Umbilical cord: an unlimited source of cells di ff erentiable towards dopaminergic neurons
2017-08-07MahdiEskandarianBoroujeniMossaGardaneh
Mahdi Eskandarian Boroujeni, Mossa Gardaneh
Department of Stem Cells and Regenerative Medicine, Faculty of Medical Biotechnology, National Institute of Genetic Engineering and Biotechnology, Tehran, Iran
Umbilical cord: an unlimited source of cells di ff erentiable towards dopaminergic neurons
Mahdi Eskandarian Boroujeni, Mossa Gardaneh*
Department of Stem Cells and Regenerative Medicine, Faculty of Medical Biotechnology, National Institute of Genetic Engineering and Biotechnology, Tehran, Iran
How to cite this article:Boroujeni ME, Gardaneh M (2017) Umbilical cord: an unlimited source of cells di ff erentiable towards dopaminergic neurons. Neural Regen Res 12(7):1186-1192.
Cell replacement therapy utilizing mesenchymal stem cells as its main resource holds great promise for ultimate treatment of human neurological disorders. Parkinson’s disease (PD) is a common, chronic neurodegenerative disorder hallmarked by localized degeneration of a speci fi c set of dopaminergic neurons within a midbrain sub-region.e speci fi c cell type and con fi ned location of degenerating neurons make cell replacement therapy ideal for PD treatment since it mainly requires replenishment of lost dopaminergic neurons with fresh and functional ones. Endogenous as well as exogenous cell sources have been identi fi ed as candidate targets for cell replacement therapy in PD. In this review, umbilical cord mesenchymal stem cells (UCMSCs) are discussed as they provide an inexpensive unlimited reservoir di ff erentiable towards functional dopaminergic neurons that potentially lead to long-lasting behavioral recovery in PD patients. We also present miRNAs-mediated neuronal di ff erentiation of UCMSCs.e UCMSCs bear a number of outstanding characteristics including their non-tumorigenic, low-immunogenic properties that make them ideal for cell replacement therapy purposes. Nevertheless, more investigations as well as controlled clinical trials are required to thoroughly con fi rm the e ffi cacy of UCMSCs for therapeutic medical-grade applications in PD.
nerve regeneration; umbilical cord; mesenchymal stem cells; di ff erentiation; neuronal; dopaminergic neurons; dopamine; substantia nigra; ventral mesencephalon; Parkinson’s disease; cell replacement therapy; neural regeneration
Accepted: 2017-05-27
Introduction
Cell replacement approaches for ultimate treatment of degenerative disorders by replenishing lost cells with fresh and functional ones largely rely on differentiation of potential cell sources.is is mainly because cell loss plays a central role in onset and progression of degenerative diseases (Fox et al., 2014).
Parkinson’s disease (PD) is a typical neurodegenerative disorder characterized by progressive but largely localized degeneration of dopaminergic (DAergic) neurons chiefly within midbrain substantianigra pars compacta (SNpc) (Hirsch et al., 1997). Current pharmacotherapeutic approaches include administration of dopamine precursors such as L-dihydroxy phenyl alanine, dopamine agonists, monoamine oxidase B inhibitors, catechol-O-methyltransferase inhibitors and adenosine A2A antagonists (Stocchi, 2014), blockage of dopamine D3 receptors (Cortés et al., 2016), inhibition of neuronal nitric oxide synthase (Maccallini and Amoroso, 2016) and use of small-molecule epigenetic modifiers (Hegarty et al., 2016). Moreover, other alternatives such as surgical options, mainly deep brain stimulation, are being used in patients with advanced PD (Odekerken et al., 2013). Nevertheless, these treatments solely provide symptomatic relief and are not able to hinder disease progression. In this review, the potential therapeutic application of umbilical cord mesenchymal stem cells (UCMSCs) in PD will be discussed with an exclusive focus on generation of DAergic neurons.
