山西太岳山典型森林物种多样性和功能多样性关系研究
2017-08-09李晓强田平程小琴
李晓强,田平,程小琴*
(1.山西省林业种苗管理总站,山西 太原 030012; 2.北京林业大学 省部共建森林培育与保护教育部重点实验室,北京 100083)
山西太岳山典型森林物种多样性和功能多样性关系研究
李晓强1,田平2,程小琴2*
(1.山西省林业种苗管理总站,山西 太原 030012; 2.北京林业大学 省部共建森林培育与保护教育部重点实验室,北京 100083)
[目的]物种多样性(SD)和功能多样性(FD)在解释生态系统功能上发挥着越来越重要的作用。为探究不同群落层次(乔木、灌木、草本)SD-FD关系以及影响SD-FD的主要因子。[方法]对太岳山华北落叶松白桦混交林以及辽东栎次生林SD和FD进行研究。[结果]不同林分、不同群落层次间SD-FD关系存在差异,华北落叶松白桦混交林内乔木层中SD与FD表现为显著正相关关系,辽东栎次生林乔木层的SD与FD无显著相关关系,而2种林分的灌木层与草本层SD与FD均呈现显著负相关;SD-FD的关系取决于物种均匀度(Jw)、物种丰富度(S)以及种间性状差异(dij)对FD、SD的综合影响,当SD-FD正相关时,3个因子与FD/SD有显著正相关关系,并且dij的解释力最大,当SD-FD负相关时,Jw、S与FD/SD呈负相关关系,dij与FD/SD正相关,并且各因子解释力大小表现为Jw>S>dij。[结论]本文为森林生态系统多样性与稳定性研究以及森林经营管理提供了理论依据。
物种多样性; 功能多样性; 物种丰富度; 物种均匀度; 种间性状差异
随着功能生态学的发展,生态学家发现基于性状研究很多生态学问题可以得到更好地解释,尤其是物种多样性(species diversity,SD)与功能多样性(functional diversity,FD)的关系在解释物种共存机制、生物多样性对生态系统功能作用机制上发挥着重要作用[1,2]。从功能属性来看,群落内物种多样性包含功能多样性与功能冗余(functional redundancy)2部分[3~5],功能多样性表示物种间功能性状的相对差异性,对生态系统功能有较强解释力,尤其在多样性与生产力关系上,其解释力要高于物种多样性[6];而功能冗余表示物种间功能性状的相似性,对生态系统抵御干扰和干扰恢复能力有重要意义[4,5]。大量研究表明SD-FD关系并不是唯一确定的,可能表现为正相关、负相关以及无相关等,多数研究认为SD-FD关系主要取决与功能冗余程度,即种间性状差异[7],而陈超等人对高寒草甸研究结果表明物种均匀度是决定功能多样性的主要因子,也是导致SD-FD正相关的主要原因[2]。但以上研究主要围绕草原生态系统展开,并且主要通过剔除物种或者组建人工群落进行,对于结构较为复杂的森林生态系统研究相对较少。自然群落中的研究结果是否与受控实验一致有待进一步确认。
