马尾松自由授粉家系产脂力遗传变异及选择
2017-09-05苏顺德黄德龙魏永平甘振栋林华章郑双全郑仁华黄秀美赵世荣
苏顺德,黄德龙,魏永平,甘振栋,林华章,郑双全,肖 晖,郑仁华,黄秀美,赵世荣
(1.福建省林业科学研究院、国家林业局南方山地用材林培育重点实验室、福建省森林培育与林产品加工利用重点实验室,福建 福州 350012;2.福建省大田桃源国有林场,福建 大田 366101; 3.福建省永安国有林场,福建 永安 366031;4.福建省邵武卫闽国有林场,福建 邵武 354006; 5.福建省沙县官庄国有林场,福建 沙县 365500)
马尾松自由授粉家系产脂力遗传变异及选择
苏顺德1,黄德龙2,魏永平3,甘振栋4,林华章2,郑双全5,肖 晖1,郑仁华1,黄秀美3,赵世荣4
(1.福建省林业科学研究院、国家林业局南方山地用材林培育重点实验室、福建省森林培育与林产品加工利用重点实验室,福建 福州 350012;2.福建省大田桃源国有林场,福建 大田 366101; 3.福建省永安国有林场,福建 永安 366031;4.福建省邵武卫闽国有林场,福建 邵武 354006; 5.福建省沙县官庄国有林场,福建 沙县 365500)
在福建省大田桃源国有林场,分析了马尾松高产脂初级种子园自由授粉子代18 a林龄时产脂力和生长性状的遗传变异规律和遗传相关性。结果表明:产脂力家系遗传力和单株遗传力分别为0.5891和0.6078;树高、胸径和单株立木材积家系遗传力介于0.2999~0.3817间,单株遗传力介于0.2033~0.2865间;产脂力与树高、胸径和单株立木材积呈高度正向遗传相关,遗传相关系数分别为0.761、0.884和0.930。对产脂力进行选择时,树高、胸径和单株立木材积的间接选择响应分别达到0.945、1.209和1.303,在获得产脂力较高遗传增益的同时,可间接获得较大生长性状遗传增益。在遗传变异及遗传相关分析基础上,筛选出高产脂家系8个、单株10株,入选的家系平均产脂力、单株立木材积、胸径和树高遗传增益分别为12.32%、4.19%、2.06%和1.07%。入选的单株平均产脂力、单株立木材积、胸径和树高遗传增益分别为58.05%、18.02%、8.38%和3.14%。
马尾松;自由授粉子代;产脂力;遗传力;育种值
松脂广泛应用于医药、造纸、橡胶合成、胶黏剂等产业,其工业功效与石油树脂类似,但具有更好的相容性,具有可再生性[1-2]。松脂的广泛应用更利于环境保护和相关产业可持续发展。我国松脂产量占全世界松脂总产量的40%以上,松香、松节油及其深加工产品也是我国林业产业的支柱产品[3-5]。我国主产松脂的乡土树种主要为马尾松(Pinusmassoniana)、云南松(P.yunnanensis)、思茅松(P.kesiyavar.langbianensis)、南亚松(P.latteri),其中马尾松的面积和蓄积分别占可采脂松树面积和蓄积的77.4%和62.0%[4],松脂产量占全国松脂总产量的90%[5]。高产松脂遗传改良是显著提高马尾松脂生产量最有效的途径之一,高产脂良种选育是维持我国松脂产业优势的关键[6-7]。