In Search of Cell Sources for PD Cell Replacement: Criteria
The proof-of-principle for feasibility of cell replacement therapy (CRT) in PD is provided by the fact that clinical transplantation of human fetal tissues (in the open-label trials) obtained from ventral mesencephalon results in striatal re-innervation of DAergic neurons and symptomatic relief (Lindvall and Björklund, 2004). Cell replacement therapy (CRT) is aimed at constitution of a neuroprotective and/ or neuroregenerative platform for human neurological disorders such as PD. However, prior to clinical translation of stem cells for PD, a number of requirements need to be taken to account in order to opt for a suitable cell type.ey include 1) a standardized, medical grade differentiation protocol with minimum reliance on various growth factors, chemicals and any animal components; 2) phenotypical resemblance of cell sources upon differentiation. In the case of PD, stem cell-derived DAergic neurons have to display DAergic phenotype, express DAergic markers such as Pitx3, Nurr1, Engrailed-1, Lmx1a, tyrosine hydroxylase, aromatic acid decarboxylase and vesicular monoamine transporter and release dopamine in a controlled fashion. They must also display electrophysiological characteristics of substantia nigra (SN) neurons; 3) long-term survival of graed DAergic neurons in the target tissue; 4) active integration into localneural network; target DAergic neurons should augment e ff ective reconstitution of neural circuits and integrate into host striatum; 5) functionality of engraed cells; functional DAergic neurons have to alleviate Parkinson’s symptoms and improve behavioral motor conditions after transplantation in a rodent PD model without causing any sign of tumorigenicity and dyskinesias (Brundin and Hagell, 2001; Isacson et al., 2003; Braak and Del Tredici, 2008). It is noteworthy that transplantation of differentiated cells into various sub-regions in the striatum where A8, A9 and A10 dopaminergic cell groups extend their fibers to release dopamine would increase the chance of recovery and movement restoration in animal models of PD. On the other hand, impurities carrying non-DAergic neurons in our cell reservoir could o ff er therapeutic advantages to pure sources for cell therapy because not just dopaminergic but also neuronal types of serotonergic and others are damaged in the course of PD.
In Search of Cell sources for PD Cell Replacement: Candidates
Cell sources are categorized based on their potentiality to undergo DAergic differentiation into endogenous and exogenous candidates. Endogenous candidates include primary cultures of ventral mesencephalon (VM) that consist of DAergic progenitor cells and neural stem cells from subventricular zone, striatum (ST) and SN (Fallon et al., 2000; Storch et al., 2004; Mohapel et al., 2005; Madhavan et al., 2009). Exogenous sources, on the other hand, include embryonic cells, induced pluripotent stem cells (iPSCs) and also mesenchymal stem cells derived from bone marrow, amniotic fl uid, sertoli cells, retinal pigment epithelium (RPE), adipocytes, carotid body cells, adrenal medullary, cervical sympathetic ganglion neurons and olfactory mucosal cells (Bankiewicz et al., 1988; Espejo et al., 1998; Subramanian et al., 2002; Nakao et al., 2004; Mc-Laughlin et al., 2006; Levy et al., 2008; Murrell et al., 2008; Glavaski-Joksimovic et al., 2009). These cell candidates have already been subjected to various treatments with inducing factors by adding these factors to cell medium and also by gene transfer to direct them towards DAergic fate. In addition, co-culture systems have been established by several helper cell lines including fi broblasts, pA6 stromal cell line, sertoli cells, RPE, as reviewed by Gardaneh (2010). They appear to secrete medium specific growth factors capable of inducing DAergic phenotype when interacting with the target cells. Furthermore, we previously proposed a combined, multi-factorial approach based on interaction of GPX-1-overexpressing DAergic neurons, GDNF secreting astrocytes and Nurr1-expressing microglia to potentiate survival and biological function of DAergic neurons against 6-OHDA toxicity (Gardaneh et al., 2010).
Mesenchymal Stem Cells (MSCs) versus Embryonic Stem Cells (ESCs)
Stem cells are regarded as undifferentiated cells that can undergo both proliferation and differentiation (Fuchs and Segre, 2000). ESCs are stem cells derived from the inner cell mass of the blastocysts (omson, 1998). MSCs are non-hematopoietic adult stem cells that possess the capacity to differentiate into various tissues including bone, cartilage and adipose tissue (Pountos and Giannoudis, 2005). MSCs can be isolated from bone marrow (Bianco et al., 2001), adipose tissue (Zuk et al., 2001), cord blood, amniotic fluid (In`t Anker, 2003) and placental tissue (Karahuseyinoglu et al., 2007).
