梭砂贝母种子萌发特性
2017-09-05高永茜肖亚琼
高永茜,肖亚琼
(1.云南林业职业技术学院生态与环境工程学院,云南 昆明 650224;2.西南林业大学林学院,云南 昆明 650224)
梭砂贝母种子萌发特性
高永茜1,2,肖亚琼1
(1.云南林业职业技术学院生态与环境工程学院,云南 昆明 650224;2.西南林业大学林学院,云南 昆明 650224)
对梭砂贝母(Fritillariadelavayi)种子的萌发特性进行研究。结果表明,梭砂贝母种子在浸种6 h后吸水率达到80%以上,不存在吸水障碍;种子浸提液显著抑制塔黄种子萌发,抑制物主要存在于胚乳中;常温层积处理能促进种胚后熟,经过冬季低温层积,次年4月完成胚形态后熟;胚发育完全后仍需低温层积处理才能萌发;15~20 ℃是种子萌发最适宜的温度。梭砂贝母种子休眠主要是由于胚发育不完全和种子中含有萌发抑制物导致,属于混合休眠类型;低温层积处理可有效打破种子休眠。
梭砂贝母;发芽抑制物;吸水率;种子休眠;形态后熟
梭砂贝母(FritillariadelavayiFranch.)为百合科(Liliaceae)贝母属多年生草本植物,主要分布在四川、云南、西藏和青海等省区海拔3800~4700 m的高山流石滩[1]。梭砂贝母与川贝母(F.cirrhosa)、暗紫贝母(F.unibracteata)、甘肃贝母(F.przewalskii)通称川贝母,它们的鳞茎含有多种生物碱、皂苷和多种微量元素等有效成分,具有清热润肺和化痰止咳的功效,是一味常用大宗名贵药材[2]。在经济利益的驱使下,梭砂贝母的野生资源遭到掠夺式的采挖,趋于濒危。不论是从保护野生资源的角度,还是人工规模化种植以满足市场药用需求,均有必要掌握梭砂贝母的繁殖生态学与种群更新机制[3]。自然条件下野生梭砂贝母能通过无性和有性2种方式繁殖。针对贝母属其它植物的研究表明,与繁殖系数低且成本高的鳞茎繁殖相比,播种繁殖具有明显的优势[4-6]。尽管高山地区环境恶劣,熊蜂传粉和较长的花期保证了梭砂贝母在自然条件下具有较高的结实率(87%)[7]。然而,研究川贝母、伊贝母、卷叶贝母、暗紫贝母和甘肃贝母几种植物发现,它们的种子均是具有后熟特性的原胚型种子,必须经过形态后熟和生理后熟2个阶段的后熟作用才具备发芽活力[5-6,8-10]。而且,经后熟作用的甘肃贝母种子萌发率仍然较低,严重限制人工规模化种植[6]。但是,目前尚未见梭砂贝母种子萌发相关研究的报道。本研究旨在弄清楚梭砂贝母种子的萌发特性,为其人工驯化与规模化栽培以及野生资源的保护提供参考。