因此,马尾松产脂力遗传改良一直是我国松脂遗传改良的重要内容。郑元英等[8]分析了马尾松高产脂种子园自由授粉子代4~5 a林龄时的产脂力,发现产脂型马尾松子代产脂力显著高于普通马尾松,认为开展高产脂家系选育,有利于提高马尾松脂产量和质量。覃冀等[9]测定了马尾松优树自由授粉子代产脂力,筛选出16个高产脂优良家系。曾令海等[10]测定了马尾松杂交子代6 a林龄时产脂量的杂种优势,认为杂交组合可从母本获得较高的增益,最优杂交组合产脂量杂种优势可达155.7%,证明了开展自由授粉子代测定,筛选一般配合力高的育种材料是可行的。张谦等[11]分析了高产脂马尾松优树半同胞子代产脂力的年度变化规律,发现11 a林龄是马尾松产脂力早期选择的最佳时间节点,产脂力早晚期遗传相关随着林龄增大而增强。鉴于马尾松为我国重要的用材树种[12],本研究利用18 a林龄马尾松产脂型初级种子园自由授粉子代开展产脂力和生长综合选择,以期为产脂型速生优良育种材料选择提供参考。
1 试验地概况
试验地位于福建省大田桃源国有林场,地理位置为117°31′E、25°48′N。属中亚热带湿润季风气候,雨热同期,夏湿冬干,干湿季节明显。年均温18.3 ℃,年均降水量1680 mm,平均相对湿度80%,年均日照时间1723.8 h,海拔400~450 m,坡度15°~20°,土壤为山地红壤,立地类型为Ⅱ-Ⅲ类地。造林地为杉木人工纯林采伐迹地。
2 材料与方法
2.1 试验材料
参试材料为福建省国有来舟林业试验场1986年嫁接营建的马尾松高产脂初级种子园自由授粉子代,参试家系24个,以大田梅山种源为对照,共计25个处理。1996年春造林。
2.2 试验方法
田间试验设计为完全随机区组设计(RCB),4株双列小区,8次重复,小区内株行距2.0 m×2.0 m,小区间的株行距为3.0 m×3.0 m。2013年底每木调查树高、胸径。根据生长调查数据,2014年每个小区中选取最优2株测定松脂产量,林龄18 a。
采脂方法为下降式采脂,割面设置在树干向阳、枝叶茂盛和便于采割的一侧,割面上缘距离地面高度2.2 m。割面的宽度不超过树围长的40%,割面长度20 cm。割面标定好后,用刮刀刮去树干粗皮至无裂纹快近韧皮部、不流脂即可,残留粗皮厚度不超过0.4 cm。然后在刮面正中开中沟,长约30 cm,宽约1.5 cm,沟槽外宽内窄,笔直而光滑,中沟下部放置受脂容器采集树脂。第1对侧沟开在中沟顶端,2条侧沟等长,侧沟与中沟的夹角为45°,沟深约0.3 cm,宽度约0.2 cm。第2对侧沟开在第1对侧沟的下方,依次从上往下开侧沟,所有侧沟等长、等深、平行。每天开1对侧沟,每15 d收脂1次,并称量松脂重量。
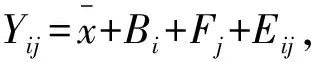
3 结果与分析
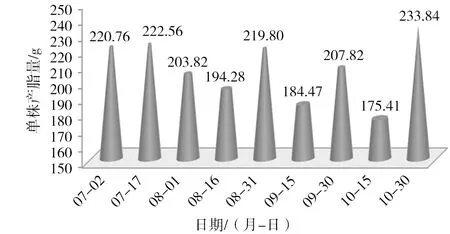
图1 2014年每15 d单株平均产脂量
3.1 产脂量年变化分析
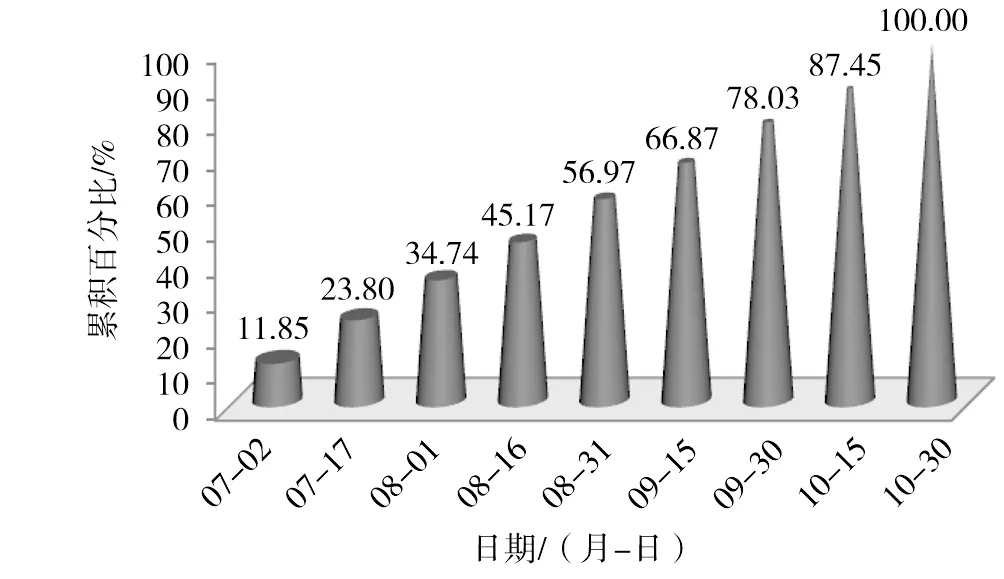