太岳林区是山西省八大林区之一,属太岳山主体部分,森林覆盖率为76.6%,人工林主要以华北落叶松人工林、油松人工林为主,天然林主要为天然栎类林[8]。本文通过对太岳山落叶松白桦混交林以及辽东栎次生林不同坡位下物种多样性和功能多样性研究,探讨不同林分物种多样性、功能多样性差异,不同植被类型SD-FD关系以及影响SD-FD的主要因子,为森林生态系统多样性研究以及森林经营管理提供一定理论依据。
1 研究地概况与研究方法
1.1 研究地概况
本研究以落叶松白桦混交林和辽东栎次生林为研究对象,研究地分别位于山西太岳山好地方林场和灵空山林场。好地方林场分布于111°59′E~122°01′E,36°44′N~36°47′N,海拔1749~2 431 m,平均海拔2 090 m,土壤类型主要为棕壤。该区位于暖温带,为半干旱大陆性季风气候,年平均气温6.2 ℃,年降雨量600 mm,降水主要集中在7-9月,无霜期120 d。主要植被类型为华北落叶松人工林、落叶松白桦混交林、灌丛、草甸。乔木主要树种为华北落叶松(Larixprincipis-rupprechtii)、白桦(Betulaplatyphylla)、黄花柳(Salixcaprea);灌木主要为黄刺玫(Rosaxanthina)、粘毛忍冬(Lonicerafargesii)、绣线菊(Spiraeasalicifolia)等;草本主要包括细叶薹草(Carexduriuscula)、小花风毛菊(Saussureaparviflora)、糙苏(Phlomisumbrosa)等。
灵空山林场分布在110°01′~112°08′E,36°33′~36°43′N。海拔分布范围为1 450~1 928 m,平均海拔为1 500 m,,土壤类型主要有棕壤、褐土。同属暖温带半干旱大陆性季风气候,年平均气温8 ℃,年降雨量600~700 mm,降水主要集中在7-9月,无霜期145 d。主要植被类型为油松林、辽东栎次生林、灌丛、草甸。乔木主要树种为油松(Pinustabuliformis)、辽东栎(Quercuswutaishanica)、山杨(Populusdavidiana)、华北落叶松;灌木主要为胡枝子(Lespedezabicolor)、绣线菊、山荆子(Malusbaccata)等;草本主要包括细叶薹草、东风菜(Doellingeriascaber)、山罗花(Melampyrumroseum)等(太岳林局介绍册)。样地信息详见表1。
1.2 研究方法
1.2.1 数据采集
2016年8月,在华北落叶松白桦混交林和和辽东栎次生林分别设立样地。每个林分设立9块20 m×20 m样地,分别位于同一坡面的不同坡位(上坡、中坡、下坡),即每个坡位设3个重复,重复样地间隔20 m。在样地四角及中心分别设置5 m×5 m的灌木层样地和1 m×1 m的草本层样地,进行基本群落调查。
样地内对优势种随机选择,进行性状调查并取样。其中乔木、灌木各取3株,记录其胸径(基径)、树高,草本取10株记录其株高。取样方法为:在冠层中部外侧随机选取健康、活力的枝叶,放在湿润的滤纸中带回驻地后立即进行称重,并用LI-3000C扫描叶面积,游标卡尺测量小枝长度以及小枝叶片数,并用排水法测小枝体积,然后将叶片和小枝放入80 ℃烘箱内烘干至恒重后取出称量干重。烘干后样品进行磨粉、过筛,进行化学计量分析(C、N、P、K),使用元素分析仪测定C、N含量,原子吸收光谱仪(火焰法)测定K含量,连续流动分析仪测定P含量。本文共选取叶性状、枝干性状和全株性状3个性状类型中的18个功能性状指标计算群落功能多样性。

表1 样地基本信息
1.2.2 相关计算公式
1)物种多样性SD以Simpson指数[7]计算
式中,Pi为第i个物种的重要值权重。
2)功能多样性FD采用Rao指数[9]进行计算
式中,Pi、Pj为第i、j个物种在群落中的相对多度;dij为物种i、j的欧氏距离(0≤dij≤1),即物种i、j的种间差异值
3)物种均匀度[10,11]计算公式为:
式中,S为样地内物种数目,即物种丰富度;Pi为物种i的重要值权重。
1.2.3 数据分析
使用excel 2010计算SD、FD、dij、Jw等指数,利用canoco 4.5软件(冗余分析)分析dij、Jw与S对SD-FD关系的影响。根据蒙特卡洛置换检验确定各个解释变量对SD-FD关系的影响大小以及影响显著性。在SigmaPlot 12.0进行制图。
2 结果与分析
2.1 样地内主要物种重要值
各样地内调查乔木的每木检尺,以及灌草层种名、株数、盖度等。根据调查结果,选取重要值大于0.1的优势物种进行取样。其中,华北落叶松白桦混交林共81个物种,共选取19个物种进行取样;辽东栎天然次生林共97个物种,共选取26种进行取样。各样地重要值大于0.1的物种见表2。
2.2 不同林分SD-FD关系
分别对华北落叶松白桦林与辽东栎次生林内不同群落层次(乔木、灌木、草本)的物种多样性与功能多样性进行线性回归(图1)。结果表明,不同群落层次的SD-FD关系存在差异。其中,华北落叶松白桦混交林内,乔木层SD-FD呈极显著正相关关系,草本层与灌木层的SD-FD均呈现显著负相关关系。辽东栎次生林内,功能多样性与物种多样性均呈现负相关关系,但在灌木、草本层中,两者之间呈现显著负相关关系,而乔木层中二者关系不显著。与华北落叶松白桦混交林相比,辽东栎次生林中,物种多样性与功能多样性相关性较低。
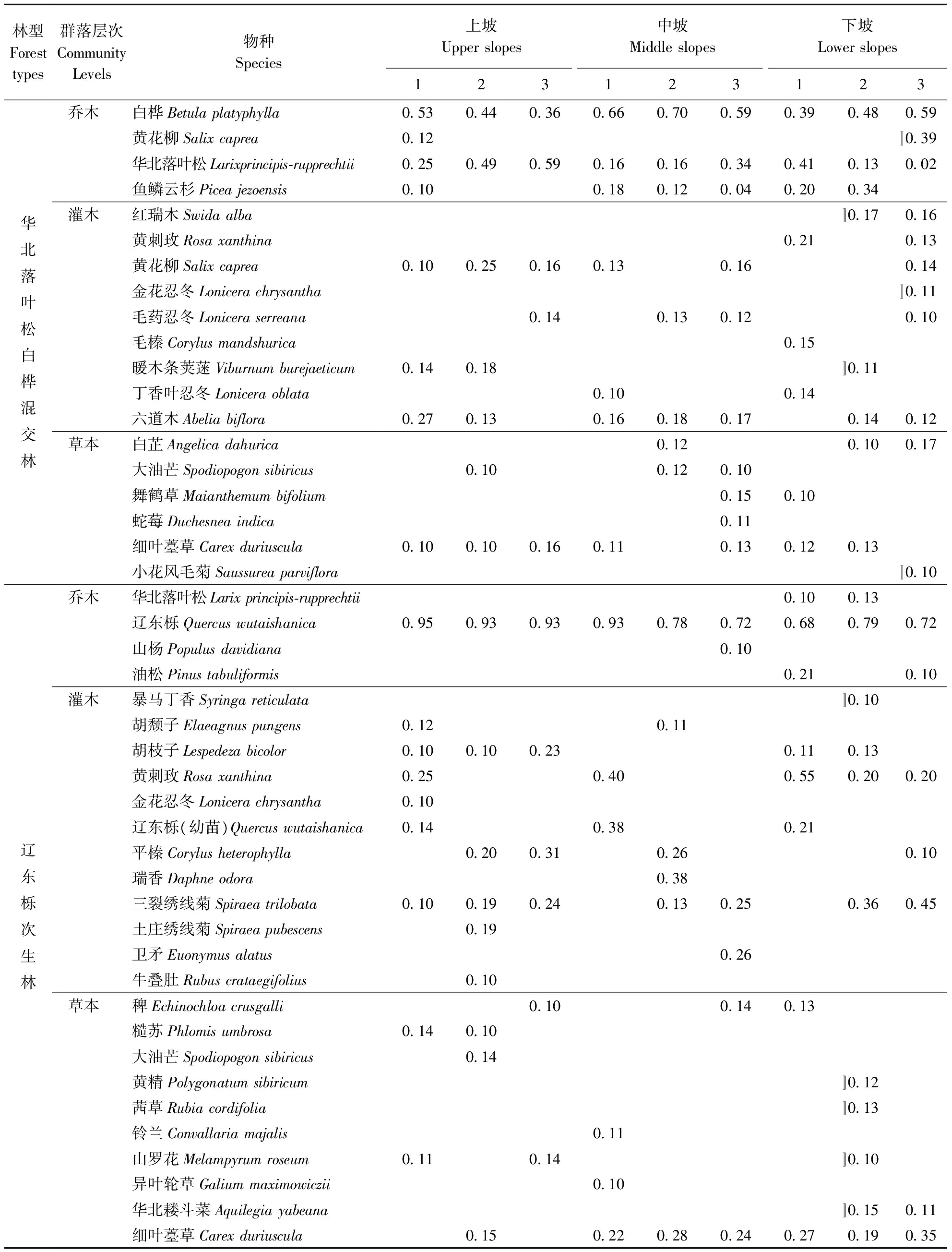
表2 各样地主要物种重要值
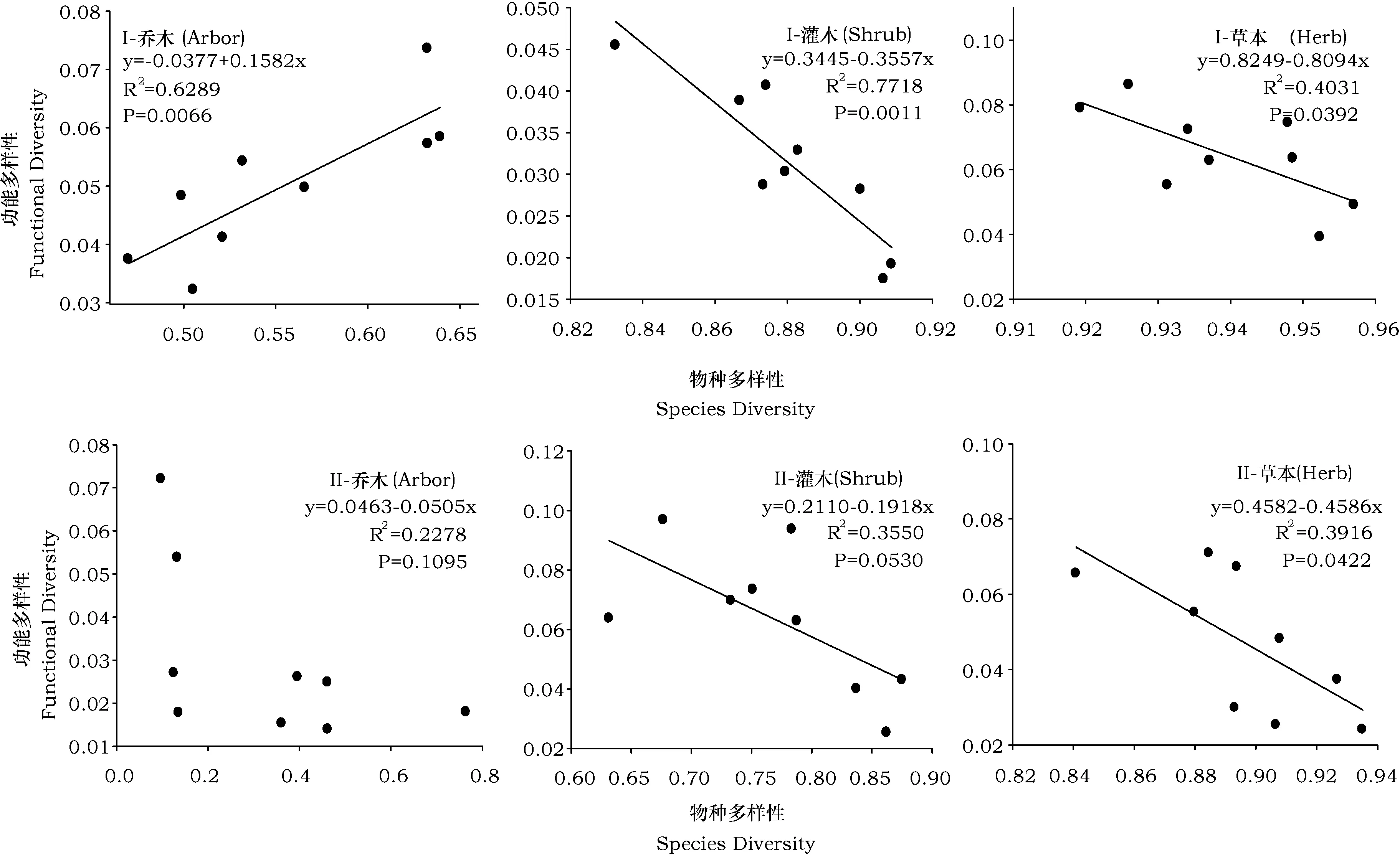
图1 不同群落层次物种多样性与功能多样性线性回归关系Fig.1 Linear regression relationship between species diversity and functional diversity of different community levelsI:华北落叶松白桦混交林(the Larch birch mixed forest); II:辽东栎次生林(the Q. wutaishanica forest)
2.3 物种均匀度、物种丰富度以及种间性状差异对SD-FD关系的影响
以物种均匀度(Jw)、物种丰富度(S)以及物种种间性状差异(dij)作为解释变量,以物种多样性与功能多样性的关系(功能多样性与物种多样性的比值,FD/SD)为响应变量进行RDA分析。结果显示(图2):Jw、S和dij3个因子对SD-FD关系的共同解释力较强,而单个因子的解释力大小与群落层次和林分类型有关。华北落叶松白桦混交林内,乔木层中dij对SD-FD关系解释力最高,能够解释34.2%的变化,Jw的解释力最低,只能解释7.4%的变化;灌木层中Jw和S对SD-FD关系的解释力明显高于dij,分别为54.9%和35.7%,dij的解释力非常微弱;草本层中,Jw、S、dij三个因子对SD-FD关系均有较强解释力,分别为53.4%、45.9%、35.2%。
辽东栎次生林内,乔木层的各因子对SD-FD关系均有较高解释力,其中,S的解释量高达41.7%;灌木层中,S、Jw、dij对SD-FD关系均有显著影响,且解释能力为Jw>S>dij;草本层中,Jw和S对SD-FD关系的解释力均显著高于dij,其中,Jw的解释能力最高,为34%,而dij的解释能力仅为0.5%。
3 结论与讨论
本研究显示,森林生态系统中,SD-FD关系与林分类型和群落层次(乔木、灌木、草本)密切相关。两种林分的灌木层与草本层SD与FD均呈现显著负相关,而华北落叶松白桦混交林内乔木层中SD与FD表现为显著正相关性关系,辽东栎次生林乔木层的SD与FD无显著相关关系。植物群落的构建理论认为,植物功能性状的构建模式一般包含种间性状趋同和性状趋异两种相反的过程[12,13],种间性状趋同受环境筛作用,导致群落物种的性状值最终趋向于达到与环境相适应的平均水平,从而引起生态位重叠,功能冗余增加,功能多样性减小[12,14];而种间性状趋异受种间竞争作用所驱动,导致生态位分化,生态位重叠减小,功能冗余减小,功能多样性增大[12,13]。本研究中,2种林分林下层植物(灌木、草本层)的SD-FD均呈显著负相关关系,说明SD的增加对FD的增加有负效应,群落物种所占总生态位减小。这说明,在2种林分中,这种关系林下环境的选择作用使物种的功能属性变化,表现出强烈趋同性,群落内有关的物种性状的组成也会被限制在适应此选择压力的范围内,因而物种丰富度的继续增加只会引起生态位的进一步分化且减小种间性状差异[7,15]。而在华北落叶松白桦混交林内乔木层中,种间竞争的趋异作用大于环境筛的趋同作用,SD的增加会导致种间竞争增强,从而导致群落整体生态位拓宽,FD增大。
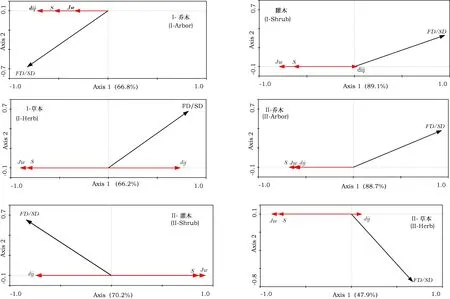
图2 各因子对SD-FD关系的解释力Fig.2 Explanation of each factor on the SD-FD relationshipI:华北落叶松白桦混交林(the Larch birch mixed forest); II:辽东栎次生林(the Q. wutaishanica forest)
Sasaki等人[7]认为若群落中每个物种的性状功能具有唯一性,SD-FD为正相关关系,当种间性状功能有所重叠时,二者表现为减速增加关系,即,SD-FD的关系主要由种间的性状差异决定。Bello等人[1,2,16]认为物种多样性降低释放出更多生态位,增大了种间性状差异,而功能多样性是否增大取决于物种均匀度的变化,但陈超等人[2]研究发现物种均匀度与种间性状差异均显著影响物种多样性和功能多样性,而非只对功能多样性起作用。从Simpson指数与Rao指数计算公式可以看出,SD-FD关系实际上受到物种丰富度、物种种间性状差异以及各物种相对多度共同影响,陈超等人[2]使用物种均匀度来代替各物种相对多度。本研究发现物种均匀度、物种丰富度以及物种种间性状差异对SD-FD关系(FD/SD,即线性回归的斜率)的解释能力较高,能够解释大部分FD/SD的变化。SD-FD的关系取决于各因子对物种多样性与功能多样性的相对影响力,当3个因子对功能多样性的综合影响大于对物种多样性影响时,SD-FD表现为正相关关系;反之,SD-FD表现为负相关关系。并且本研究中,SD-FD表现为负相关的群落层次中,3个因子的解释力均表现为Jw>S>dij,而SD-FD表现为正相关的群落层次中,dij的解释力最高。
辽东栎次生林乔木层的SD与FD无显著相关关系,但RDA分析结果显示,3个因子对物种多样性的综合影响大于对功能多样性的影响,按本研究研究结果,辽东栎次生林乔木层的SD-FD应表现为负相关关系,推测其原因可能为:(1)回归模型选择不当,由图2可以看出辽东栎次生林乔木层的SD-FD大体符合幂函数的分布规律,但目前为止的研究中,几乎未见SD-FD呈现幂分布的研究;(2)取样样本过少,这也是本试验的主要不足之处,尤其是在辽东栎次生林内,物种极分布不均匀,试验结果存在一定偶然性。本研究中,2种林分环境(密度、海拔等)存在较大差异,这可能是导致SD-FD关系在不同林分间相关性表现不一致的原因之一[16]。林分密度和海拔与林下光环境、土壤水分含量、土壤pH等密切相关[17],进而影响物种分布以及物种功能分布[15,16],其具体影响机制有待进一步研究。为进一步证实本文研究结果,需要对森林群落进行进一步研究,深入探讨环境因子对SD-FD关系的影响。
[1]Bello F D, Jan L, Sebastià M T. Variations in species and functional plant diversity along climatic and grazing gradients[J]. Ecography,2006,29(6):801-810.
[2]陈超.高寒草甸种间性状差异和物种均匀度对物种多样性与功能多样性关系的影响[J].生态学报,2016,36(3):661-674.
[3]Elmqvist T, Folke C, Nyström M,et al.Response diversity,ecosystem change,and resilience[J].Frontiers in Ecology & the Environment,2003,1(9):488-494.
[4]Bello F D, Lepš J, Lavorel S, et al. Importance of species abundance for assessment of trait composition: an example based on pollinator communities[J]. Community Ecology,2007,8(2):163-170.
[5]姚天华,朱志红,李英年,等.功能多样性和功能冗余对高寒草甸群落稳定性的影响[J].生态学报,2016,36(6):1547-1558.
[6]潘石玉,孔彬彬,姚天华,等.刈割和施肥对高寒草甸功能多样性与地上净初级生产力关系的影响[J].植物生态学报,2015,39(9):867-877.
[7]Sasaki T, Okubo S, Okayasu T, et al. Two-phase functional redundancy in plant communities along a grazing gradient in Mongolian rangelands.[J]. Ecology, 2009, 90(9):2598-2608.
[8]太岳林局概况[EB/OL].http://www.sxstylj.cn/about.php?id=1(2016.1.7)
[9]Pielou E C.Ecological Diversity[M].New York:John Wiley and sons lac.1975:1-165.
[10]Lepš J, Bello F D, Lavorel S, et al. Quantifying and interpreting functional diversity of natural communities: Practical considerations matter[C]The Japan Society of Mechanical Engineers, 2006:481-501.
[11]Whittaker R H. Evolution and measurement of species diversity[J].Taxon,1972,21(2/3):213-251.
[12]孔彬彬,卫欣华,杜家丽,等.刈割和施肥对高寒草甸物种多样性和功能多样性时间动态及其关系的影响[J].植物生态学报,2016,40(3):187-199.
[13]Pillar V D,Duarte L D S,Sosinski E E,et al.Discriminating trait-convergence and trait-divergence assembly patterns in ecological community gradients[J]. Journal of Vegetation Science,2009,20(2):334-348.
[14]Cornwell W K, Ackerly D D. Community assembly and shifts in plant trait distributions across an environmental gradient in coastal California[J]. Ecological Monographs,2009,79(79):109-126.
[15]李晓刚.模拟放牧和土壤资源获得性对高寒草甸物种多样性、功能多样性与初级生产力关系的影响[D].西安:陕西师范大学, 2012.
[16]Chesson P, Gebauer R L E, Schwinning S, et al. Resource pulses, species interactions, and diversity maintenance in arid and semi-arid environments[J].Oecologia,2004,141(2):236-53.
[17]田平,韩海荣,康峰峰,等.密度调整对太岳山华北落叶松人工林冠层结构及林下植被的影响[J].北京林业大学学报,2016,38(8):45-53.
(编辑:马荣博)
The species diversity and functional diversity of the typical forest of Taiyue Mountain Shanxi, China
Li Xiaoqiang1, Tian Ping2, Cheng Xiaoqin2*
(1.ShanxiAdministrationStationofForestrySeedlings,Taiyuan030012,China; 2.KeyLaboratoryofMinistryofForestCultivationandConservationofMinistryofEducation,BeijingForestryUniversity,Beijing100083,China)
[Objective]Species diversity (SD) and functional diversity (FD) play a more and more important role in explaining the ecosystem function. In order to identify which basic component of species diversity and functional diversity, such as species evenness (Jw), species richness (S) and trait dissimilarity among species (dij), contributes more to determine or alter theSD-FDrelationship. [Methods]We measured 17 functional traits of main species (the total important value >80%), and then calculated theSD,FD,Jw,S,dijof different community levels of two typical forests in Shanxi province. [Results]The result indicated that there were differences between the two forests and different community levels in theSD-FDrelationship, which was significant negative correlation in shrub layer and herb layer of both the two forests, while it was significant positive correlation in the arbor layer of the mixed forest and no significant correlation in theQuercuswutaishanicaforest. TheSD-FDrelationship was determined by the comprehensive influence of species evenness (Jw), species richness (S), trait dissimilarity among species (dij). The slope ofFDandSDwas significantly positively correlated with the three factors, in whichdijhad the largest explanatory, when the slope was greater than zero. Conversely, the slope had a significant negative correlation withJwandS, and a positive correlation withdijwhen it was less than zero, where the explanation of the three factors were shown asJw>S>dij. [Conclusion]This paper provides a theoretical basis for the study of the diversity and stability of forest ecosystems and forest management.
Pecies diversity, Functional diversity, Species evenness, Species richness, Trait dissimilarity among species
2017-05-15
2017-06-06
李晓强(1965-),男(汉),山西交口人,高级工程师,研究方向:林木种苗、森林培育
*通信作者:程小琴,讲师,Tel:13811206707;Email: cxq_200074@163.com
国家林业公益性行业科研专项(201404213)
S753.3
A
1671-8151(2017)09-0628-07