MSCs have been described as plastic adherent multipotent cells represented by distinct terminologies such as “colony-forming fi broblastic cells” (Kuznetsov et al., 1997), “bone marrow stromal cells (BMSC)” (Peister, 2004), “multipotent adult progenitor cells” (Jiang et al., 2002) and “marrow isolated adult multi-lineage inducible cells” (D’Ippolito, 2004; Boroujeni et al., 2012). ESCs may appear as an appealing source for any cell-based therapy but their possible complications such as tumor formation, the need for immunosuppression, limited ESCs supply and above all, ethical concerns have substantially restricted their therapeutic use.erefore, the employment of MSCs in the tissue regeneration has attracted great interest as therapeutic agents. Moreover, these cells are capable of treating a variety of maladies including spinal cord injury (Hofstetter et al., 2002) and stroke (Chen et al., 2001), although UCMSC-derived dopaminergic neurons have not be utilized in the clinic.is means that steps have to be taken to clarify both beneficial and deleterious consequences of such a therapy for human patients.
The plasticity and transdifferentiation capacity of MSCs have provided an e ff ective platform as they di ff erentiate into other lineages of ectodermal and endodermal cells. Mezey et al. (2000) initially described thein vivodi ff erentiation of transplanted adult bone marrow cells into glial cells. To be utilized speci fi cally for PD cell therapy, studies have reported the feasibility of neuronal di ff erentiation of MSCs in which the paracrine e ff ect of the cells has been taken into account (Kitada and Dezawa, 2012).
Umbilical Cord: a Reservoir of MSCs
The umbilical cord consists of two umbilical arteries and also one umbilical vein which delivers oxygenated, nutrient-rich blood to the fetus (Meyer et al., 1978).is vascular structure is buried within a jelly-like tissue called umbilical cord matrix or Wharton’s jelly which is counted as the gelatinous connective tissue (Wang et al., 2004). These cells express MSC markers SH2 and SH3 but not CD35 and CD45 which are regarded as hematopoietic markers. In addition, they exhibit the capacity to differentiate into a wide range of lineages including adipocytes, osteocytes, chondrocytes, and neural lineages (Mitchell et al., 2003; Wei et al., 2012). UCMSCs have shown scores of advantages over other stem cell sources outlined below: 1) they exist in more primordial stages of differentiation than other mesenchymal cells including BMSCs (Hao et al., 1995). 