图2 2014年每15 d单株产脂量累积百分比
图1为试验林2014年7—10月每隔15 d测定的单株平均松脂产量锥形图。6月下旬刚开始采脂时,松脂产量较高,持续至7月上旬,每15 d平均单株产量达220 g以上。此后每15 d单株平均松脂产量呈现高、低交替变化规律。测定时期内(6月下旬—10月底),单株平均松脂产量为1862.77 g(各测定值之和),从6月下旬的产量看,每15 d松脂产量均处于峰值,据松脂产量和气象因子的相关性[18-19],结合福建省气候特点,福建省马尾松脂采集起始时间可提前至5月下旬。图2溪每15 d单株平均松脂产量占全年单株产脂量的累计百分比锥形图。由图2可知,到8月底时,单株松脂产量已达全年单株产量的56.97%,也即9月、10月产量占43.03%。同时,7—10月每个月单株松脂产量贡献率介于21%~23%间(如7月贡献率为34.74%~11.85%=22.89%),单株平均松脂产量在采脂期内的月份上分布较均匀,10月还未表现出明显下降趋势,表明福建省马尾松采脂期终止时间可延长。据松脂产量和气象因子的相关性[5,18],结合福建省气候特点,福建省马尾松脂采集终止时间可延长至11月上旬。
3.2 产脂力及生长性状遗传变异分析
以家系均值分析参试家系平均产脂力及生长性状表现(表1)。由表1可知,家系平均产脂力、树高、胸径和单株立木材积分别为77.69 g·cm-1、14.94 m、17.5 cm和0.18111 m3,表型变异系数分别为17.06%、3.22%、7.50%和15.46%,产脂力和生长性状在家系间表型变异丰富,为高产脂速生家系选择奠定了基础。
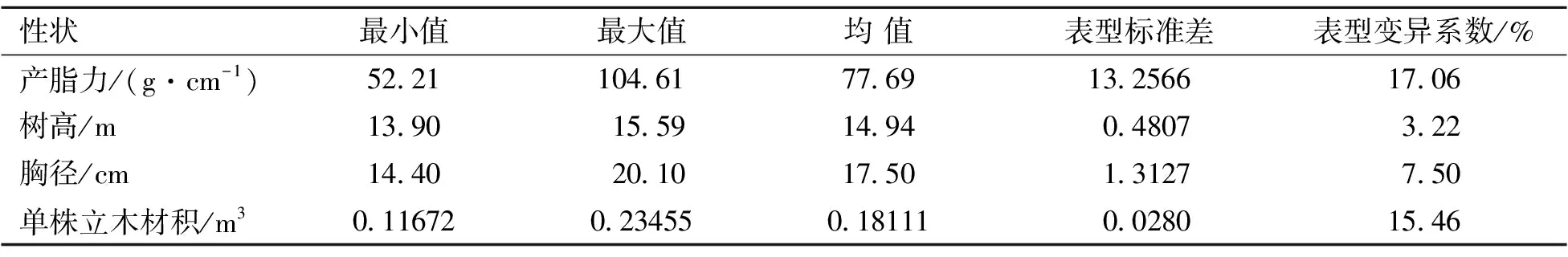
表1 产脂力及生长平均表现
对生长性状以单株为单位进行方差分析表明,重复与家系的交互作用不显著,家系间差异极显著。因此以小区平均值为单位估算产脂力和生长性状方差分量,并估算家系遗传力和单株遗传力,结果见表2。产脂力和生长性状在家系水平上差异均达极显著水平。产脂力受中度遗传控制,家系遗传力和单株遗传力分别为0.5891和0.6078,与产脂型初级种子园建园优树自由授粉子代12 a林龄产脂力的遗传力较为接近[19]。树高、胸径和单株立木材积受中偏弱遗传控制,家系遗传力介于0.2999~0.3817间,单株遗传力介于0.2033~0.2865间。以上结果表明从参试材料中筛选优良家系,可以获得较高的产脂力遗传增益和中等的生长性状遗传增益,为产脂型家系、速生型家系选择提供了可能。
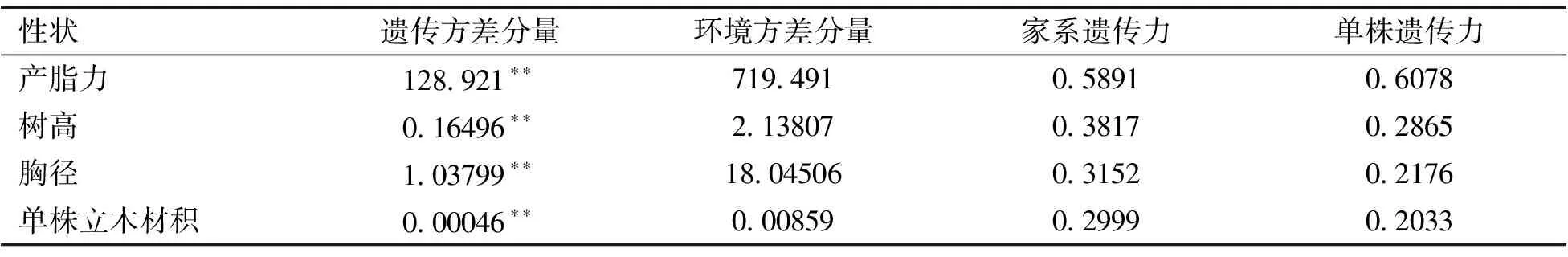
表2 产脂力及生长性状方差分析及遗传参数估算
*:*为在0.05水平上差异显著,**为在0.01水平上差异显著。下同。
3.3 产脂力与生长性状相关性及间接选择响应