1 材料与方法
1.1 试验材料
供试梭砂贝母种子于每年8月下旬—9月中旬采自云南省香格里拉县格咱乡翁水村康特矿山(28°24′N、99°55′E,海拔4415 m)。
1.2 种子形态特征观察
取成熟的梭砂贝母种子50粒,在体视镜(Olympus SZX 16,Olympus Corporation,Tokyo,Japan,下同)下观察种子形态,测定种子大小,包括长、宽、高。随机选取净种子1000粒称重,重复5次,计算种子千粒重。
1.3 种子吸水规律测定
随机挑选饱满度均匀的梭砂贝母种子600粒,分为3组(每组200粒),称取干质量后,装入纱布中,室温浸种,每次取出后(浸种1~12 h内每隔1 h取出,之后每隔12 h取出)用滤纸吸干种子表面的水分,称重,计算种子吸水率,连续测定17次。种子吸水率(%)=[(吸水后种子质量-风干种子质量)/风干种子质量]×100。
1.4 发芽抑制物检测
参考王有生等[11]方法,研钵中加少量石英砂及水匀浆,2~3倍水稀释,加热至70~80 ℃提取10~20 min保存,种子提取部位见表1。以蒸馏水为对照,比较各种提取液对塔黄(Rheumnobile)种子发芽的抑制作用。发芽抑制检测方法:每个培养皿中加浸提液或蒸馏水5 mL,播塔黄种子20粒,置于15 ℃恒温箱内培养,统计塔黄种子的发芽情况,直至连续15 d没有新种子萌发为止。每个处理重复5次。

表1 梭砂贝母种子提取液
1.5 层积处理
1.5.1 层积处理对种子形态后熟的作用 9月于云南省香格里拉县格咱乡翁水村康特矿山(梭砂贝母的自然生长地点)采集成熟的梭砂贝母种子,采用常温层积方法在野外对种子进行沙藏,种子与细沙比例为1∶3,适时浇水,使基质的湿度保持在50%~60%,每隔30 d选取10粒种子,采用分离胚的方法,在体视镜下将胚挑出,测定胚长和胚乳长,计算胚率,以干燥未经处理的种子作为对照,每个处理重复3次。胚率(%)=(胚长/胚乳长)×100。
1.5.2 层积处理对种子萌发的影响 常温层积处理至翌年4月,胚率达到61%以上的种子,即已基本完成形态后熟过程。此后每隔30 d随机挑选20粒种子在15 ℃恒温箱内进行萌发试验,每天观察种子的发芽情况,连续15 d没有新种子萌发时停止观察,计算种子的发芽率。每个处理10次重复。
1.6 温度条件对种子萌发的影响
种子经常温层积处理至第3年2月时能正常萌发。随机挑选该类种子置于恒温箱内进行萌发试验,温度条件设置为5、10、15、20、25 ℃ 5个处理。每天观察种子的发芽情况,连续15 d没有新种子萌发时停止观察,计算种子的发芽率。每个处理20粒种子,10次重复。
1.7 数据分析
采用SPSS 18.0软件对数据进行分析,利用Sigmaplot 10.0软件作图。其中,发芽抑制物对塔黄种子萌发的影响、层积处理对梭砂贝母种子萌发的影响和温度条件对梭砂贝母种子萌发的影响采用单因素方差分析(one-way ANOVA)进行检测,层积处理和对照对梭砂贝母种子形态后熟的影响采用双因素方差分析(two-way ANOVA)进行检测。
2 结果与分析
2.1 种子形态
种子呈倒三角状卵形,外种皮棕黄色,表面有不规则褶皱,边缘有膜质翼。种子的千粒重为(7.24±0.21)g,长为(7.94±0.80) mm,宽为(5.80±0.91) mm,厚为(0.46±0.05) mm。胚乳丰富,胚较小,约0.1~0.2 mm。
2.2 种子吸水率
梭砂贝母种子的吸水率随浸种时间的变化格局呈近似“S”型曲线,可以明显分为3个阶段:急剧吸水期、缓慢吸水期和饱和吸水期(图1)。梭砂贝母的种子在浸种前6 h内,吸水率呈显著上升的趋势(急剧吸水期);而在吸水6~10 h内,种子吸水率呈现缓慢上升的趋势(缓慢吸水期);到48 h时,吸水率最大,达到饱和(饱和吸水期)。说明梭砂贝母种子不存在透水障碍。
2.3 发芽抑制物的生物鉴定
梭砂贝母种子的不同浸提液对塔黄种子萌发的抑制作用差异极显著(F(5,24)=69.09,P<0.001)。由图2可知,与蒸馏水对照(处理6)相比,种皮(处理2)和胚(处理4)的浸提液显著抑制塔黄种子的萌发;而胚乳的浸提液(处理3)对塔黄种子萌发的抑制作用显著强于种皮和胚,但是与整粒种子的浸提液(处理1)没有明显的差异,说明发芽抑制物主要存在于胚乳中。已经发芽的种子浸提液(处理5)对塔黄种子的萌发没有显著的抑制作用,说明其中的萌发抑制物已基本消除或产生的萌发促进物质抵消其效应[11]。
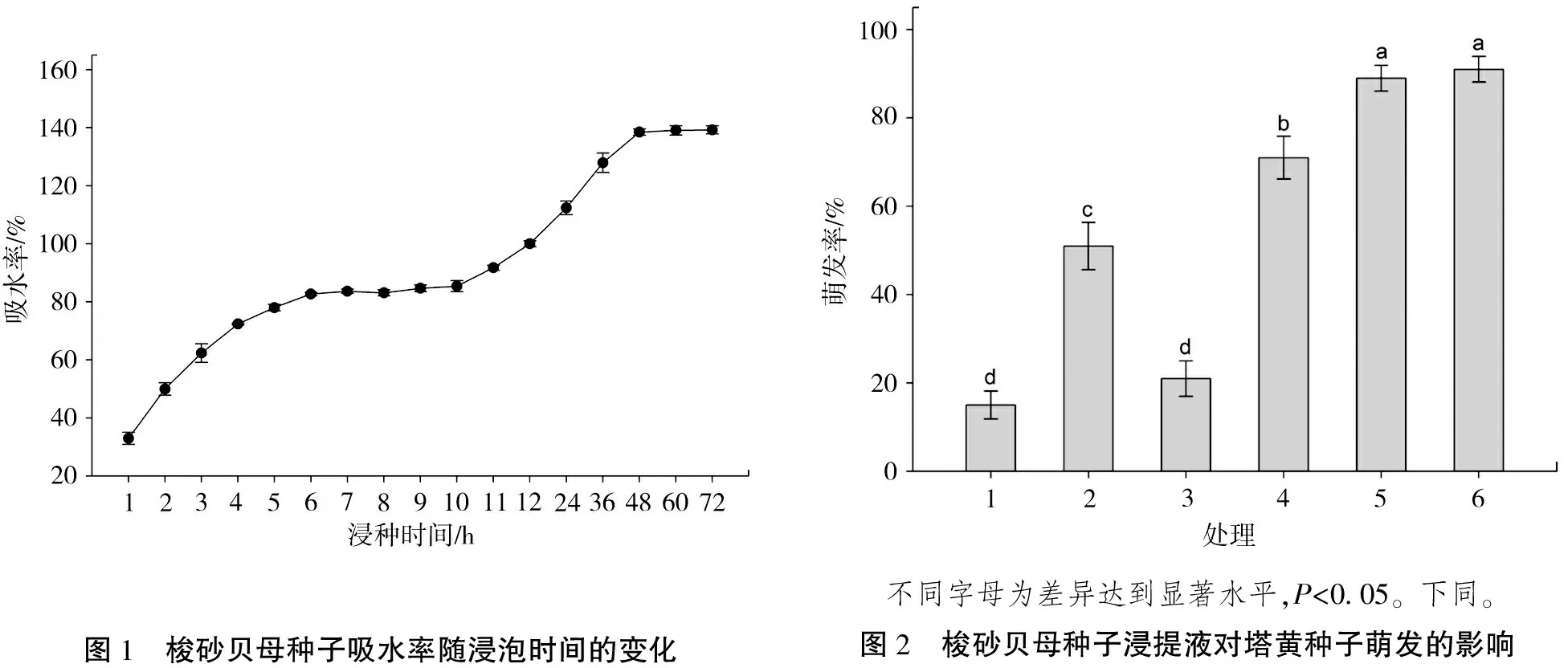
图1 梭砂贝母种子吸水率随浸泡时间的变化 不同字母为差异达到显著水平,P<0 05。下同。图2 梭砂贝母种子浸提液对塔黄种子萌发的影响
2.4 层积处理效果
2.4.1 层积处理对种子形态后熟的作用 梭砂贝母种子在成熟采收时,其胚尚未发育成熟,处于刚分化的时期,体积很小,胚率仅为2.8%。由图3可知,与未层积相比,常温层积处理对胚率的影响达极显著差异(F(1,144)=3522.93,P<0.001),而随着层积时间的延长,胚率显著升高,胚发育逐渐完成(F(7,144)=119.17,P<0.001)。在自然条件下,从11月开始层积,至翌年4月时胚率达到61%,之后胚率没有显著的变化,说明种胚形态后熟基本完成。
2.4.2 层积处理对种子萌发的影响 在常温层积的条件下,从形态后熟完成(翌年4月)开始至翌年10月,梭砂贝母的种子均不能萌发(图4)。层积至翌年12月时种子开始萌发(21.5%),且萌发率随层积时间显著上升,至第3年2月时最高达91.5%(F(11,108)=718.78,P<0.001),说明种子在形态后熟完成以后还需要一个低温处理才能完全打破休眠。
2.5 温度条件对种子萌发的影响
温度对梭砂贝母种子萌发具有显著的影响(F(4,20)=143.26,P<0.001)。梭砂贝母种子在5 ℃时不能萌发,随着温度的升高,萌发率显著上升,15、20 ℃时萌发率达到最大,温度进一步升高萌发率则显著降低(图5),说明梭砂贝母种子萌发的最适温度为15~20 ℃,过高或过低的温度都不利于种子萌发。
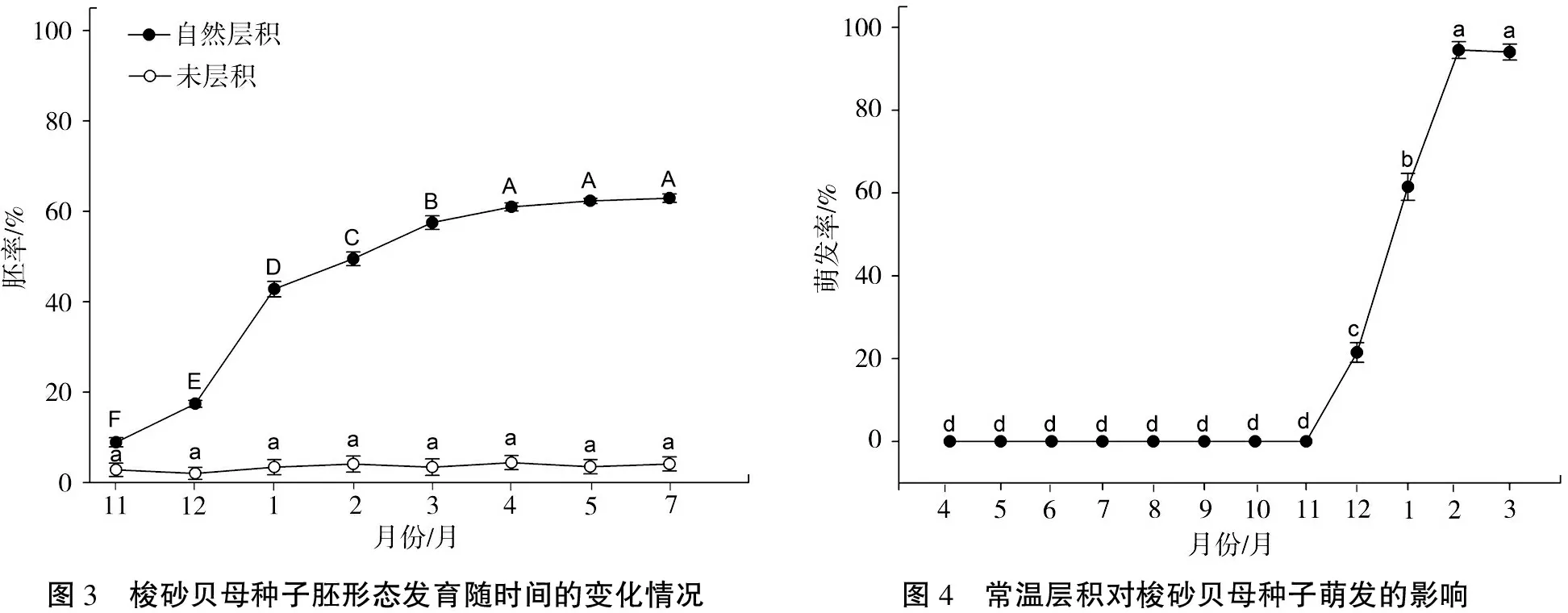
图3 梭砂贝母种子胚形态发育随时间的变化情况图4 常温层积对梭砂贝母种子萌发的影响
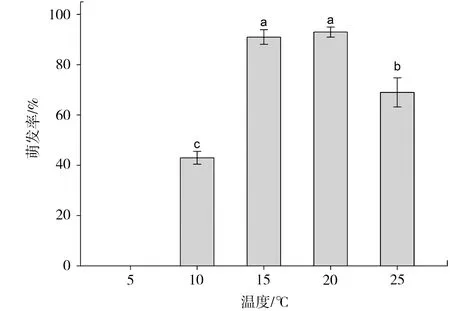
图5 温度对梭砂贝母种子萌发的影响
3 结论与讨论
本研究说明,梭砂贝母种子存在休眠现象,主要是由于胚发育不全和种子中含有萌发抑制物质导致,属于综合休眠类型。在自然条件下,经过2个冬季的低温层积处理能有效打破其休眠。种子休眠打破以后,15~20 ℃是最适宜梭砂贝母种子萌发的温度条件。
梭砂贝母种子与其它贝母属植物种子一样,存在休眠机制。一些植物种子的种皮上含有丰富的脂类物质和油类物质,阻止水分进入或不透气,从而导致种子发芽的环境条件难以满足而休眠[13]。梭砂贝母种子在浸种6 h以后吸水率达到80%以上,48 h后吸水饱和,最大吸水率为139.24%,说明梭砂贝母的种子休眠并不是由于种皮不透水导致[5-6]。尽管本研究中使用的种子是在蒴果已充分成熟时采收,种子仍然没有完整的胚结构(胚率仅为2.8%)。这可能是由于梭砂贝母主要生长在高山冰缘带环境,生长期短,没有足够的时间供胚生长发育[7,13]。将梭砂贝母种子置于原生境条件,从11月开始层积,经冬季低温处理,至翌年4月时胚率达到最大,胚分化完成,说明低温刺激在种子形态后熟过程中起到了关键作用[5,14]。与川贝母等植物一样,经低温层积处理形态后熟完成的梭砂贝母种子仍然不能萌发,还需要低温层积至翌年12月才开始萌动,至第3年2月时萌发率达到最高,说明梭砂贝母种子在胚完成发育以后,仍需要进一步低温处理才能完全打破休眠。马生军等[10]研究伊贝母发现,种子内含有抑制种子萌发的物质。本研究也发现梭砂贝母种子浸提液对塔黄的种子萌发具有显著的抑制作用,而且萌发抑制物质主要存在于胚乳中。因此,梭砂贝母的种子休眠主要是由胚发育不完全和种子中含有萌发抑制物导致,其种子属于综合休眠类型。
种子在萌发过程中新陈代谢十分活跃,而新陈代谢活动对温度非常敏感,因此,大量研究表明温度是影响种子萌发的重要因素[5,15]。每一个物种的种子萌发均有一个最适温度范围,温度过高或过低均不利于种子的萌发。例如,川贝母种子最适萌发温度为25 ℃[5]。本研究表明,温度对梭砂贝母种子萌发也具有显著的影响:梭砂贝母的种子在10~25 ℃范围内均可以萌发,但是,在15~20 ℃时萌发率最高,达到94%。梭砂贝母的种子萌发最适宜温度明显低于川贝母,这可能是由于梭砂贝母分布的海拔比川贝母高,是对所处环境条件适应性进化的结果。
[1]欧珠朗杰.藏药材——梭砂贝母组织培养研究[J].西藏科技,2012(9):72.