2)ey do not express many of immunological markers involved in tissue rejection as shown by successful transplantation of umbilical cord blood nucleated cells in a 23-month-old child su ff eringfrom hemophagocytic lymphohistiocytosis (Schwinger et al., 1998). 3) Isolation, expansion, and freezing of these cells are easier and less expensive compared to many other sources such as neural stem cells (Taghizadeh et al., 2011; Dalous et al., 2012). 4)ey demonstrate high proliferation rate compared to BMSCs (Baksh et al., 2007; Boroujeni et al., 2012). 5)ey can be genetically manipulated to express various factors and/or used as delivery vehicles for therapeutic applications (Kim et al., 2008; Li et al., 2013; Zhang et al., 2014).
Dopaminergic Di ff erentiation of UCMSCs
Production of functional DAergic neurons relies fundamentally on signaling factors such as Shh, FGF8 and Wnt1 that initiate DAergic neurogenesis. Subsequently, the gene expression of LIM homeodomain family members (Lmx1a, Lmx1b) and FoxA2 facilitates specification of DAergic progenitors, which paves the way for terminal differentiation, promoted by cooperative function of Nurr1 and Pitx3 (Chakrabarty et al., 2012; Hegarty et al., 2013). In order to demystify the precise mechanisms of DAergic di ff erentiation in MSCs, early events parallel with late events need to be examined. Such studies will clarify the innate preparedness and potential of MSCs for neuronal/DAergic di ff erentiation. Reports indicate that UCMSCs are capable of displaying neuronal phenotype by expressing neuron-specific enolase (Mitchell et al., 2003), astrocytic marker GFAP and oligodendrocytic marker CNPase (Ha et al., 2004; Tracy et al., 2008).e UCMSCs can be induced to di ff erentiate into DAergic neurons comparatively to a high or low degree of success as shown in Table 1. These studies mainly applied transcription factors and/or growth factors as transgenes from within or as supplements from outside cells as inducing forces for neurogenesis and neuroprotection. Inclusion of exogenous genes in target cells that are destined to be part of patients’live tissue can be a source of biological and ethical concerns. It is not possible to readily predict the physiological consequences of exogenous gene expression, neither are scientists prepared to fully satisfy the society of unforeseeable complications inherent with transgene transfer into human body. Further, undi ff erentiated UCMSCs were used for transplantation and mostly resulted in behavioral recovery in animal models of PD based on parkinsonian toxins 6-hydroxydopamine (6-OHDA), 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine (MPTP) or rotenone. For instance, Weiss et al. (2006) and Xiong et al. (2010) merely used undi ff erentiated UCMSCs without applying any growth factors for transplantation and made promising observations for decreased rotations by 50% and 67% respectively. Other investigators employed several inducing factors including epidermal growth factor (EGF), sonic hedgehog (SHH), nerve growth factor (NGF) and/or speci fi c conditioned media (e.g., from amniotic epithelial cells, AEC) to facilitate DAergic neuron production and, upon PD brain transplantation, observed alleviation of motor symptoms (Fu et al., 2006; Li et al., 2010; Kang et al., 2013; Shetty et al., 2013; Yang et al., 2013a; Zhao et al., 2016). Our laboratory has recently tested the inducing e ff ect of murine cerebrospinal fl uid on DAergic di ff erentiation of UCMSCs which were then transplanted to rat striatum (Aliaghaei et al., 2016). The cells survived and resulted in rotational recovery and reduced rate of neuronal apoptosis in animal’s injured brain. Besides, we successfully generated MSCs-de-rived DAergic secreting neurons using a cocktail of choroid plexus epithelial cells-conditioned medium and knockout serum replacement (Boroujeni et al., 2017). A combinatorial approach to convert UCMSCs to DAergic neurons employed both transgenes (such as Lmx1a and neurturin, NTN) and external inducing factors (basic fibroblast growth factor, β-mercaptoethanol, SHH, fi broblast growth factor-8, retinoic acid) that upon cell transplantation demonstrated improvements in behavioral de fi cits against MPTP (Yan et al., 2013). Measures outlined above could pave the ground for survival as well as regeneration of host neurons and ultimately for augmented e ffi cacy of repairing target neural tissues. In accordance with this notion, MSCs transplanted into the brain stimulate endogenous neuronal growth and synaptogenesis and enhance functional recovery of damaged neurons (Lin et al., 2011; Olson et al., 2012).ese trophic e ff ects are implemented by MSC-mediated secretion of a variety of cytokines and neurotrophic factors (Joyce et al., 2010). Besides, the analysis of UCMSCs conditioned medium secretome revealed it contains neuro-regulatory components [such as Cyr61 protein and colony-stimulating factor 1 (CSF-1)] essential for the maintenance and maturation of neural stem cells, respectively (Pires et al., 2016). Hence, it is important in these studies to distinguish between neurogenic and paracrine protective e ff ects of the administered UCMSCs so their fate as well as their ultimate therapeutic e ff ects can be clearly determined.
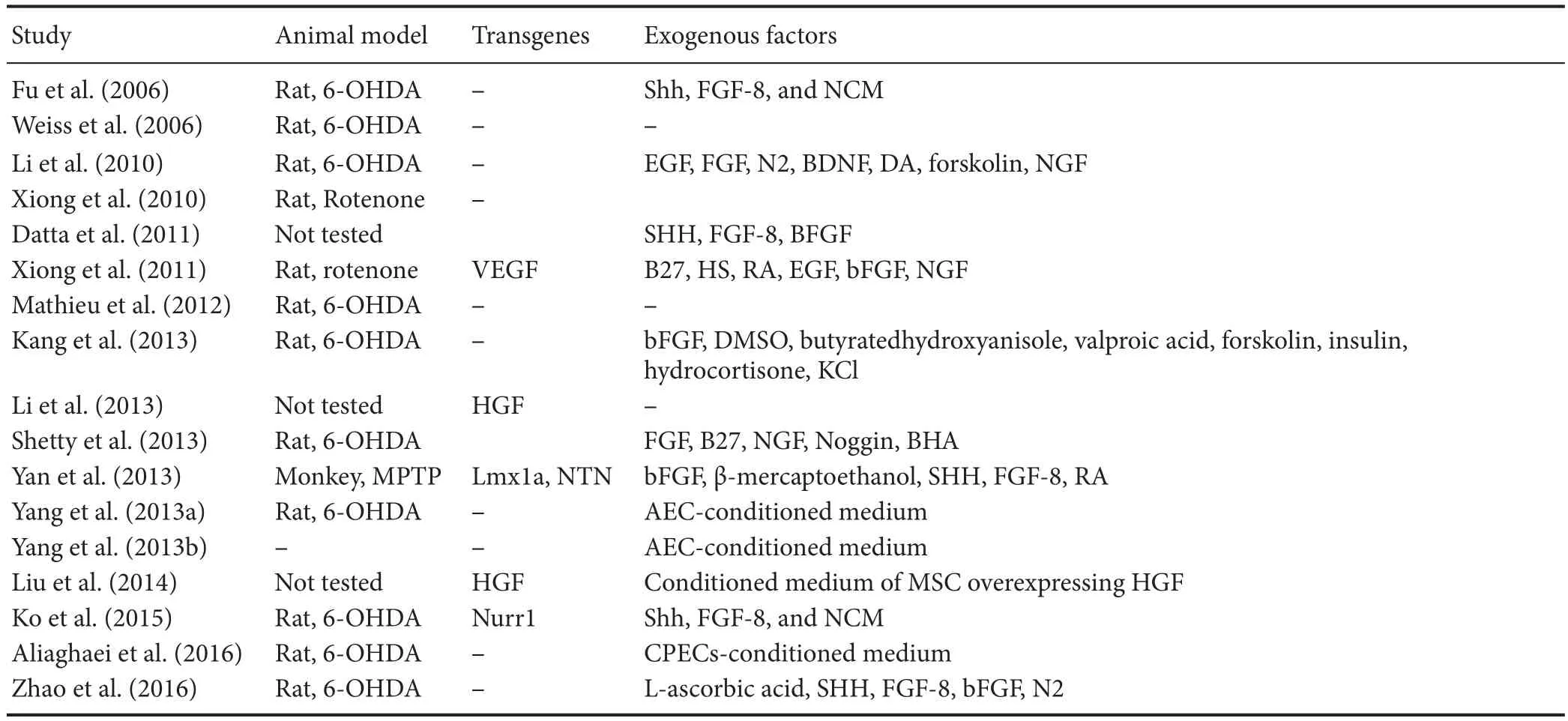
Table 1 Umbilical cord mesenchymal stem cells-based therapy forin vitrogeneration of dopaminergic neurons
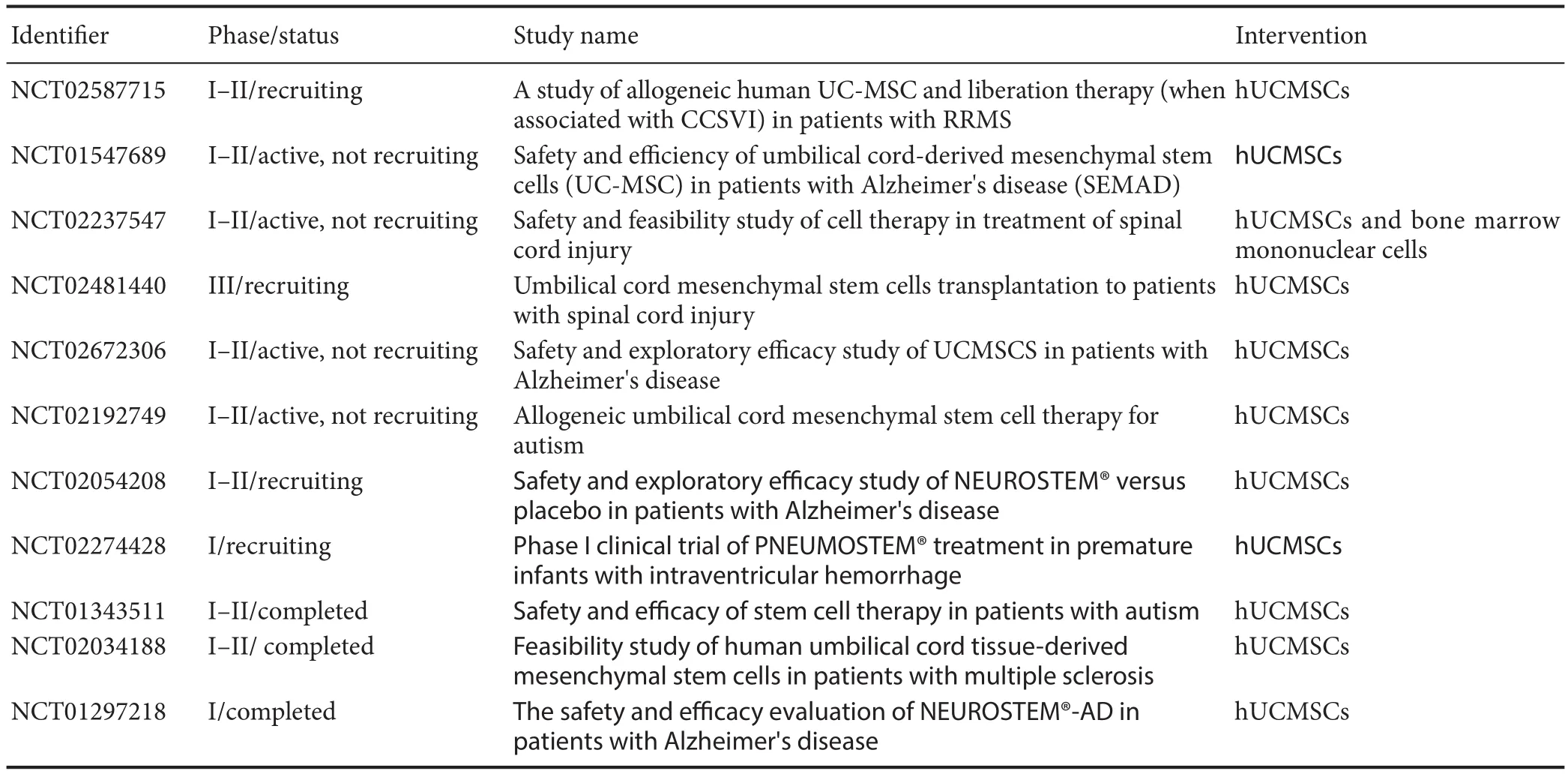
Table 2 Current trials using human umbilical cord mesenchymal stem cells (UCMSCs) for neurological disorders (retrieved from ClinicalTrials.gov, April 2017)
MiRNAs-Mediated Neuronal Di ff erentiation of UCMSCs
In order to develop UCMSCs-based therapeutic strategies, a comprehensive understanding of signaling pathways involved in proliferation and differentiation of UCMSCs is needed. These biological processes are considerably modulated by genetic and epigenetic mechanisms. Recently, microRNAs (miRNAs) as small non-coding RNA have been demonstrated to play essential roles in a plethora of biological functions including neural differentiation and neurodegeneration (Christensen and Schratt, 2009). Extensive studies have been carried out and shed light on the gene expression pro fi le of miRNAs involved particularly in neurogenesis. For instance, miR-124a and miR-9 have been regarded as most specific and well-studied brain miRNAs which promote neural di ff erentiation (Liu and Zhao, 2009). Four miRNAs (miR-1290, miR-26b, miR-194, and miR-124a) have been recently shown to be up-regulated during neuronal di ff erentiation of UCMSCs.is study also disclosed two down-regulated miRNAs (miR-4521 and miR-543) with unknown functions in neurogenesis (Zhuang et al., 2015). Moreover, Meng et al. (2016) compared the gene expression pro fi le of miRNAs in umbilical cord and cord blood-derived MSCs and interestingly demonstrated that the expression of genes related to neurogenesis was increased in UCMSCs, which is contrary to the finding performed by Secco et al. (2009). This dissimilarity demands a detailed investigation into miRNA signature of tissue-speci fi c MSCs prior to any clinical application.
As mentioned above, miRNAs have critical roles in cellular processes. Autophagy is a self-degradation process of cytosolic components, in which defective cellular constituents are degraded through delivery to lysosomes. A recent study has suggested the signi fi cant implication of autophagic fl ux in neural differentiation (Vessoni et al., 2011). Indeed, genetic ablation of Atg5 and Atg7, both involved in autophagic machinery, in the mouse brain has been shown to lead toneurodegenerative disorders (Nikoletopoulou et al., 2015). Further, emerging evidence indicates a strong link between miRNA regulatory networks and autophagy pathways (Frankel and Lund, 2012). Thus, miRNA-autophagy interactions during neuronal di ff erentiation of UCMSCs should be well elucidated to improve the therapeutic potential of UCMSCs.