研究性状间的相关性对家系选择具有重要参考价值。产脂性状与生长性状的表型相关和遗传相关系数见表3。产脂力与树高、胸径和单株立木材积的表型相关系数分别为0.274、0.473和0.438,呈中度正相关,遗传相关系数分别为0.761、0.884和0.930,呈高度正向遗传相关。对产脂力进行选择的同时也间接对生长性状进行了正向选择。对产脂力进行选择时生长性状的间接选择响应,以及对生长性状进行选择时,产脂力的间接选择响应见表4。由表4可知,当对产脂力进行选择时,单株立木材积的间接选择响应最大,为1.303;但对生长性状进行选择时,产脂力的间接选择响应介于0.613~0.664间。可见,性状间高度遗传相关时,可对遗传力较高的性状进行直接选择,一方面可获得直接选择性状较高遗传增益,另一方面可对遗传较低的性状进行较高效率的间接选择,获得的遗传增益甚至超过对其本身的直接选择。本研究中对产脂力进行选择时,对胸径和单株立木材积的选择效果均大于它们本身的直接选择。
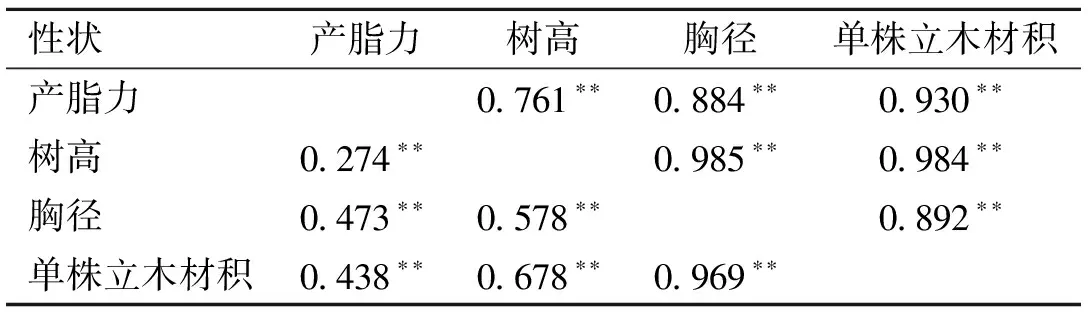
表3 性状间相关系数
*:表格右上部分为遗传相关系数,左下部分为表型相关系数。
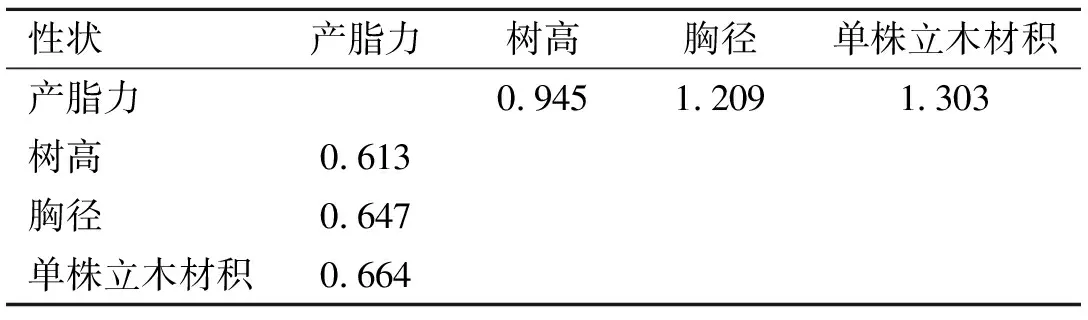
表4 产脂力与生长性状的间接选择相应
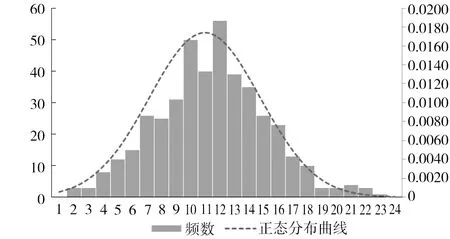
图3 单株产脂力遗传增益分布图
3.4 高产脂家系及单株选择
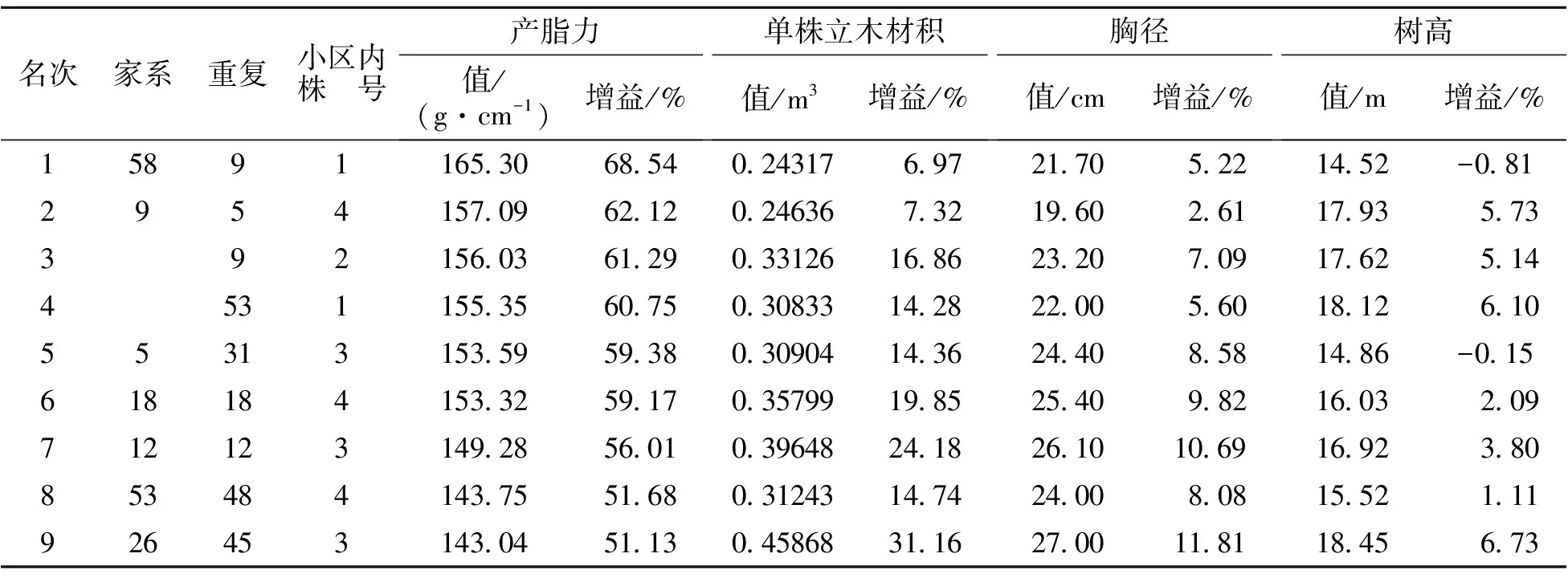
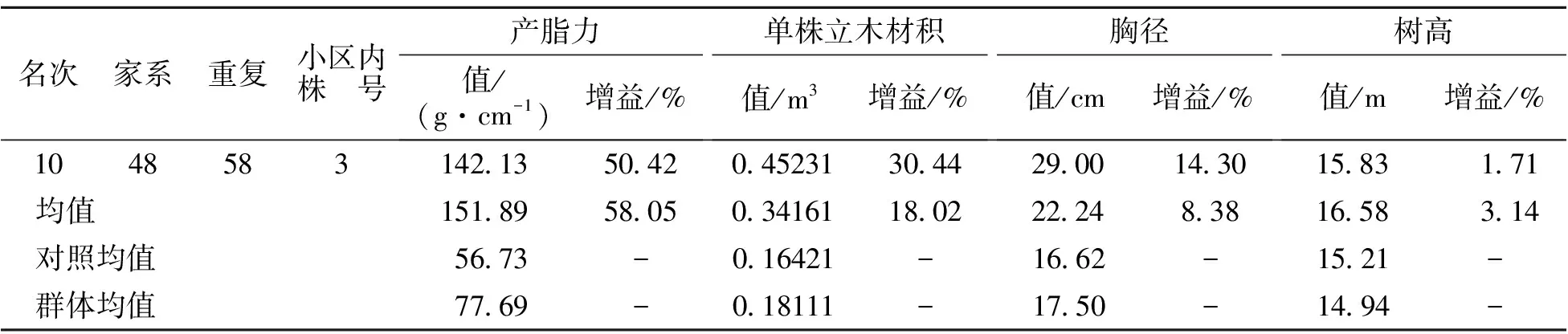
上述分析表明对产脂力进行选择的同时,对生长性状也起到了良好的间接选择。因此,以家系产脂力育种值为依据,30%入选率[20]筛选高产脂家系8个,结果见表5。入选家系平均产脂力育种值为11.906。平均产脂力、单株立木材积、胸径和树高分别为93.94 g·cm-1、0.20639 m3、18.64 cm和15.36 m,分别比大田梅山种源(对照)大65.59%、25.69%、12.15%和0.99%。以参试群体均值为参照,遗传增益分别为12.32%、4.19%、2.06%和1.07%。筛选的家系可作为马尾松高产脂种子园建园材料,也可作为现有初级产脂种子园去劣疏伐的依据。为了最大限度地利用基因加性效应和非加性效应,在试验林中选择优良单株(基因型)作为无性扩繁材料是林木良种繁育的重要途径。
本研究中测定产脂力的单株均为每个小区中的最优2株,因此以单株产脂力遗传增益为依据筛选优良单株。单株产脂力遗传增益频数分布图见图3,图中虚线为拟合正态分布曲线。由图3可知,单株产脂力遗传增益在参试单株中基本呈正态分布,根据正态分布3σ原则:P(μ-σ 表5 高产脂优良家系 表6 高产脂优良单株 表6(续) 我国松脂产量在世界中占比较高,因此我国在高产脂松树良种选育方面的研究水平明显优于国外[11]。而在高产脂松树选育中,马尾松良种选育为主要内容,近年来浙江、广东和广西等省区在马尾松高产脂良种选育方面的研究较为活跃,福建省趋缓[3,7,10-11]。这主要是由于马尾松产脂力测定对试验林林龄要求较高,加之遗传力在11 a林龄以后才趋于稳定,福建省1990年以后营建的高产脂遗传测定林大多还未进入产脂力测定阶段。