[2]杨曦亮,张勇慧,阮汉利,等.中药川贝的本草考证[J].亚太传统医药,2006(7):69-72.
[3]Gulias J,Traveset A,Riera N,et al.Critical stages in the recruitment process of Rhamnus alaternus L[J].Ann Bot,2004,93(6):723-731.
[4]徐世安.“安徽贝母”野生改家种第二报——解除种子休眠与促进萌发的试验研究[J].安徽中医学院学报,1989,8(3):61-62.
[5]于婧,魏建和,陈士林,等.川贝母种子休眠及萌发特性的研究[J].中草药,2008,39(7):1081-1084.
[6]常彦莉,陈垣,郭凤霞,等.甘肃贝种子吸水及发芽特性研究[J].草业学报,2010,19(4):41-46.
[7]高永茜,张立轩,汪明润,等.梭砂贝母的传粉生物学研究[J].中国中药杂志,2014,39(10):1795-1798.
[8]刘潮,饶富民,王曙,等.暗紫贝母人工繁育中应注意的技术问题[J].四川农业科技,2006(7):35.
[9]王跃华,代勇,胡胜玲,等.卷叶贝母种子萌发和多倍体诱导研究[J].中药材,2010,33(8):1201-1204.
[10]马生军,沙红,包晓伟,等.伊贝母种子发芽率影响因素的研究[J].时珍国医国药,2011,22(6):1481-1482.
[11]王有生,王文杰.伊贝母种子的休眠特性及化学药剂对解除休眠的效应[J].中药材,1989,12(4):5-8.
[12]唐安军,龙春林,刀志灵.种子休眠机理研究概述[J].云南植物研究,2004,26(3):241-251.
[13]张维经,胡正海,宇文强.伊贝母(FritillariapallidifloraSchrenk.)种子休眠特性的研究[J].植物学报,1978,20(2):175-177.
[14]张耀甲,王红梅,周瑞莲.甘肃马啣山地区榆中贝母(Fritillariayuzhongensis)的生态学特性及种子发芽规律的初步研究[J].生态学报,1992,12(2):155-159.
[15]宋兆伟,郝丽珍,黄振英,等.光照和温度对沙芥和斧翅沙芥植物种子萌发的影响[J].生态学报,2010,30(10):2562-2568.
Seed Germination ofFritillariadelavayi
GAO Yongqian1,2,XIAO Yaqiong1
(1.DepartmentofEcologicalandEnvironmentalEngineering,YunnanForestryTechnologicalCollege,Kunming650224,Yunnan,China; 2.FacultyofForestry,SouthwestForestryUniversity,Kunming650224,Yunnan,China)
In this study,the seed germination characteristics ofFritillariadelavayiFranch were investigated.The rate of water absorption was more than 80% at 6 h after submerging in water.Seed extracts significantly inhibited the seed germination ofRheumnobileand germination inhibitors were mainly found in endosperm.Natural cold stratification could promote the growth of embryo.Seed did not germinate immediately after embryo growth was completed,and an additional cold stratification period was required to break dormancy completely.Once dormancy was broken,seeds germinated at a range of temperatures,but seed germination rate was the highest at 15~20 ℃.These results indicate that seeds ofF.delavayihave a low-temperature requirement for embryo growth and an additional cold stratification period after completion of embryo growth was needed,exhibiting characteristics of complex morphophysiological dormancy.
Fritillariadelavayi;germination inhibitors;rate of water absorption;seed dormancy;embryo growth
2016-10-20;
2017-02-20
云南省教育厅科学研究基金资助性项目(2016ZZX255);云南林业职业技术学院科学研究项目(KY201503)
高永茜(1983—),女,云南昭通人,云南林业职业技术学院讲师,硕士,从事森林培育与植物资源方面的教学与研究工作。E-mail:gaoqian1028@163.com。
10.13428/j.cnki.fjlk.2017.02.015
S567.23+1
A
1002-7351(2017)02-0077-05
