG ZH H,GAO Y K,LIU Q Y,etal.Variation analysis of important ornamental traits in F2hybrids ofIrisdichotomaandI.domestica[J].JournalofChinaAgriculturalUniversity,2014,19(1):67-73(in Chinese with English abstract).
[19] 阮丽丽,高亦珂,吴 琦,等.糖果鸢尾开花结实相关性状遗传分析[J].中国农业大学学报,2016,21(4):35-41.
RUAN L L,GAO Y K,WU Q,etal.Genetic analysis on flowering and fruiting related traits inPardancandanorrisii[J].JournalofChinaAgriculturalUniversity,2016,21(4):35-41(in Chinese with English abstract).
[20] 毕晓颖,李 卉,娄 琦,等.野鸢尾和射干属间杂交亲和性及杂种鉴定[J].园艺学报,2012,39(5):931-938.
BI X Y,LI H,LOU Q,etal.Studies on inter-generic compatibility ofIrisdichotomaandBelamcandachinensisand their hybrids identification[J].ActaHorticulturaeSinica,2012,39(5):931-938(in Chinese with English abstract).
StudiesonPollenViabilityandStigmaReceptivityofPardancandanorrisii
RUAN Lili, GAO Yike, LIU Wei, YANG Zhanhui, SHI Yanyan and ZHANG Qixiang
(Beijing Key Laboratory of Ornamental Plants Germplasm Innovation & Molecular Breeding/National Engineering Research Center for Floriculture/Beijing Laboratory of Urban and Rural Ecological Environment, College of Landscape Architecture, Beijing Forestry University, Beijing 100083, China)
In order to improve artificial-pollination possibility in hybridization ofPardancandanorrisii, the morphological changes of stigma and pollen were observed and the stigma receptivity and pollen viability were tested.The benzidine-H2O2testing stigma receptivity, pollen germination test in vitro and morphological observation of stigma and pollen were adopted in the study. The pollen ofP.norrisiihas the highest viability at the beginning, then its viability declines gradually.The stigma has the lowest receptivity at the beginning, then increasing and declining again.The wilted stigma has low or null receptivity.Pollen matures earlier than stigma, and there was no overlap between the highest period of pollen viability and stigma receptivity, which is the mechanism ofP.norrisiito avoid self-pollination.Collecting pollen in 1-2 h at the beginning and pollinating at the highest period of stigma receptivity, can improve the possibility of hybridization.
Pardancandanorrisii; Pollen viability; Stigma receptivity
2016-07-12
2016-09-10
Graduate Training and Development Program of Beijing Municipal Commission of Education(No.BLCXY201627);the State Forestry Administration of the People’s Republic of China(No.2013-04-46).
RUAN Lili, female,master student.Research area:innovation and breeding of flower germplasm resources.E-mail:lilyruan2014@bjfu.edu.cn
S682.1+9
A
1004-1389(2017)09-1379-06
(责任编辑:潘学燕Responsibleeditor:PANXueyan)
日期:2017-09-12
网络出版地址:http://kns.cnki.net/kcms/detail/61.1220.S.20170912.1740.032.html
2016-07-12
2016-09-10
北京市教育委员会科学研究与研究生培养共建项目(BLCXY201627);国家林业局948引进项目(2013-04-46)。
阮丽丽,女,在读硕士研究生,研究方向为花卉种质创新与育种。E-mail:lilyruan2014@bjfu.edu.cn
高亦珂,女,教授,主要从事花卉育种与花卉混播等研究工作。E-mail:gaoyk@bjfu.edu.cn
CorrespondingauthorGAO Yike, female, professor.Research area:flower breeding and mixed planting of flowers and plants.E-mail:gaoyk@bjfu.edu.cn