不同学者对产脂力的估算方法不同,有的以单刀每10 cm长度割沟产脂量估算[19],有的以单位胸径长度松脂产量估算[9]。不管哪种估算方法,其目的在于对松脂产量进行换算,使其具有可比性。但最为直接的方法是以单位胸径长度进行换算,其以胸径量对松脂产量进行度量,兼顾了生长性状与产脂量间的相关性。本研究以单位胸径长度松脂年产量估算产脂力,采脂时间长,测定工作量大,但其对参试家系的产脂能力的估算较为准确。 本研究分析了马尾松高产脂初级种子园自由授粉子代18 a林龄时产脂力和生长性状的遗传变异规律及其遗传相关性,发现产脂力受中度遗传控制,家系遗传力和单株遗传力分别为0.5891、0.6078;树高、胸径和单株立木材积受中偏弱遗传控制,家系遗传力介于0.2999~0.3817间,单株遗传力介于0.2033~0.2865间;产脂力与树高、胸径和单株立木材积呈高度正向遗传相关,遗传相关系数分别为0.761、0.884、0.930。对具有高遗传力的产脂力进行选择时,生长性状间接选择响应较大,其中胸径和单株立木材积的间接选择响应分别达到1.209、1.303,在获得产脂力较高遗传增益的同时,可间接获得较大的胸径和单株立木材积的遗传增益,这种增益甚至大于对它们的直接选择增益。 在遗传变异及遗传相关性分析基础上,以家系产脂力育种值为依据,30%入选率筛选出高产脂家系8个,入选率家系平均产脂力育种值为11.906,平均产脂力、单株立木材积、胸径和树高分别为93.94 g·cm-1、0.20639 m3、18.64 cm和15.36 m,遗传增益分别为12.32%、4.19%、2.06%和1.07%。同时,以单株产脂力遗传增益为依据,2.3%入选率筛选出优良单株10株,入选单株平均产脂力、单株立木材积、胸径和树高分别为151.89 g·cm-1、0.34161 m3、22.24 cm和16.58 m,遗传增益分别为58.05%、18.02%、8.38%和3.14%。 试验发现参试家系单株平均松脂年产量为1862.77 g,并在年采脂期内的月份上均匀分布,依据松脂产量年变化规律,推断福建省马尾松采脂期为每年5月下旬—11月上旬,时长6个月。 [1]Lee H J,Ravn M M,Coates R M.Synthesis and characterization of abietadiene,levopimaradiene,and neoabietadiene:hydrocarbon precuisors of the abietane diterpene resin acids[J].Tetrahedron,2001,57(29):6155-6177. [2]Manninen A M,Utriainen J,Holopainen T,et a1.Comparing the variation of needle and wood terpenoids in Scots pine provenances[J].Can J For Res,2002,28(1):211-228. [3]董虹妤,刘青华,周志春,等.