Conclusion
Pharmacotherapeutic options (including Mitoquinone, PYM50028 and Rasagiline) hitherto failed to improve PD patients in recent clinical trials, despite their great promise in experimental researches (Athauda and Foltynie, 2015). Likewise, Levodopa administration to restore dopamine levels causes dyskinesia few years aer treatment initiates (Prashanth et al., 2011). Moreover, the new drug development process is very costly and time-consuming (Sherer et al., 2012). MSCs-based approaches have opened a promising avenue into treatment of patients with neurological disorders such as PD. In this review, the potential therapeutic application of UCMSCs in PD patients was outlined. As discussed above, due mainly to their non-tumorigenic as well as low immunogenic capacity, UCMSCs could be considered as suitable cell candidates for neuro-regeneration and neuro-repair in treating neurodegenerative diseases with speci fi c focus on PD. Above and beyond, the release of dopamine by UCMSCs has to be under control by measures such as use of regulating genetic elements. However, clinical trials using UCMSCs for PD have not been registered in ClinicalTrials.gov (in April 2017) whereas for other brain diseases, studies are in progress as shown in Table 2. The completed studies are basically indicating that UCMSCs are stable and safe. Considering collected data from clinical trial failures related to neuroprotective drugs, enriched comprehension of mechanisms underlying PD is highly demanded if CRT is being employed. Additionally, the detailed outcomes of these trials will surely assist in setting up improved methods of cell preparation, purification and DAergic neuron enrichment, pre-injection cell manipulation, transplantation procedures, cell integration to neural network and their paracrine e ff ects beside long-term potential recovery from PD symptoms.
Acknowledgments:
Author contributions:Both of these two authors conceived, designed, wrote and approved the fi nal version of this paper.
Con fl icts of interest:None declared.
Open access statement:
Contributor agreement:A statement of “Publishing Agreement” has been signed by an authorized author on behalf of all authors prior to publication.
Plagiarism check:This paper has been checked twice with duplication-checking soware ienticate.
Peer review:A double-blind and stringent peer review process has been performed to ensure the integrity, quality and signi fi cance of this paper.
Aliaghaei A, Gardaneh M, Maghsoudi N, Salehinejad P, Gharib E (2016) Dopaminergic induction of umbilical cord mesenchymal stem cells by conditioned medium of choroid plexus epithelial cells reduces apomorphine-induced rotation in Parkinsonian rats. Arch Iran Med 19:561-570.
Athauda D, Foltynie T (2015)e ongoing pursuit of neuroprotective therapies in Parkinson disease. Nat Rev Neurol 11:25-40.
Baksh D, Yao R, Tuan RS (2007) Comparison of proliferative and multilineage di ff erentiation potential of human mesenchymal stem cells derived from umbilical cord and bone marrow. Stem Cells 25:1384-1392.
Bankiewicz KS, Plunkett RJ, Kophin IJ, Jacobowitz DM, London WT, Oldfield EH (1988) Transient behavioral recovery in hemiparkinsonian primates after adrenal medullary allografts. Prog Brain Res 78:543-549.
Bianco P, Riminucci M, Gronthos S, Robey PG (2001) Bone marrow stromal stem cells: nature, biology, and potential applications. Stem cells 19:180-192.
Boroujeni ME, Gowda P, Johnson J, Rao J, Saremy S (2012)e proliferation and di ff erentiation capacity of bone marrow derived human mesenchymal stem cells in early and late doubling. Asian J Biochem 7:27-36.
Boroujeni ME, Gardaneh M, Shahriari MH, Aliaghaei A, Hasani S (2017) Synergy between choroid plexus epithelial cells-conditioned medium and knockout serum replacement converts human adipose-derived stem cells to dopamine-secreting neurons. Rejuvenation Res doi: 10.1089/rej.2016.1887.
Braak H, Del Tredici K (2008) Assessing fetal nerve cell gras in Parkinson’s disease. Nat Med 14:483-485.
Brundin P, Hagell P (2001)e neurobiology of cell transplantation in Parkinson’s disease. Clin Neurosci Res 1:507-520.
