青稞籽粒阿拉伯木聚糖含量的遗传分析
2017-08-16刘新春巴桑玉珍冯宗云
刘新春,巴桑玉珍,2,冯宗云
(1.四川农业大学农学院作物遗传育种学系大麦青稞研究中心,四川成都 611130;2.西藏自治区农牧科学院农业研究所,西藏拉萨 850032)
青稞籽粒阿拉伯木聚糖含量的遗传分析
刘新春1,巴桑玉珍1,2,冯宗云1
(1.四川农业大学农学院作物遗传育种学系大麦青稞研究中心,四川成都 611130;2.西藏自治区农牧科学院农业研究所,西藏拉萨 850032)
为了解青稞籽粒阿拉伯木聚糖的遗传特性, 以喜马拉雅8号和藏青320及二者构建的F1和F2群体为材料, 使用主基因+多基因模型, 对青稞籽粒总阿拉伯木聚糖含量、水溶性阿拉伯木聚糖含量、不溶性阿拉伯木聚糖含量及不溶性阿拉伯木聚糖/水溶性阿拉伯木聚糖比值进行了遗传分析。结果表明,青稞籽粒总阿拉伯木聚糖含量与不溶性阿拉伯木聚糖含量显著相关, 而与水溶性阿拉伯木聚糖含量相关不显著。籽粒总阿拉伯木聚糖符合1对显性-加性主基因+多基因遗传模式。不溶性与水溶性阿拉伯木聚糖及不溶性与水溶性阿拉伯木聚糖之比均受2对主基因+多基因调控,只是主基因的作用方式存在着差异。控制水溶性阿拉伯木聚糖和不溶性阿拉伯木聚糖含量的主基因加性效应均为负值,可能为负调控因子。除调控水溶性阿拉伯木聚糖含量的主基因加性效应较小外,其余性状的主基因加性效应均较大。各性状的主基因遗传率都较高,为62.5%~88.49%。
青稞;阿拉伯木聚糖;主基因+多基因模型;遗传估计
青稞是裸大麦的一种别称,在世界各个地区均有栽培。青稞籽粒含有多种营养成分和活性物质[1],虽然适口性差,但因为富含可食性纤维和β-葡聚糖,近年来被人们重新认识和发现,并成为保健食品的原料之一。王显萍[2]对我国195份青稞测定发现,其平均β-葡聚糖含量为5.1%,显著高于Tato等[3]对268份日本皮大麦的测定结果(4.26%),也高于我国皮大麦的平均含量(4.84%)[4]。阿拉伯木聚糖是植物细胞壁上两种主要分布的非淀粉性多糖之一,在谷物的非胚乳组织中有较高的含量,在小麦种皮中戊聚糖(阿拉伯木聚糖属于其中的一类)含量达到64%[5]。阿拉伯木聚糖与β-葡聚糖的功能类似,对植物细胞骨架具有一定的支持作用[6]。其对哺乳生物是一种抗营养的拮抗因子。王修启等[7]研究发现,饲料中添加0.12%的木聚糖酶会使公鸡对小麦籽粒干物质的消化率比对照组高7.10%以上。由于阿拉伯木聚糖具有高黏度、高持水力、氧化交联等性质,其在食品加工中扮演多种不同的角色[8]。Michniewicz 等[9]认为,适量添加戊聚糖可以增加小麦面粉吸水率,提高面团形成时间和稳定时间,增强面筋弹性,进而改善面粉品质。Gamlath等[10]通过相关分析得出,大麦籽粒高含量的阿拉伯木聚糖会引发籽粒硬度系数增加,改变其研磨的性能。阿拉伯木聚糖可分为水溶性阿拉伯木聚糖(WEAX)和不溶性阿拉伯木聚糖(WUAX),两者的区别主要是WEAX中阿拉伯糖与木糖的比值及其分子量小于WUAX[11-12]。相关研究发现,大麦汁中过多的WEAX会影响啤酒酿造,导致麦汁粘度增加,使啤酒-酵母凝聚提前,降低啤酒泡沫的稳定性和品质[13-14]。但总体来说,由于阿拉伯木聚糖在谷物籽粒中分布的特点,其研究现状落后于同属非淀粉多糖类的β-葡聚糖。谷物籽粒阿拉伯木聚糖及戊聚糖的含量通常受遗传因素和环境效应影响,而两方面的影响大小通常存在着争议。李春喜等[15]认为,小麦籽粒戊聚糖含量主要受环境因素制约;而张岐军等[16]则认为,基因型和环境是影响戊聚糖的主要因素,同时基因型也是影响水溶性戊聚糖含量的主要因子。现在的相关研究表明,大麦籽粒阿拉伯木聚糖的总含量一般在4%~7%[17-18],且在不同用途的大麦间差异较大[17]。
谷物籽粒阿拉伯木聚糖的含量高低是数量性状,受多基因控制[19]。Yang等[20]研究发现,在PH82-2/Neixiang 188小麦群体中B组和D组染色体上存在着多个调控籽粒阿拉伯木聚糖相关性状的遗传位点。Bordes等[21]和Quraishi等[22]分别利用关联分析在第1、第5和第7同源染色体中发现了调控小麦籽粒阿拉伯木聚糖相关性状的QTL。主基因+多基因混合遗传模型是盖均镒、章元明发展的一种研究数量性状遗传规律的分析方法[23-24]。运用该方法对大麦相关遗传性状的分析有一定报道。吕亮杰等[25]发现,在大麦品种Noso Nijo与泰兴9425形成的DH群体中,籽粒直、支链淀粉的含量受2对主基因和多个微效基因控制,总淀粉含量则受3对主基因和多个微效基因控制。杨晓梦等[26]对S500和宽颖大麦的BC3F1和BC3F2后代籽粒的4种功能成分考察后认为,大麦籽粒的总黄酮、γ-氨基丁酸和抗性淀粉含量均受2对主基因和多基因调控,而生物碱则主要以1对主基因控制为主。目前尚未见有运用该方法研究青稞籽粒阿拉伯木聚糖相关性状的报道。本研究以藏青320和喜马拉雅8号及二者杂交形成的F1、F2代为材料,对青稞籽粒阿拉伯木聚糖相关性状进行遗传模型解析,以期了解青稞籽粒阿拉伯木聚糖的遗传特性,为多用途和专用型优质青稞新品种的培育提供一定的理论指导。
1 材料与方法
1.1 材 料
本试验所用的材料为喜马拉雅8号(P1)和藏青320(P2)及其杂交形成的后代群体。两个亲本于2013年6月在西藏拉萨进行杂交形成F1子代,部分F1群体籽粒于2013年10月种植于农业部现代农业产业体系成都大麦试验站崇州基地,2014年5月收获F2单株籽粒。喜马拉雅8号为西藏日喀则农科院培育优质抗寒性好的青稞品种;藏青320为西藏农牧科学院以藏青334与当地白青稞杂交形成的F6品系7327 再和藏青7239 杂交选育的优质广适性青稞品种[27]。经本课题组前期研究发现,两亲本的籽粒阿拉伯木聚糖含量存在着差异。
1.2 试验方法
1.2.1 田间试验设计
试验于2014年9月在崇州基地将喜马拉雅8号、藏青320、F1和F2进行单粒点播。行长1.5 m,行距0.5 m,株距0.3 m。正常田间水肥管理。于2015年5月选取 P1、P2各10株,F1群体14株及F2群体 121株,单株收获,籽粒烘干磨粉备用。
1.2.2 青稞籽粒阿拉伯木聚糖相关性状测定
参考陈锋等[28]和张晓勤等[29]报道的方法提取青稞籽粒总阿拉伯木聚糖和水溶性阿拉伯木聚糖,并做适当修改。
总阿拉伯木聚糖(TAX)测定:取0.1 g 样本粉末置于15 mL 离心管,加入4 mL 1 mol·L-1H2SO4并混匀,放入沸水中抽提10 min后,冷却至室温。6 000 r·min-1离心5 min,取1 mL上清液至于15 mL离心管,加入等体积的H2O,再加入10 mL反应液(110 mL冰乙酸,2 mL 浓HCl,1 mL 1.75 g·mL-1葡萄糖,5 mL 10%间苯三酚-乙醇溶液)并混匀,置于沸水中反应25 min后,迅速冷却至室温,终止反应,采用双波长法(552和510 nm)测定和计算反应液吸光度之差,根据木糖标准曲线计算样品中总阿拉伯糖木聚糖含量。
木糖标准曲线绘制:准确移取0.1 g·mL-1木糖标准溶液0、0.5、1.0、1.5、2.0 mL,分别置于15 mL离心管,补加水至2.0 mL,再加入10 mL反应液,混合均匀。其他步骤同前。由木糖浓度△A(552-510)做木糖标准曲线。
木聚糖含量=c×0.88×n/105×100%
式中c表示由木糖标准曲线得到的木糖值;0.88 为木糖与木聚糖的转换系数;n为稀释倍数。
水溶性阿拉伯木聚糖(WEAX)和不溶性阿拉伯木聚糖 (WUAX) 含量的测定:取0.1 g 样本粉末置于15 mL 离心管,加入10 mL双蒸水置于反复振荡器中,在室温条件下抽提30 min后。6 000 r·min-1离心5 min,取1 mL上清液进行WEAX含量的测定,后续步骤与计算方法同籽粒总阿拉伯糖提取与计算方法。WUAX=TAX-WEAX。
1.3 数据处理与分析
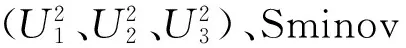
2 结果与分析
2.1 青稞籽粒阿拉伯木聚糖及相关性状统计分析
测定结果(表1和图1)表明,喜马拉雅8号和藏青320的籽粒WEAX平均含量分别为0.332%和0.625%,F1和F2群体的籽粒WEAX平均含量均介于两亲本之间,分别为0.491%和0.596%,但F2群体的平均值偏向于高值亲本藏青320。F2单株的WEAX含量表现为近连续分布,有42.97%的单株的WEAX含量高于高值亲本,表明WEAX在F2群体中具有一定的超亲分布,其表达可能为多基因控制。
籽粒TAX、WUAX含量的各世代变异趋势与WEAX的变异情况相似。喜马拉雅8号和藏青320的籽粒TAX平均含量分别为3.445%和5.328%,F1和F2群体的籽粒TAX含量均位于两亲本之间,分别为4.434%和5.245%, F2群体的平均值偏向于高值亲本藏青320。喜马拉雅8号和藏青320的籽粒WUAX平均含量分别为2.972%和5.114%,F1和F2群体的籽粒WUAX平均含量分别为3.907%和4.649%,均介于两亲本之间,F2群体的平均值也偏向于高值亲本藏青320。籽粒TAX、WUAX含量高于藏青320的F2单株数分别占总调查株数的50.41%、37.19%。
不溶/水溶性阿拉伯木聚糖比(WUAX/WEAX)却呈现不同的趋势。喜马拉雅8号和藏青320的WUAX/WEAX平均比值分别为8.945和8.118,F1和F2群体的平均值均小于低值亲本。

表1 P1、P2、F1和F2多世代青稞群体籽粒阿拉伯木聚糖相关性状的表型分布Table 1 Phenotypic distribution of grain arabinoxylan and related traits in hulless barley from P1,P2,F1 and F2 multi-generation population
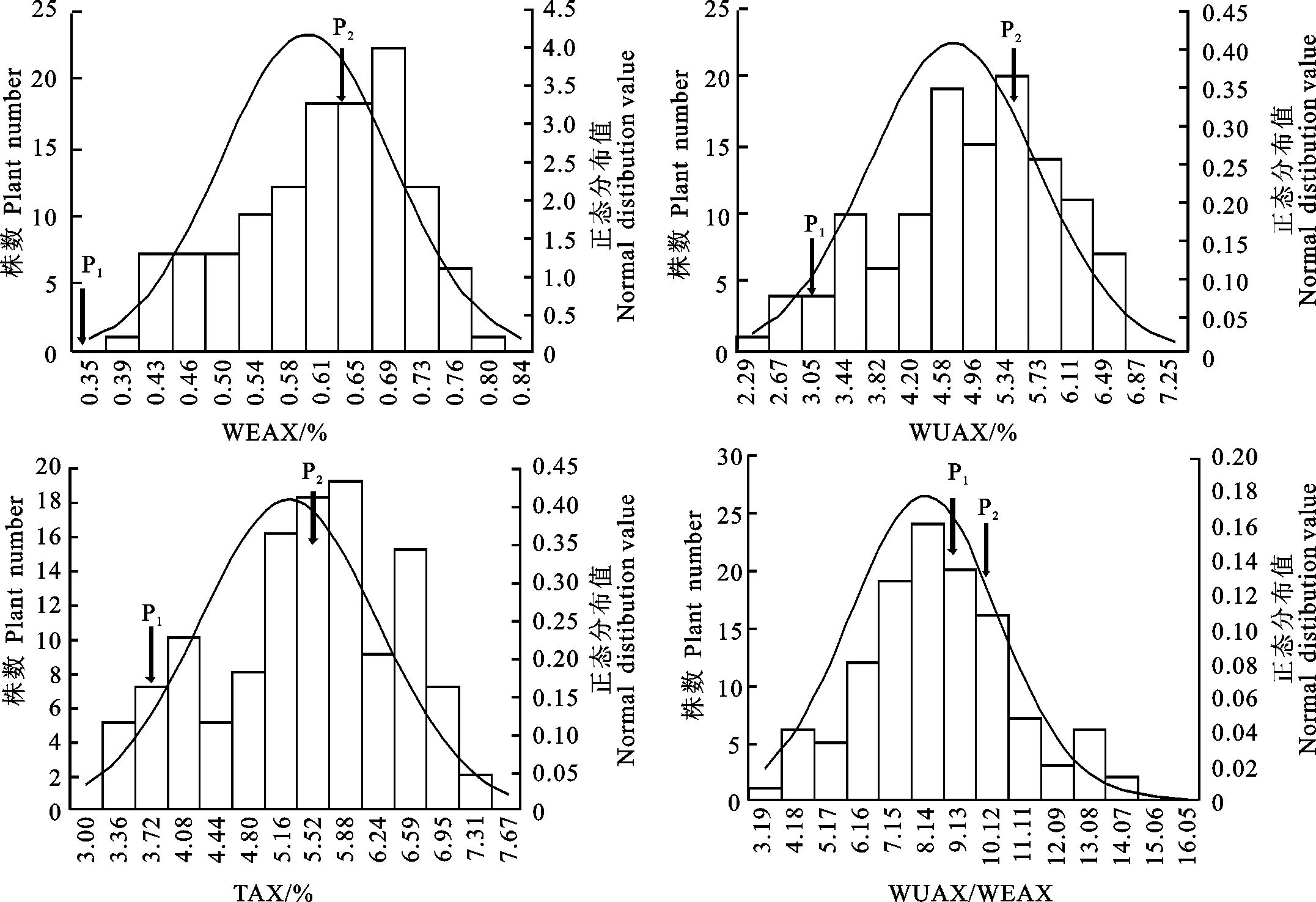
图1 青稞F2群体籽粒的WEAX、WUAX和AX含量及WUAX/WEAX的分布Fig.1 Distribution of grain WEAX,WUAX,TAX content and WUAX/WEAX in F2 population of hulless barley
2.2 青稞籽粒阿拉伯木聚糖及相关性状的相关性分析
对F2群体的TAX、WUAX和 WEAX含量及WUAX/WEAX值间进行相关分析,结果(表2)表明,WUAX/WEAX值与WEAX含量呈极显著负相关,与WUAX和TAX含量均呈极显著正相关;WUAX含量与TAX含量也呈极显著正相关。这表明青稞籽粒 WUAX与WEAX的合成受不同基因调控。
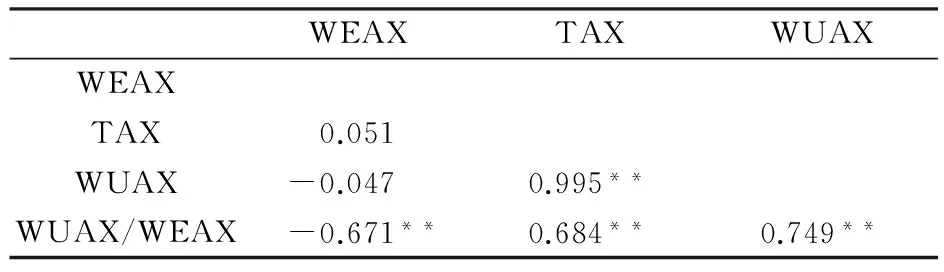
表2 青稞F2群体中的籽粒WEAX、TAX和WUAX含量及WUAX/WEAX值的相关性Table 2 Correlation analysis between grain WEAX,TAX,WUAX and WUAX/WEAX in F2 population of hulless barley
**:P<0.01.
2.3 青稞籽粒阿拉伯木聚糖及相关性状的最适遗传模型选择
通过SEA 软件包中的G4F2程序对P1、P2、F1和F2代进行联合表型遗传分析,青稞籽粒WEAX含量的最小AIC值遗传模型为MX2-AD-AD。MX1-AD-ADI、MX2-ADI-AD模型的AIC值与MX2-AD-AD相近,可以作为备选模型(表3)。经适应性检验,3个模型参数值显著的个数为均为6个(表4),故确定MX2-AD-AD模型为控制籽粒WEAX含量的最优模型,说明喜马拉雅8号和藏青320的F2群体的籽粒WEAX含量受2对加性-显性主基因+多基因控制,且多基因具有加性-显性效应。依据相同的方法与原则,青稞籽粒WUAX含量的最适遗传模型为MX2-AD-AD,该性状遗传符合2对显性-加性主基因+多基因方式(表3和表5);而籽粒TAX含量的最适遗传模型为MX1-AD-ADI,即其受1对加性-显性主基因+加性-显性-上位性多基因控制(表3和表6);WUAX/WEAX的最适遗传模型为MX2-EAED-AD,即其受2对等显等加性主基因+多基因控制,且多基因以加显性的方式对改性状进行影响(表3 和表7)。
表3 青稞籽粒阿拉伯木聚糖性状的候选遗传模型的MLV和AIC值Table 3 Log-maximum likelihood (MLV) and Akaike information criterion value (AIC) of candidate genetic model for grain arabinoxylan traits of hulless barley population
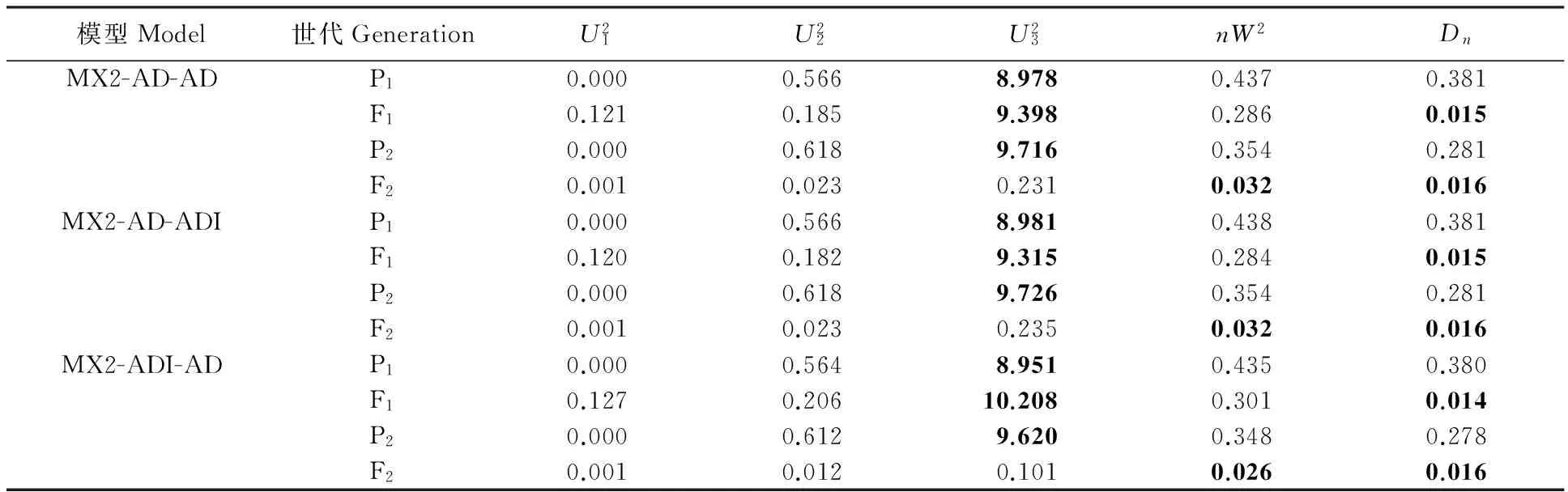
表4 青稞籽粒WEAX含量候选遗传模型的适合性检验Table 4 Goodness-of-fit test of candidate genetic models for grain WEAX content trait of hulless barley population
表中粗体值显著(P<0.05)。下表同。
Bold values in the table are significant at 0.05 level. The same in following tables.
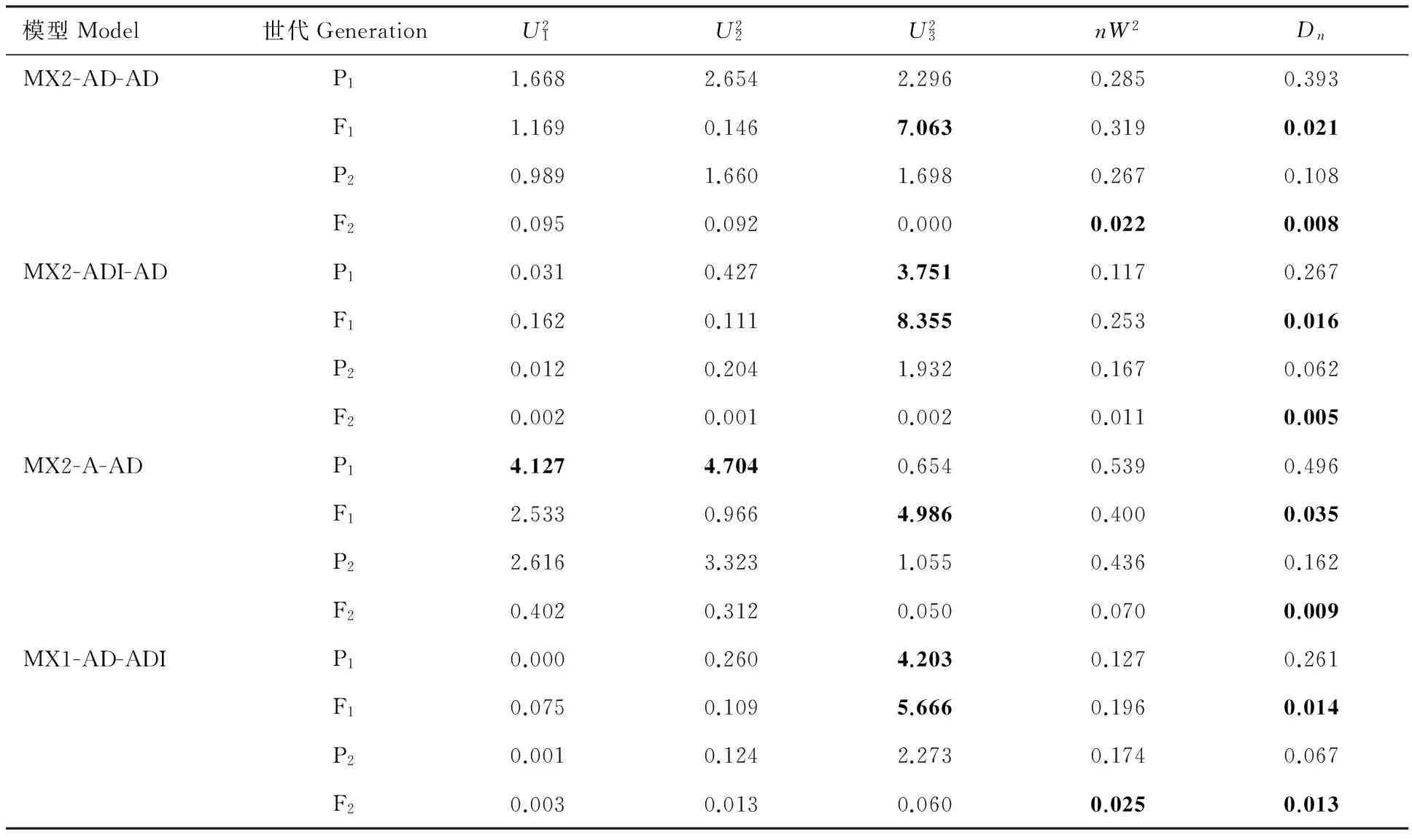
表5 青稞籽粒WUAX含量候选遗传模型的适合性检验Table 5 Goodness-of-fit test of candidate genetic models for grain WUAX content trait of hulless barley population
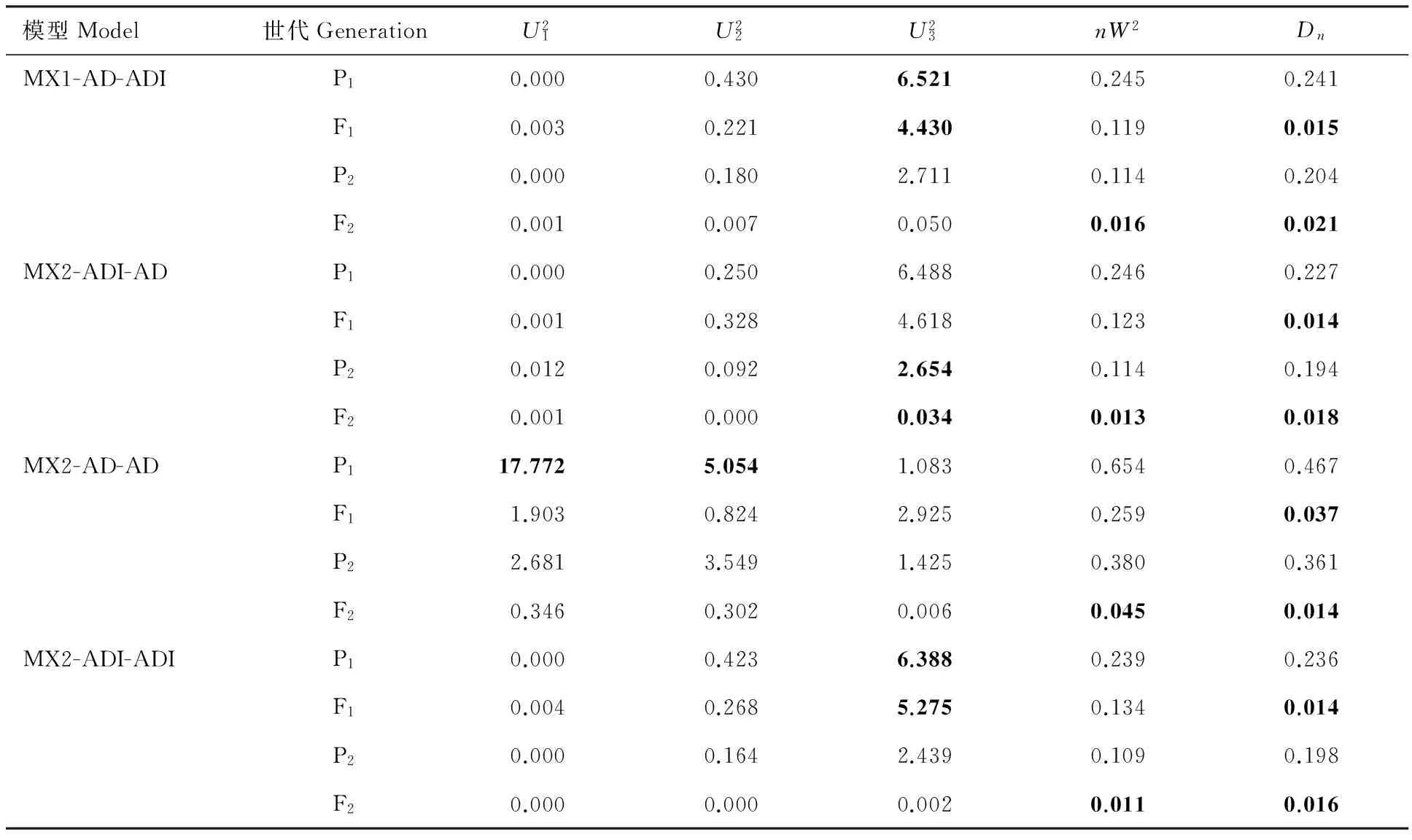
表6 青稞籽粒TAX含量候选遗传模型的适合性检验Table 6 Validation of candidate genetic models for grain TAX content trait of hulless barley population
2.4 青稞籽粒阿拉伯木聚糖及相关性状的遗传参数估计
利用最小二乘法对获得各个籽粒性状的最佳遗传模型进行相关遗传一阶和二阶参数估计(表8),结果表明,在MX2-AD-AD 模型下,影响籽粒WEAX含量的两对主基因的加性效应均为负值,且两对主基因的加性效应的绝对值略大于显性效应,表明这2对主基因具有降低籽粒WEAX的作用,其显性效应与加性效应比值的绝对值为0.923,表现为显性遗传。而多基因的加性效应略小于显性效应。进一步分析发现,2对主基因的遗传率为62.5%。在相同的模型影响下,控制籽粒WUAX含量的2对主基因的加性效应也为负值,且两对主基因的加性效应远大于显性效应,其比值分别为8.49和1.72,其显性效应与加性效应比值的绝对值为0.35,表现为部分显性。以加性遗传为主。其2对主基因的遗传率为88.84%,表明该性状受环境影响较小。在MX1-AD-ADI模型下控制青稞籽粒TAX含量性状的1对主基因的加性效应也远大于显性效应,其遗传率为82.37%,其显性效应与加性效应的比值的绝对值为0.14,表现为加性遗传为主。控制WUAX/WEAX的多基因的加性效应为主基因加性效应的2倍,主基因和多基因的遗传率总计高达86.91%,表明该性状的遗传主要受基因型控制,受环境的影响较小,同时从主基因的遗传方差占总表型方差的90.93%,也验证了该性状受环境影响较小的结论。

表7 青稞籽粒 WUAX/WEAX候选遗传模型的适合性检验Table 7 Validation of candidate genetic models for grain WUAX/WEAX ratio of hulless barley population
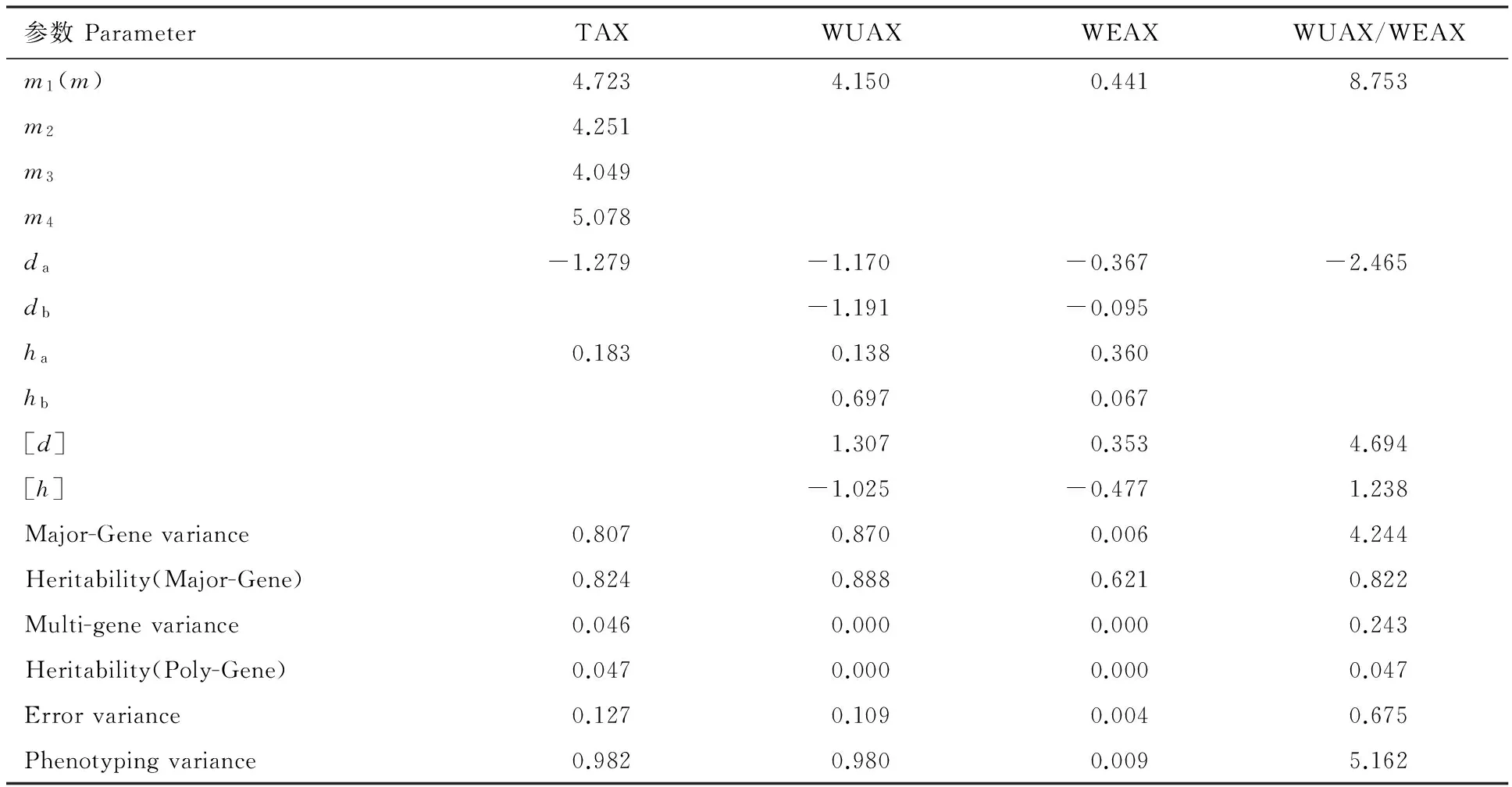
表8 青稞籽粒阿拉伯木聚糖性状最适模型遗传参数的估计Table 8 Estimated genetics parameters for grain arabinoxylan traits of hulless barley population with best-fitted models.
m为群体平均数;d和h分别代表主基因的加性效应和显性效应;a和b分别代表第1 对和第2 对主基因;[d]和[h] 分别代表多基因的加性效应和显性效应; Major-gene variance、Multi-gene variance、Error variance和Phenotyping variance分别代表主基因方差、多基因方差、误差方差、表型方差; Heritability (Major-gene)、Heritability (multi-gene)分别代表主基因遗传率、多基因遗传率。
mrepresents the mean of population;dandhmean the additive effect and dominant effect of major gene;aandbrefer to the 1st and 2nd major genes,respectively.[d] and [h] represent additive effect,dominant effect of multi-genes,respectively; major-gene variance,multi-gene variance,error variance and phenotyping variance indicate the mean square of major gene,multi-gene,error and phenotyping,respectively; heritability (Major-gene),heritability (multi-Gene) represent heritability of major gene and multi-gene,respectively.
3 讨 论
青稞因籽粒富含多种功能活性成分,日益受到食品工业者和植物遗传育种家的重视。阿拉伯木聚糖是一种重要的植物非淀粉性多糖,对植物细胞壁合成和逆境响应有着重要作用。前人对阿拉伯木聚糖遗传关系研究主要集中于普通大麦上,而作为我国特色的高原作物青稞却研究较少。Henry等[31]研究表明,大麦的阿拉伯木聚糖含量变异既受基因型控制,也受环境影响。不同用途及种皮类型大麦的阿拉伯木聚糖含量也存在较大的差异[17]。Zhang等[18]对8个二棱大麦在我国7个不同生态区种植获得的籽粒WEAX含量研究发现,环境因素对籽粒WEAX含量的影响高于基因型的影响,环境与基因型互作对其含量的变异呈显著相关;同时发现这些材料的籽粒WEAX、TAX的含量分别为0.27%~0.36%和4%~7%,而本研究的F2群体WEAX和TAX的平均含量为0.60%和 5.11%,WEAX的含量远大于Zhang等[18]的研究结果,这可能有两方面的原因,一是本研究的供试材料与前人的不同,二是可能与本研究种植的环境有一定关系。张晓勤等[29]的研究结果显示,氮肥运筹与种植地点显著影响大麦籽粒WEAX含量。而本研究材料种植地是灰色冲积土,这与Zhang等[18]研究中的种植环境存在着差异。
目前在小麦的1BL、7AS等多条染色体上发现了控制籽粒TAX、WEAX和WUAX含量的主效QTL[20]。Saulnier等[32]在小麦1AS和1BL 染色体上分别检测到一个控制籽粒WEAX含量的主效QTL,其解释表型变异率分别为17.5%和24.7%。在控制籽粒WEAX含量的QTL附近还发现了2个分别控制籽粒WUAX和TAX含量的主效QTL,其解释表型变异率为16.9%~29.8%。本研究利用主基因+多基因模型对喜马拉雅8号和藏青320构建的F2群体和两个亲本进行遗传分析,初步确定了青稞籽粒TAX含量受1对主基因+多基因控制,且主基因的遗传率比较高,主要以主基因遗传为主,环境因素对其影响较小。籽粒WEAX含量受2对主基因+多基因调控,2对主基因的遗传率为62.5%,籽粒WUAX也是受2对总遗传率为88.24%的主基因调控。这与姜丽娜等[33]对河南小麦研究获得的小麦籽粒WEAX、TAX的遗传率分别为91.0%和86.1%的结论相一致。而本研究获得分别控制青稞籽粒的WEAX和WUAX含量的2对主基因是连锁还是分布于不同染色体上,有待下一步进行研究。据相关研究发现,与大麦阿拉伯木聚糖相似性质的β-葡聚糖在分离后代中的变异主要由加性效应控制[34],这与本研究发现的控制大麦籽粒TAX含量的主基因的加性效应远大于显性效应的结论相似。同时,与青稞籽粒TAX极显著相关的WUAX也表现出相似趋势。这暗示WUAX的生物合成与TAX合成代谢存在着一定的内在生物学相关性。
本研究利用F2群体和其亲本考察了青稞籽粒阿拉伯木聚糖相关性状的分布及其遗传特点,发现籽粒WEAX、WUAX和TAX性状均受主基因调控,可以通过对其进行杂交、回交和系统选育等方法进行进一步的改良,但由于群体类型的限制,本研究的结论需要进一步完善。国外有文献[17]报道,皮大麦的TAX高于青稞,青稞籽粒阿拉伯木聚糖相关性状的遗传特性能否用于皮大麦育种,现在还未知。为了解决这些问题,后期我们一方面将继续进行RIL群体材料构建和建立一个皮裸大麦的双亲分离群体,运用多年多点数据进行完善和发掘影响阿拉伯木聚糖的遗传基因在不同种皮类型中的表现情况;另一方面,将对上述群体进行分子标记的基因型分型,从分子层面上解析控制皮大麦和青稞籽粒阿拉伯木聚糖及其相关性状的遗传机制。
[1]BAIK B K,ULLRICH S E.Barley for food:Characteristics,improvement,and renewed interest [J].JournalofCerealScience,2008,48(2):234.
[2]王显萍.青稞β-葡聚糖含量的基因型差异及优质种质的筛选[J].麦类作物学报,2013,33(1):185.
WANG X P.Genotypic difference of beta glucan content in hulless barley and identification of elite germpalsm [J].JournalofTriticeaeCrops,2013,33(1):185.
[3]KATO T,SASAKI A,TAKEDA G.Genetic variation of β-glucan contents and β-glucanase activities in barley,and their relationships to malting quality [J].BreedingScience,1995,45:471.
[4]赵慧芬.西藏裸大麦β-葡聚糖及食用纤维含量的遗传与环境效应研究[D].拉萨:西藏大学,2009:3.
ZHAO H F.Influence of genetic and environment on content of glucans and dietary fiber from hulless barley in Tibet [D].Lhasa:Tibet University,2009:3.
[5]孙元琳,陕方,赵立平.谷物膳食纤维-戊聚糖与肠道菌群调节研究进展[J].食品科学,2012,33(9):328.
SUN Y L,SHAN F,ZHAO L P.Research progress in pentosans as a kind of cereal dietary fiber and their gut microflora modulatory effect [J].FoodScience,2012,33(9):328.
[6]SKENDI A,BILIADERIS C G,LAZARIDOU A,etal.Structure and rheological properties of water soluble β-glucans from oat cultivars ofAvenasativa,andAvenabysantina[J].JournalofCerealScience,2003,38(1):17.
[7]王修启,李春喜,林东康,等.小麦中的戊聚糖含量及添加木聚糖复酶对鸡表观代谢能值和养分消化率的影响[J].动物营养学报,2002,14(3):58.
WANG X Q,LI C X,LIN D K,etal.The content of pentosan in wheat grain and effects of xylanase supplementation on AME values and nutrient digestibility of cocks [J].ChineseJournalofAnimalNutrition,2002,14(3):58.
[8]MURALIKRISHNA G,RAO M V.Cereal non-cellulosic polysaccharides:structure and function relationship-an overview [J].CriticalReviewsinFoodScienceandNutrition,2007,47(6):604.
[9]MICHNIEWICZ J,BILIADERIS C G,BUSHUK W.Effect of added pentosans on some properties of wheat bread [J].FoodChemistry,1992,43(4):251.
[10]GAMLATH J,ALDRED G P,PANOZZO J F.Barley (1-3;1-4)-β-glucan and arabinoxylan content are related to kernel hardness and water uptake [J].JournalofCerealScience,2008,47(2):365.
[11]冯 波,舒守贵,张爱民,等.麦类作物种子中戊聚糖研究进展[J].植物学报,2006,23(2):217.
FENG B,SHU S G,ZHANG A M,etal.Research progress on pentosan inTriticeaecrops [J].ChineseBulletinofBotany,2006,23(2):217.
[12]PHILIPPE S,TRANQUET O,UTILLE J P,etal.Investigation of ferulate deposition in endosperm cell walls of mature and developing wheat grains by using a polyclonal antibody [J].Planta,2007,225(5):1290.
[13]HAN J.Structural characteristics of arabinoxylan in barley,malt and beer [J].FoodChemistry,2000,70:134.
[14]EVANS D E,SHEEHAN M C,STEWART D C.The impact of malt derived proteins on beer foam quality.Part II:The influence of malt foam-positive proteins and non-starch polysaccharides on beer foam quality [J].JournaloftheInstituteofBrewing,1999,105(3):171.
[15]李春喜,邱宗波,姜丽娜,等.河南若干小麦品种籽粒戊聚糖含量的初步研究[J].西北植物学报,2002,22(5):1187.
LI C X,QIU Z B,JIANG L N,etal.Research on the content of pentosan in some winter wheat varieties in Henan [J].ActaBotanicaBoreali-occidentaliaSinica,2002,22(5):1187.
[16]张岐军,钱森和,张 艳,等.中国软质小麦品种戊聚糖含量的遗传变异及其与饼干加工品质的关系[J].中国农业科学,2005 ,38(9):1734.
ZHANG Q J,QIAN S H,ZHANG Y,etal.Variation of pentosans in Chinese soft wheat cultivars and correlations with cookie quality [J].ScientiaAgriculturaSinica,2005,38(9):1734.
[17]HOLTEKJOLEN A K,UHLEN A K,BRATHEN E,etal.Contents of starch and non-starch polysaccharides in barley varieties of different origin [J].FoodChemistry,2006,94(3):352.
[18]ZHANG X Q,XUE D W,WU F B,etal.Genotypic and environmental variation of arabinoxylan content and endoxylanase activity in barley grains [J].JournalofIntegrativeAgriculture,2013,12(8):1489.
[19]GAN Y,MCLEOD J G,SCOLES G J,etal.Extract viscosity of winter rye:heritability and correlation to kernel characters [J].PlantBreedSeedScience,1996,40(3-4):103.
[20]YANG L,ZHAO D,YAN J,etal.QTL mapping of grain arabinoxylan contents in common wheat using a recombinant inbred line population[J].Euphytica,2016,208(2):1.
[21]BORDES J,RAVEL C,GOUIS J L,etal.Use of a global wheat core collection for association analysis of flour and dough quality traits [J].JournalofCerealScience,2011,54(1):142.
[22]QURAISHI U M,MURAT F,ABROUK M,etal.Combined meta-genomics analyses unravel candidate genes for the grain dietary fiber content in bread wheat (TriticumaestivumL.) [J].Functional&IntegrativeGenomics,2011,11(1):71
[23]盖钧镒,章元明,王建康.植物数量性状遗传体系[M].北京:科学出版社,2003:15.
GAI J Y,ZHANG Y M,WANG J K.Genetic system of quantitative traits in plants [M].Beijing:Science Press,2003:15.
[24]章元明.植物数量遗传学的建立、发展与展望[J].南京农业大学学报,2012,35(5):20.
ZHANG Y M.Establishment,development and prospects of quantitative genetics in plant [J].JournalofNanjingAgriculturalUniversity,2012,35(5):20.
[25]吕亮杰,郭元世,杜丽杰,等.大麦籽粒淀粉含量的主基因十多基因遗传模型分析[J].麦类作物学报,2014,34(1):13.
LÜ L J,GUO S Y,DU L P,etal.Major genes plus polygenes mixed inheritance model for starch contents in barley seed [J].JournalofTriticeaeCrops,2014,34(1):13.
[26]杨晓梦,曾亚文,普晓英,等.大麦籽粒功能成分含量的遗传效应分析[J].麦类作物学报,2013,33(4):635.
YANG X M,ZENG Y W,PU X Y,etal.Inheritance analysis of functional components in barley grains [J].JournalofTriticeaeCrops,2013,33(4):635.
[27]巴桑玉珍,强小林.西藏青稞育种的成就与经验分析[J].西藏农业科技,2004,27(1):29.
BASANG Y Z,QIANG X L.Analysis on the achievement and experiences of hulless barley breeding in Tibet [J].TibetanAgriculturalScienceandTechnology,2004,27(1):29.
[28]陈 锋,夏先春,何中虎.普通小麦阿拉伯糖基木聚糖含量快速测定及其遗传特性分析[J].中国粮油学报,2007,22(5):143.
CHEN F,XIA X C,HE Z H.Arabinoxylans in common wheat:rapid determination and genetic analysis [J].JournaloftheChineseCereals&OilsAssociation,2007,22(5):143.
[29]张晓勤,薛大伟,张国平.不同环境及栽培条件下啤用大麦阿拉伯木聚糖含量的变化[J].作物学报,2013,39(9):1675.
ZHANG X Q,XUE D W,ZHANG G P.Changes of arabinoxylan content in malting barley under different environments and cultivation conditions [J].ActaAgronomicaSinica,2013,39(9):1675.
[30]曹锡文,刘 兵,章元明.植物数量性状分离分析Windows软件包SEA的研制[J].南京农业大学学报,2013,36(6):1.
CAO X W,LIU B,ZHANG Y M.SEA:a software package of segregation analysis of quantitative traits in plants [J].JournalofNanjingAgriculturalUniversity,2013.36(6):1.[31]HENRY R J.Genetic and environmental variation in the pentosan and beta-glucan contents of barley,and their relation to malting quality [J].JournalofCerealScience,1986,4(3):269.
[32]SAULNIER L,GUILLON F,SADO P E,etal.Plant cell wall polysaccharides in storage organs:xylans (food applications) [J].ComprehensiveGlycoscience,2007(1991):653.
[33]姜丽娜,邵 云,张 霞,等.小麦籽粒戊聚糖含量及相关遗传特性分析[J].麦类作物学报,2004,24(4):28.
JIANG L N,SHAO Y,ZHANG X,etal.Studies on the pcentosan content in wheat grains and analysis on its genetic characteristics [J].JournalofTriticeaeCrops,2004,24(4):28.
[34]POWELL W,CALIGARI P D,SWANSTON J S,etal.Genetical investigations into β-glucan content in barley [J].TheoreticalandAppliedGenetics,1985,71(3):463.
Genetic Analysis of Arabinoxylan Content in Grain of Hulless Barley
LIU Xinchun1,BASANG Yuzhen1,2,FENG Zongyun1
(1.Barley and Hulless Barley Research Centre,College of Agronomy,Sichuan Agricultural University,Chengdu,Sichuan 611130,China; 2.Agricultural Research Institute,Tibet Academy of Agriculture and AnimalHusbandy Sciences,Lhasa,Tibet 850032,China)
In order to understand the genetics model and genetics basis of arabinoxylan and the related traits of hulless barley,a set of F1and F2population derived from a cross between Himalaya 8 and Zangqing 320,along with their parents were employed to uncover the genetics basis of grain AX content by using the methodology called “mixed major gene plus multi-gene inheritance model”. The result showed that significant correlation between water-unextractable arabinoxylan (WUAX) and total arabinoxylan (TAX) were detected,while WUAX was not significantly correlated with water-extractable arabinoxylan (WEAX). TAX was controlled by one major gene with dominant-additive effect but without polygenic affect. WEAX,WUAX and WUAX/WEAX were controlled by two major gene with polygenic effect,and the expression patterns of the major gene were different among three traits. The additive effect of the major genes controlling WEAX and WUAX were negative,which suggested as a negative regulatory factor. The additive effects of major genes controlling all the traits,except for WEAX,were strong. The major gene with high heritability,with a range from 62.5% to 88.49%,determined all the trait variation in this research.
Hulless barley; Arabinoxylan; Major gene plus multi-gene; Genetic evaluation
时间:2017-07-07
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20170707.1815.010.html
2017-01-05
2017-03-07
国家现代农业产业技术体系(大麦青稞)建设专项(CARS-05)
E-mail:ku0082005@126.com
冯宗云(E-mail:zyfeng49@126.com)
S512.3;S330
A
1009-1041(2017)07-0890-10