马尾松3代杂交子代幼林松脂化学组分的GCA/SCA分析[J].林业科学研究,2016,29(5):654-661. [4]董静曦,郭辉军,赵元藩,等.我国的采脂松树资源和松脂贮量——基于“连清”数据的估测[J].林业科学,2009,45(120):112-117. [5]刘云.气象因子对马尾松松脂产量影响的研究[J].中南林业科技大学学报,2011,31(6):65-69. [6]楼浙辉,舒洪岚.松树高产脂力遗传改良的研究进展[J].江西林业科技,2002(5):40-41,44. [7]杨章旗.广西松脂原料林发展现状、存在问题与对策研究[J].广西林业科学,2007,36(3):143-146. [8]郑元英,丁羽,邹高顺,等.高产脂马尾松自由授粉子代林的产脂力测定和松脂组分分析[J].福建林业科技,1990(2):27-34. [9]覃冀,连辉明,曾令海,等.高产脂马尾松半同胞子代20年生测定林产脂力分析[J].广东林业科技,2005,21(2):30-34. [10]曾令海,连辉明,何波祥,等.马尾松杂交组合生长及产脂力早期评价[J].广东林业科技,2006,22(4):1-6. [11]张谦,曾令海,何波祥,等.马尾松自由授粉家系产脂力的年度变化及遗传分析[J].林业科学,2013,49(1):48-52. [12]周政贤.中国马尾松[M].北京:中国林业出版社,2001:1-5. [13]中华人民共和国林业部.LY208—77立木材积表[S].北京:技术标准出版社,1978. [14]Namkoog G..Introduction to quantitative genetics in forestry[M].London:Castla a House pub,1981:210-259. [15]T.L.White,W.T Adams,D B.Neale.Forest Genetics[M].Oxford:CABI Publishing,2007:149-186. [16]Kumar S,Lee J.Age-age correlations and early selection for end-of-rotation wood density in radiate pine[J].Forest Genetics,2002,9(4):323-330. [17]Wu H X,Powell M B,YANG J L.Efficiency of early selection for rotation-aged wood quality traits in radiate pine[J].Annals of Forest science,2007,64(1):1-9. [18]安宁,丁贵杰.主要气象因子对马尾松松脂产量的影响初报[J].山地农业生物学报,2010,29(2):177-180. [19]刘云.气象因子对马尾松松脂产量影响的研究[J].中南林业科技大学学报,2011,31(6):65-69. [20]刘月蓉.高产脂马尾松优树自由授粉子代林产脂力测定[J].福建林学院学报,2005,25(3):229-233. [21]郑仁华,杨宗武,傅玉狮,等.马尾松优良育种资源长期保存、评价和第二代利用研究[R].福州:福建省林业科学研究院,2011. Genetic Variation and Selection of the Resin-Yielding Capacity of Open-Pollinated Families of Masson Pine SU Shunde1,HUANG Delong2,WEI Yongping3,GAN Zhendong4,LIN Huazhang2,ZHENG Shuangquan5,XIAO Hui1,ZHENG Renhua1,HUANG Xiumei3,ZHAO Shirong4 (1.FujianAcademyofForestry,KeyLaboratoryofSouthMountainTimberCultivationofStateForestryAdmioistration,FujianKeyLaboratoryofForestCultivationandForestProductsProcessing,Fuzhou350012,Fujian,China;2.DatianTaoyuanNationalForestFarmofFujian,Datian366101,Fujian,China;3.Yong′anNationalForestFarmofFujian,Yong′an366031,Fujian,China;4.ShaowuWeiminNationalForestFarmofFujian,Shaowu354006,Fujian,China;5.ShaxianGuanzhuangNationalForestFarmofFujian,Shaxian365500,Fujian,China) For studying the genetic variation rule of masson pine (Pinusmassoniana),the resin-yielding capacity and growth traits such as tree height,diameter at breast height (DBH) and volume of 24 families at age of 18 years were examined in an open-pollinated Progeny test on Datian Taoyuan national forest farm,in Fujian Province.The analysis observed significant differences on resin-yielding capacity and growth among tested families.The family and individual heritability of resin-yielding capacity were 0.5891 and 0.6078,and those of growth were between 0.2033 and 0.3817,indicating genetic factor performance moderate control to resin yield but mild control to growth.Furthermore,resin-yielding capacity and growth showed close positive genetic correlation.For instance,the genetic correlation coefficient between resin-yielding capacity and tree height,DBH and volume was 0.761,0.884 and 0.930,respectively.Moreover,the indirect selection response of tree height,DBH and volume to resin-yielding capacity was 0.945,1.209 and 1.303,separately.8 families and 10 individuals were identified as superior families and individuals based on genetic analysis,getting average family genetic gain of 12.32%,4.19%,2.06% and 1.07%,average individual genetic gain of 58.05%,18.02%,8.38% and 3.14% on resin-yielding capacity,volume,DBH and tree height. Pinusmassoniana;open-pollinated progeny;resin-yielding capacity;heritability;genetic correlation;breeding value;selction 2017-03-13 三明市科技项目(马尾松速生及高产脂优良家系选择,明科办[2013]21号);国家科技支撑计划子课题(中带马尾松速生丰产林定向培育技术研究,2015BAD09B010201) 苏顺德(1980—),男,重庆人,福建省林业科学研究院高级工程师,从事林木遗传育种研究。E-mail:ssdforest@163.com。 郑仁华(1970—),男,福建罗源人,福建省林业科学研究院教授级高工,从事林木遗传育种研究。E-mail:zrh08@126.com。 10.13428/j.cnki.fjlk.2017.02.001 S791.248;S722.8 A 1002-7351(2017)02-0001-06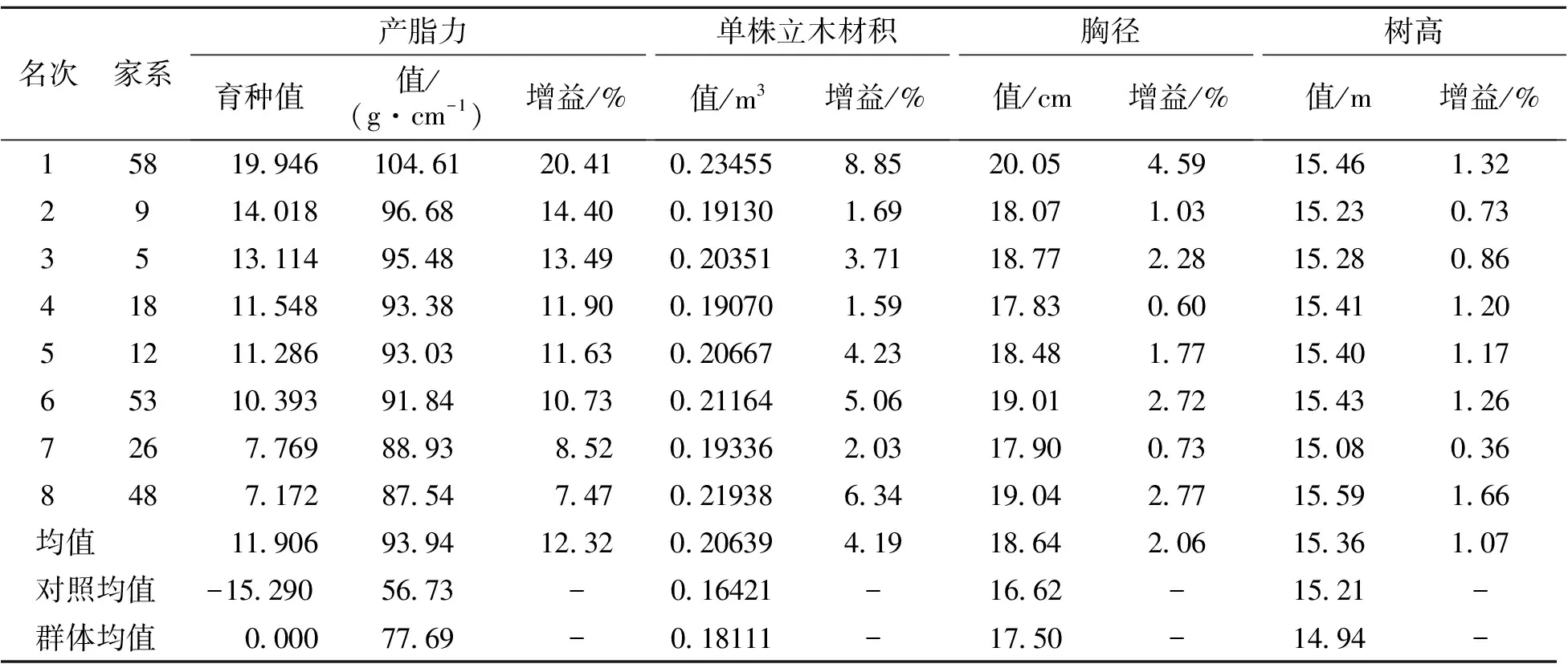
4 结论与讨论