Chakrabarty K, Von Oerthel L, Hellemons A, Clotman F, Espana A, Koerkamp MG, Holstege FC, Pasterkamp RJ, Smidt MP (2012) Genome wide expression pro fi ling of the mesodiencephalic region identi fi es novel factors involved in early and late dopaminergic development. Biol Open 1:693-704.
Chen J, Sanberg PR, Li Y, Wang L, Lu M, Willing AE, Sanchez-Ramos J, Chopp M (2001) Intravenous administration of human umbilical cord blood reduces behavioral deficits after stroke in rats. Stroke 32:2682-2688.
Christensen M, Schratt GM (2009) MicroRNA involvement in developmental and functional aspects of the nervous system and in neurological diseases. Neurosci Lett 466:55-62.
Cortés A, Moreno E, Rodríguez-Ruiz M, Canela EI, Casadó V (2016) Targeting the dopamine D3 receptor: an overview of drug design strategies. Expert Opin Drug Discov 11:641-664.
D’Ippolito G, Diabira S, Howard GA, Menei P, Roos BA, Schiller PC (2004) Marrow-isolated adult multilineage inducible (MIAMI) cells, a unique population of postnatal young and old human cells with extensive expansion and di ff erentiation potential. J Cell Sci 117:2971-2981.
Dalous J, Larghero J, Baud O (2012) Transplantation of umbilical cord-derived mesenchymal stem cells as a novel strategy to protect the central nervous system: technical aspects, preclinical studies, and clinical perspectives. Pediatr Res 71:482-490.
Datta I, Mishra S, Mohanty L, Pulikkot S, Joshi PG (2011) Neuronal plasticity of human Wharton’s jelly mesenchymal stromal cells to the dopaminergic cell type compared with human bone marrow mesenchymal stromal cells. Cytotherapy 13:918-932.
Espejo EF, Montoro RJ, Armengol JA, López-Barneo J (1998) Cellular and functional recovery of Parkinsonian rats aer intrastriatal transplantation of carotid body cell aggregates. Neuron 20:197-206.
Fallon J, Reid S, Kinyamu R, Opole I, Opole R, Baratta J, Korc M, Endo TL, Duong A, Nguyen G, Karkehabadhi M, Twardzik D, Patel S, Loughlin S (2000) In vivo induction of massive proliferation, directed migration, and di ff erentiation of neural cells in the adult mammalian brain. Proc Natl Acad Sci U S A 97:14686-14691.
Fox IJ, Daley GQ, Goldman SA, Huard J, Kamp TJ, Trucco M (2014) Use of di ff erentiated pluripotent stem cells in replacement therapy for treating disease. Science 345.
Frankel LB, Lund AH (2012) MicroRNA regulation of autophagy. Carcinogenesis 33:2018-2025.
Fu YS, Cheng YC, Lin MYA, Cheng H, Chu PM, Chou SC, Shih YH, Ko MH, Sung MS (2006) Conversion of human umbilical cord mesenchymal stem cells in Wharton’s jelly to dopaminergic neurons in vitro: potential therapeutic application for Parkinsonism. Stem Cells 24:115-124.
Fuchs E, Segre JA (2000) Stem Cells. Cell 100:143-155.
Gardaneh M (2010) Dopamine-synthesizing neurons: An overview of their development and application for cell therapy. Iran J Biotechnol 8:8.
Gardaneh M, Panahi Y, Shojaei S, Mazaheri-Tehrani E, Maghsudi N (2010) Neuroprotection in Parkinson’s disease: a multi-directional genetic strategy for maximum protection of dopaminergic neurons against Parkinsonian toxicity. J Med Hyp Ideas 4:7.
Glavaski-Joksimovic A, Virag T, Chang QA, West NC, Mangatu TA, McGrogan MP, Dugich-Djordjevic M, Bohn MC (2009) Reversal of dopaminergic degeneration in a Parkinsonian rat following micrografting of human bone marrow-derived neural progenitors. Cell Transplant 18:801-814.
Ha Y, Yoon SH, Park HC, Kim KN, Yoon DH, Cho YE (2004) Transplantation of human umbilical cord blood improves neurological outcomes in the rats after traumatic spinal cord injury. J Korean Neurosurg Soc 35:302-308.
Hegarty SV, Sullivan AM, O’Kee ff e GW (2013) Midbrain dopaminergic neurons: a review of the molecular circuitry that regulates their development. