庙岛群岛海域网采浮游植物种类组成及分布
2017-08-12王文君郝彦菊
喻 龙,王 磊,王文君,李 峰,郝彦菊
(1.烟台市海洋环境监测预报中心,山东烟台264003;2.中国农业大学烟台研究院,山东烟台264670)
庙岛群岛海域网采浮游植物种类组成及分布
喻 龙1,王 磊1,王文君1,李 峰1,郝彦菊2*
(1.烟台市海洋环境监测预报中心,山东烟台264003;2.中国农业大学烟台研究院,山东烟台264670)
于2015-05(春季)、2015-08(夏季)对北至北隍城岛、南至蓬莱近岸的庙岛群岛海域进行网采浮游植物调查。样品共鉴定出67种浮游植物物种,其中硅藻60种,甲藻5种,甲藻孢囊1种,金藻1种。该海区春季浮游植物群落优势种为具槽帕拉藻(Paralia sulcata,优势度0.384)、刚毛根管藻(Rhizosolenia setigera,优势度0.319)、冰河拟星杆藻(Asterionellopsis glacialis,优势度0.054)、布氏双尾藻(Ditylum brightwellii,优势度0.021);夏季优势种为高齿状藻(Odontella regia,优势度0.787),庙岛海峡南部海区出现高齿状藻的密集分布区域。夏季浮游植物总丰度是春季的2.17倍。春、夏季浮游植物多样性指数和均匀度指数都较低,且其数量的时间、空间分布变化明显。春季,浮游植物细胞丰度、主要优势种数量、多样性指数、种类丰富度指数的空间分布特征为海区北部高,南部低;夏季与春季相反,为海区南部高,北部低。
庙岛群岛海域;网采浮游植物;种类组成;优势种;空间分布
海洋浮游植物作为初级生产者是海洋食物链的第一环节,是大多数浮游动物和滤食性贝类的饵料,在海洋生态系统的物质循环和能量转换过程中起到重要的作用。浮游植物的种类组成和数量分布与渔业关系较为密切[1-4]。其群落结构、生物量及生产力的变化将直接影响到海洋生态系统的结构与功能,同时也对气候变化和人为活动做出响应[5-8]。
庙岛群岛(120°36'~120°56'E,37°53'~38°23'N)位于山东半岛和辽东半岛之间,地处渤海海峡,为黄海和渤海的交汇处,由32个岛屿组成,地理位置特殊。庙岛群岛海域浮游植物和浮游动物种类较多,各岛周围海域底栖生物种类繁多,是多种鱼类洄游必经之路,海洋哺乳动物资源比较丰富,并分布有多处水产养殖区,生态环境特殊[9-11]。
我们于2015-05(春季)、2015-08(夏季)对北至北隍城岛、南至蓬莱近岸的整个庙岛群岛海域进行了取样调查,分析网采浮游植物群落结构,为庙岛群岛海域生态系统的研究和保护提供基础资料和科学依据。
1 调查站位与方法
2015-05和2015-08在庙岛群岛海域共设22个站位(图1)。
按照《海洋监测规范》[13],样品采集使用浅水Ⅲ型网,自水体底层至表层垂直拖取,样品用体积分数为5%的福尔马林溶液固定保存,在实验室奥林巴斯CKX-41倒置显微镜下进行种类分类鉴定和个体计数,计算总丰度(个·m-3)。数据用Microsoft Excel软件进行统计处理,用Surfer 8软件制图。
(高 峻 编辑)

图1 2015-05和2015-08庙岛群岛海域浮游植物调查站位图Fig.1 Phytoplankton survey sites in the Miaodao Archipelago waters in May and Agust,2015
浮游植物群落多样性指数Shannon-Wiener指数H':

优势度指数Y:

种类丰富度指数D:

均匀度指数J':

式中:S为种类数;Pi为i种所占比例;ni为第i种的细胞数量;N为总个体数;fi为该种在各站位出现频率。
2 结 果
2.1 种类组成
本次调查采集的浮游植物样品共鉴定出67种浮游植物物种(表1),其中硅藻门60种,占总种数的89.55%;甲藻门5种,并有1种甲藻孢囊,共占8.96%;金藻门1种,占1.49%。
2.2 各调查站位的丰度分布
春季(2015-05),海区各调查站网采浮游植物丰度为(1.99~86.78)×105个·m-3,平均值为15.22×105个·m-3,以具槽帕拉藻(Paralia sulcata)、刚毛根管藻(Rhizosolenia setigera)、冰河拟星杆藻(Asterionellopsis glacialis)、布氏双尾藻(Ditylum brightwellii)、圆筛藻(Coscinodiscus sp.)等种类为主,长山水道以南的海区丰度较低、以北丰度较高,总体分布为海区西南部较低,东北部较高(图2)。
夏季(2015-08),各站位网采浮游植物丰度为(0.336~576.320)×105个·m-3,平均达33.04×105个· m-3,为春季的2.17倍,以高齿状藻(Odontella regia)、中华半管藻(Hemiaulus sinensis)、角毛藻(Chaetoceros sp.)、泰晤士旋鞘藻(Helicotheca tamesis)、波状石丝藻(Lithodesmium undulatus)等种类为主。但夏季浮游植物空间分布极不均匀,海区南部庙岛海峡近岸丰度最高,密集分布明显。北长山岛西部丰度较高,东部和以北的海区丰度较低,均低于1×105个·m-3(图3)。

表1 浮游植物种类名录Table 1 Species list of phytoplankton

图2 春季丰度分布(×105个·m-3)Fig.2 Distributions of abundance in spring(×105ind·m-3)
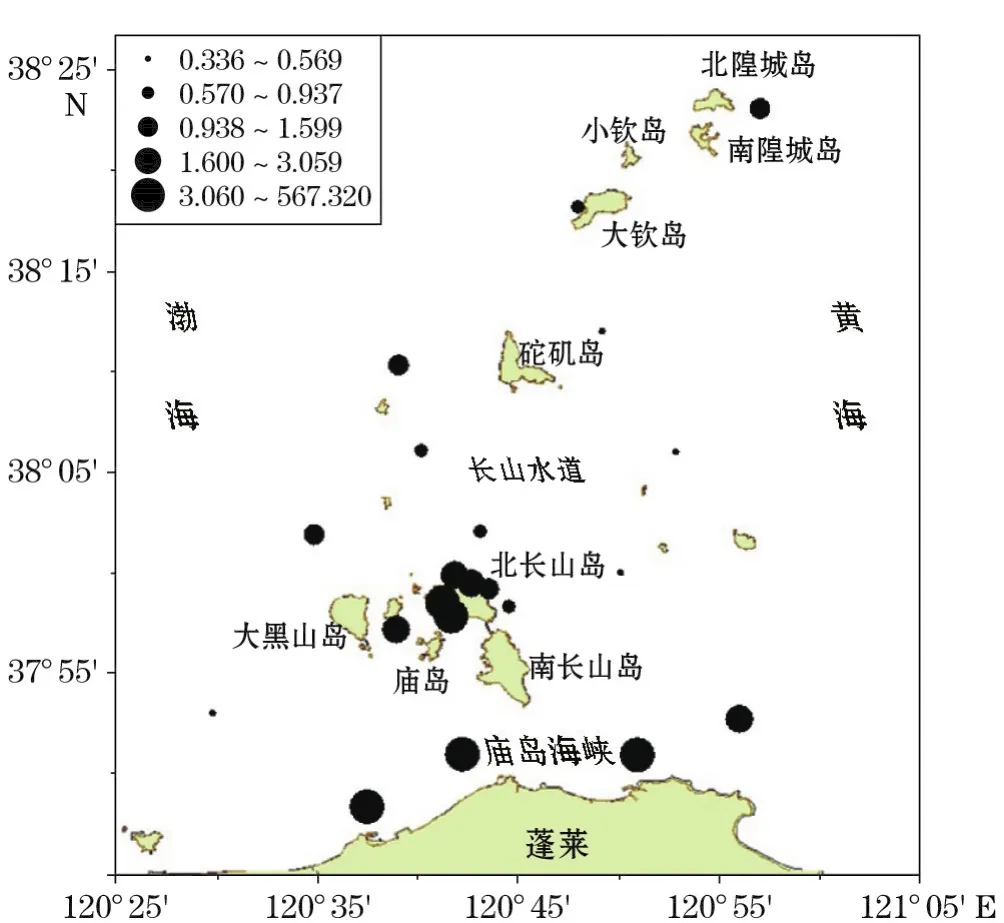
图3 夏季丰度分布(×105个·m-3)Fig.3 Distributions of abundance in summer(×105ind·m-3)
2.3 优势种及其丰度分布
2.3.1 优势种
庙岛群岛海区浮游植物主要类群的优势度计算结果见表2。依据优势度Y>0.02时定为优势种,春季庙岛群岛海区浮游植物优势种为具槽帕拉藻、刚毛根管藻、冰河拟星杆藻、布氏双尾藻四种;夏季优势种仅为高齿状藻,显著不同于春季。该海区浮游植物优势种的优势度较为明显。如春季具槽帕拉藻和刚毛根管藻的优势度分别为0.384和0.319,夏季高齿状藻的优势度则高达0.787。

表2 庙岛群岛海区浮游植物主要种类及其优势度Table 2 The main species and dominances of phytoplankton in the Miaodao Archipelago waters
2.3.2 主要优势种丰度分布
春季(2015-05),具槽帕拉藻为庙岛群岛海区的主要优势种之一。具槽帕拉藻属于世界性广布的半咸水、近岸性海水底栖种,受海流、潮汐等作用常出现在浮游生物种群中,在中国沿海皆有分布。本次调查中,具槽帕拉藻在各调查站位中出现的频率为100%,平均丰度为5.85×105个·m-3,总丰度占所检浮游植物细胞总量的38.44%。其数量分布由南向北逐渐增多,并在南、北长山岛沿岸有较高的数量分布(图4)。1,2, 21号站浮游植物细胞丰度较低,其中具槽帕拉藻的数量虽然也不高,但其在浮游植物细胞总量中所占的比例却很高,均高于90%;在浮游植物细胞丰度较低的3,15,16,17,18,19,20,22号站,具槽帕拉藻数量占各自站位浮游植物细胞总量的比例亦较高,均高于80%。
该海区2015-05的另一主要优势种刚毛根管藻为沿岸广温、广盐性种,早春或盛夏为盛产期,在我国南海、东海、黄海和渤海均有分布。本次调查出现频率为90.90%,平均丰度为5.87×105个·m-3,总丰度占所检浮游植物细胞总量的35.07%。其数量在北长山岛以西、以南的海区较少,长山水道以北数量分布显著升高(图5)。砣矶岛以西的5号站、长山水道东部的10号站浮游植物细胞丰度最高,这2个站位刚毛根管藻的数量亦高,所占比例均高于52%。
夏季(2015-08),庙岛群岛海区的主要优势种为高齿状藻。高齿状藻属于暖温带至热带近海种,在中国沿海皆有分布。本次调查除13号站(大钦岛西北岸)外,其余站位均出现高齿状藻,但丰度分布极不均匀,除15号站(北长山岛西南)、20号站、21号站、22号站(庙岛海峡南部)细胞丰度超过1×105个·m-3外,其余站位细胞丰度均低于1×105个·m-3;尤其是20号站(庙岛海峡东南),丰度达到529.12×105个·m-3,为海区平均值的19.4倍,占整个海区浮游植物细胞总数的76.26%,占该站位浮游植物细胞总数的91.81%。在本次调查中,高齿状藻细胞丰度的空间分布呈北长山岛的西部、南部数量高,东部、北部数量低的趋势(图6)。整个调查海区的平均丰度为27.29×105个·m-3,细胞丰度占整个调查海区浮游植物细胞总数的82.61%。

图4 春季具槽帕拉藻数量分布(×105个·m-3)Fig.4 Distributions of Paralia sulcata in spring (×105ind·m-3)
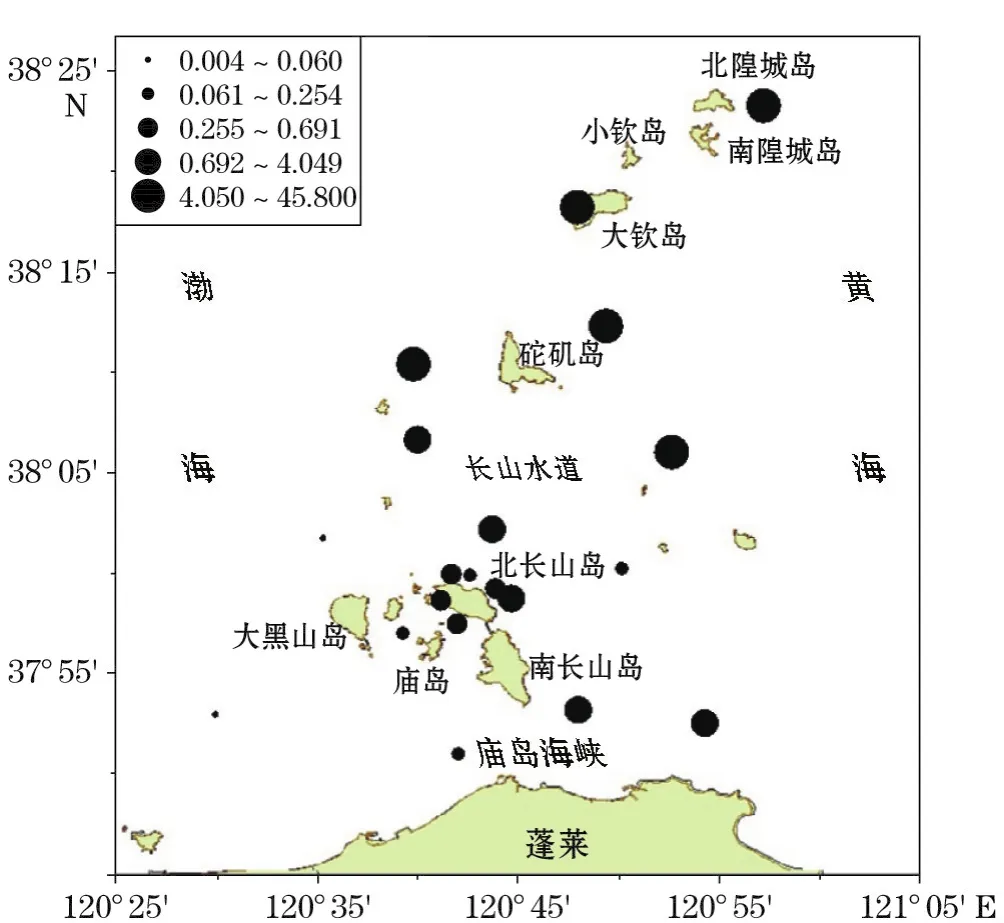
图5 春季刚毛根管藻数量分布(×105个·m-3)Fig.5 Distributions of Rhizosolenia setigera in spring (×105ind·m-3)

图6 夏季高齿状藻数量分布(×105个·m-3)Fig.6 Distributions of Odontella regia in summer(×105ind·m-3)
2.4 多样性、均匀度和种类丰富度指数
2015年春季(2015-05)和夏季(2015-08),整个调查海区浮游植物多样性指数均较低。春季,浮游植物多样性指数范围为0.437~2.547,平均为1.187;均匀度指数范围0.126~0.600,平均0.303;种类丰富度指数范围0.469~1.360,平均为1.006。多样性指数和均匀度指数分布特征相似,均是长山水道以北和南、北长山岛以东的海区较高(图7和图8)。而种类丰富度指数分布则是在北长山岛东北沿岸及其以北的海区较高(图9)。夏季(图10)调查海区浮游植物多样性指数范围0.666~3.382,平均为2.354,分布特征为北长山岛附近海区和庙岛海峡南部海区较高;均匀度指数范围0.145~0.823,平均为0.669,北长山岛附近及东部海区较高(图11);种类丰富度指数范围0.501~1.784,平均为0.961,与多样性指数的分布特征相似(图12)。2015-08浮游植物多样性指数和均匀度指数虽高于2015-05的,但优势种高齿状藻数量最多的20号站(庙岛海峡东南),其多样性指数和均匀度指数都最低。
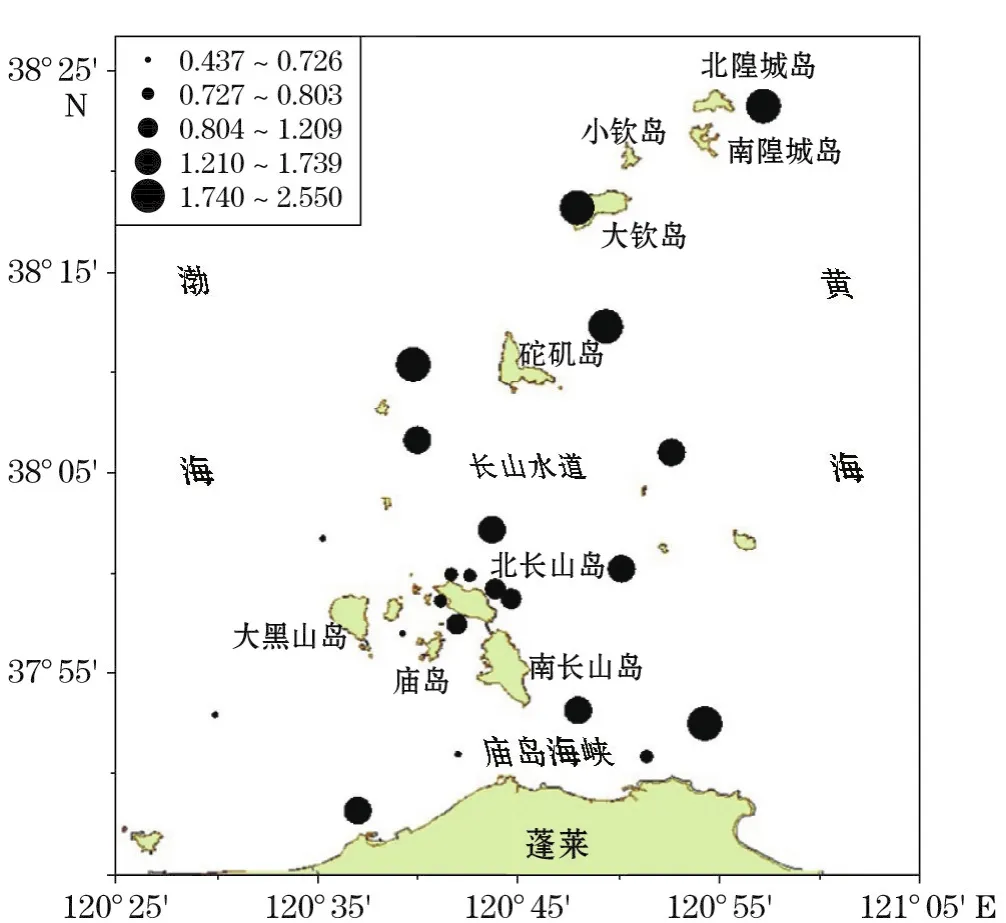
图7 2015年春季浮游植物多样性指数分布Fig.7 Distributions of phytoplankton diversity index in spring

图8 2015年春季浮游植物均匀度指数分布Fig.8 Distributions of phytoplankton uniformity index in spring

图9 2015年春季种类丰富度指数分布Fig.9 Distributions of species richness index in spring

图10 2015年夏季浮游植物多样性指数分布Fig.10 Distributions of phytoplankton diversity index in summer

图11 2015年夏季浮游植物均匀度指数分布Fig.11 Distributions of phytoplankton uniformity index in summer

图12 2015年夏季种类丰富度指数分布Fig.12 Distributions of species richness index in summer
3 讨 论
庙岛群岛海域隶属渤海海域,地处渤海海峡中部海区。关于渤海浮游植物研究的历史资料较多。与前人的研究[10,12,14-16]相比,本次调查中鉴定出的物种在种类组成和分布上差异较大。
结果分析显示,调查海区浮游植物的细胞丰度较高,这与渤海海区浮游植物具有春、夏季双峰值的特征[17]一致。1998年,渤海近岸浮游植物数量高峰出现在春季(5月)和秋季(10月)[18];从丰度的空间分布看,1999年春季(4—5月)渤海海区浮游植物斑块是其平面分布的主要特征,其丰度的高值区出现在渤海中部靠近渤海海峡处[15](庙岛群岛以西海区)。本次调查研究中,2015-05北部海区浮游植物的丰度较高,而2015-08南部海区浮游植物的丰度较高。春季各站位具槽帕拉藻在浮游植物群落中所占的比例均超过80%,丰度较低;而刚毛根管藻在浮游植物群落中所占的比例均超过52%,且丰度较高,但未形成斑块。黄风洪等[10]亦发现冬季庙岛海峡附近浮游植物细胞丰度较高。
2015年春、夏季浮游植物数量的时间、空间分布变化明显。浮游植物细胞丰度、主要优势种数量、多样性指数、种类丰富度指数的空间分布在2015-05与2015-08呈大致相反的趋势(图2~图6):2015-05北高南低,2015-08南高北低;浮游植物种间数量分布不均匀,多样性指数和均匀度指数都较低。
孙萍等研究[16]表明,2005-08渤海网采浮游植物群落的多样性随细胞丰度值的增加而降低。而本次调查中,浮游植物群落的多样性和细胞丰度的空间分布趋势大致相似,仅庙岛海峡西南部(20号站)较为异常——该站位出现密集成团的高齿状藻,浮游植物种类多达24种,细胞丰度高,但多样性指数低(图3和图10)。黄风洪等研究[12]发现,2013-08北长山岛西南岸(本次研究中的15号站)表层水样中浮游植物物种数最少,仅圆筛藻和具槽帕拉藻两种;在南长山岛以西最远的调查站位处(本次研究中的1号站位附近海区),浮游植物只有3种,且发现有密集成团的具槽帕拉藻。但是我们研究主要为网采浮游植物分析,而近几年对该海域浮游植物的研究多为水样分析,无法与本次研究结果进行有效比较。因而,本次研究结果可为该海域网采浮游植物的后续研究提供更准确的参考。
以往的研究表明,黄海海区和渤海海区浮游植物种类组成在不同的调查时间内变化明显。董婧等[19]发现,1998-06黄海北部近岸水域中甲藻数量占浮游植物细胞总数的比重高达50.45%。孙军等[15]发现,渤海海峡浮游植物群落在1999年春季(4—5月)主要由硅藻、甲藻组成,以冰河拟星杆藻、虹彩圆筛藻(Coscinodiscus oculus-iridis)、太阳双尾藻(Ditylum sol)、刚毛根管藻等小型细胞硅藻占优势的群落为主。孙晓霞等[20]2011年对黄海、东海浮游植物粒级结构的研究也表明,夏季(8月)中小型的表层浮游植物对海区Chl-a总量的贡献率最大。1998-09—10渤海海峡以大型细胞硅藻-甲藻联合的优势群落为主,群落季节和周年的变化以种类演替表明浮游植物群落由硅藻占绝对优势逐渐转变为硅藻-甲藻共存为主的群落[14];2005-08渤海的浮游植物以硅藻为主,优势种主要为中肋骨条藻(Skeletonema costatum)、拟旋链角毛藻(Chaetoceros pseudocurvisetus)、菱形海线藻(Thalassionema nitzschioides)和旋链角毛藻(Chaetoceros curvisetus)[16]; 2011-11—12主要由硅藻和甲藻组成,还有少量的金藻,主要优势种为具槽帕拉藻、偏心圆筛藻(Coscinodiscus excentricus)、梭角藻(Ceratium fusus)、相似曲舟藻(Pleurosigma affine)、星脐圆筛藻(Coscinodiscus asteromphalus)和菱形藻(Nitzschia sp.)等[21]。庙岛群岛南部海域2013年冬季(2013-02)表层水样中浮游植物优势种为太平洋海链藻(Thalassiosira pacirica)、具槽帕拉藻、加拉星平藻(Asteroplanus karianus)、离心列海链藻(Thalassiosira excentricus)和圆海链藻(Thalassiosira rotula)[10];2013-08优势种为圆筛藻、具槽帕拉藻[12]。本次研究结果显示,2015-05庙岛群岛海域浮游植物优势种为具槽帕拉藻、刚毛根管藻、冰河拟星杆藻、布氏双尾藻;2015-08优势种为高齿状藻。
综上所述,该海域浮游植物优势种的变迁无显著规律。浮游植物群落组成、浮游植物数量和空间分布的变化反映了其栖息环境的变化。庙岛群岛海区水文、营养环境的变化与浮游植物群落的季节变化、年际变化的关系有待于更深入、全面的研究。
4 结 论
1)2015年春、夏季庙岛群岛海区网采浮游植物共鉴定出67种。其中硅藻门60种,占总种数的89.55%;甲藻门5种,并有1种甲藻孢囊,共占8.96%;金藻门1种,占1.49%。
2)庙岛群岛海区春季浮游植物群落优势种为具槽帕拉藻、刚毛根管藻、冰河拟星杆藻、布氏双尾藻共4种;夏季优势种只1种,为高齿状藻,庙岛海峡南部出现高齿状藻的密集分布区域。
3)浮游植物细胞丰度、主要优势种数量、多样性指数、指数丰富度指数的空间分布季节差异性较大:春季北高南低,夏季南高北低。浮游植物种间数量分布不均匀,多样性指数和均匀度指数都较低。
[1] CARDINALE B J,PALMER M A,COLLINS S L.Species diversity enhances ecosystem functioning through interspecific facilitation[J]. Nature,2002,415:426-429.
[2] HAO Y J,WANG Z L,ZHU M Y,et al.Investigation and assessment of nutrients and phytoplankton diversity in the Laizhou Bay[J]. Advances in Marine Science,2005,23(2):197-204.郝彦菊,王宗灵,朱明远,等.莱州湾营养盐与浮游植物多样性调查与评价研究[J].海洋科学进展,2005,23(2):197-204.
[3] ZHANG Z L,YE S Z,YE Q T.The species composition and quantity distribution characteristics of phytoplankton in Fujian sea area[J]. South China Fisheries Science,2006,2(5):45-50.张壮丽,叶孙忠,叶泉土.福建海区浮游植物种类组成及数量分布特点[J].南方水产,2006,2(5):45-50.
[4] ZHANG X J,WEN R B,YU H Y,et al.Seasonal changes of the structure in the phytoplankton community Huanghua Waters[J].Ad-vances in Marine Science,2013,31(2):221-228.张晓举,温若冰,于海洋,等.黄骅海域浮游植物群落结构的季节变化[J].海洋科学进展,2013,31(2):221-228.
[5] SAEMIENTO J L,TOGGWEILER J R,NAAJJAR R.Ocean carbon-cycle dynamics and atmospheric pCO2[J].Philosophical Transactions of the Royal Society of London A,1988,325:3-21.
[6] CADEE G C,HEGEMAN J.Phytoplankton in the Marsdiep at the end of the 20th century:30 years monitoring biomass,primary production,and Phaeocystis blooms[J].Journal of Sea Research,2002,48(2):97-110.
[7] PODER T,MAESTRINI S Y,BALODE M,et al.The role of inorganic and organic nutrients on the development of phytoplankton along a transect from the Daugava River mouth to the Open Baltic,in spring and summer 1999[J].Ices Journal of Marine Science,2003,60: 827-835.
[8] HUANG B Q,HU J,LIU X,et al.Review on phytoplankton community structure and its coupling with biological carbon pump under global climate changes[J].Journal of Xiamen University(Natural Science),2011,50(2):402-410.黄邦钦,胡俊,柳欣,等.全球气候变化背景下浮游植物群落结构的变动及其对生物泵效率的影响[J].厦门大学学报(自然科学版),2011,50(2):402-410.
[9] FAN G K,HAN J B,HUANG J C,et al.The spotted seals in Miaodao Islands[J].Fisheries Science,2005,24(3):16-18.范国坤,韩家波,黄继成,等.庙岛群岛海域斑海豹的分布与保护[J].水产科学,2005,24(3):16-18.
[10] HUANG F H,WANG Y Y,LI J,et al.Phytoplankton community structure and the relationship to environment in southern waters of Miaodao Archipelago in winter in 2013[J].Journal of Waterway and Harbor,2015,36(3):258-263.黄风洪,王媛媛,李捷,等.冬季庙岛群岛南部海域浮游植物群落结构及其与环境因子的关系[J].水道港口,2015,36(3):258-263.
[11] XU Y D,WEI X,WU X W,et al.Characteristics of nutrients and eutrophication assessment in the southern waters of Miaodao Archipelago[J].Science Technology and Engineering,2015,15(22):1671-1851.徐艳东,魏潇,吴兴伟,等.庙岛群岛南部海域海水营养盐特征和富营养化评价[J].科学技术与工程,2015,15(22):1671-1851.
[12] HUANG F H,SHI H H,ZHENG W,et al.Phytoplankton diversity distribution and its influence factors in southern waters of Miaodao Archipelago in summer[J].Marine Environmental Science,2015,34(4):530-535.黄风洪,石洪华,郑伟,等.夏季庙岛群岛南部海域浮游植物多样性分布及其影响因子[J].海洋环境科学,2015,34(4):530-535.
[13] National Technical Comnittees of Ocean Standardization.Specification for marine monitoring:part 7 Ecological investigation and biological monitoring of offshore pollution:GB 17378.7-2007[S].Beijng:China Standard Press,2008.全国海洋标准化技术委员会.海洋监测规范:第7部分 近海污染生态调查和生物监测GB 17378.7-2007[S].北京:中国标准出版社,2008.
[14] SUN J,LIU D Y,YANG S M,et al.The preliminary study on phytoplankton community structure in the central Bohai Sea and the Bohai Strait and its adjacent area[J].Oceanologia et Limnologia Sinica,2002,33(5):461-471.孙军,刘东艳,杨世民,等.渤海中部和渤海海峡及邻近海域浮游植物群落结构的初步研究[J].海洋与湖沼,2002,33(5):461-471.
[15] SUN J,LIU D Y,XU J,et al.The netz-phytoplankton community of the central Bohai Sea and its adjacent waters in spring 1999[J]. Acta Ecologica Sinica,2004,24(9):2003-2016.孙军,刘东艳,徐俊,等.1999年春季渤海中部及其邻近海域的网采浮游植物群落[J].生态学报,2004,24(9):2003-2016.
[16] SUN P,LI R X,LI Y,et al.The net-phytoplankton community structure of the Bohai Sea in late summer,2005[J].Advances in Marine Science,2008,26(3):354-363.孙萍,李瑞香,李艳,等.2005年夏末渤海网采浮游植物群落结构[J].海洋科学进展,2008,26 (3):354-363.
[17] LIU F F,CHEN X E.Simulation on seasonal variation of the phytoplankton biomass in the Bohai Sea[J].Periodical of Ocean University of China(Natural Science),2014,44(2):17-26.刘菲菲,陈学恩.渤海浮游植物生物量季节变化的模拟研究[J].中国海洋大学学报(自然科学版),2014,44(2):17-26.
[18] WANG J.Species composition and quantity variation of phytoplankton in inshore waters of the Bohai Sea[J].Marine Fisheries Research, 2003,24(4):44-50.王俊.渤海近岸浮游植物种类组成及其数量变动的研究[J].海洋水产研究,2003,24(4):44-50.
[19] DONG J,LIU H Y,BI Y B,et al.Ecological study on planktonic dinoflagellates in inshore of Northern Yellow Sea[J].Marine Fisheries Research,2002,23(4):46-50.董婧,刘海映,毕远溥,等.黄海北部近岸的浮游甲藻生态[J].海洋水产研究,2002,23(4):46-50.
[20] SUN X X,REN L L,ZHENG S,et al.Phytoplankton size structurein the Yellow Sea and East China Sea in the spring and summer of 2011[J].Oceanologia et Limnologia Sinica,2012,43(3):419-428.孙晓霞,任琳琳,郑珊,等.2011年春夏季黄、东海浮游植物粒级结构[J].海洋与湖沼,2012,43(3):419-428.
[21] GUO S J,LI Y Q,ZHANG C X,et al.Phytoplankton community in the Bohai Sea and its relationship with environmental factors[J]. Marine Science Bulletin,2014,33(1):95-105.郭术津,李彦翘,张翠霞,等.渤海浮游植物群落结构及与环境因子的相关性分析[J].海洋通报,2014,33(1):95-105.
The Net-phytoplankton Species Composition and Distribution of the Miaodao Archipelago Waters
YU Long1,WANG Lei1,WANG Wen-jun1,LI Feng1,HAO Yan-ju2
(1.Yantai Monitoring and Forecasting Center of Marine Environment,Yantai 264003,China; 2.Yantai Research Institute,Agriculture University of China,Yantai 264670,China)
The net-phytoplankton of the Miaodao Archipelago waters was investigated in May(spring)and August(summer),2015.67 phytoplankton species were identified,including 60 diatom species,5 dinoflagellate species,1 dinoflagellates cysts,and 1 chrysophyta species.In spring,Paralia sulcata (dominance 0.384),Rhizosolenia setigera(dominance 0.319),Asterionellopsis glacialis(dominance 0.054),and Ditylum brightwellii(dominance 0.021)were the dominant species.In summer,Odontella regia(dominance 0.787)was the only dominant species,and there was mass Odontella regia distributing in the south of Miaodao strait.The total phytoplankton cell abundance in summer was 2.17 times of that in spring.Both phytoplankton diversity index and uniformity index were low in spring and summer,the temporal and spatial distributions of phytoplankton quantities were obviously variational.In spring,the spatial distributions of phytoplankton abundance,the quantities of main dominant species,diversity index,and species richness index were lower in the south and higher in the north of the sea area.However,the characteristic of spatial distribution of these parameters were opposite in summer,demonstrating higher in the south and lower in the north of the sea area.
the Miaodao Archipelago waters;net-phytoplankton;species composition;dominant species; spatial distribution
May 3,2016
Q178.53
A
1671-6647(2017)03-0404-10
10.3969/j.issn.1671-6647.2017.03.010
2016-05-03
山东省渤海海洋生态修复及能力建设项目——山东省渤海海洋保护区生物多样性数据库构建及生物保护方案制定(20140601)
喻 龙(1974-),男,江西南昌人,高级工程师,硕士,主要从事海洋生态环境方面研究.E-mail:yulong589@126.com
*通讯作者:郝彦菊(1980-),女,山东日照人,讲师,博士,主要从事海洋环境方面研究.E-mail:yanju_hao@126.com
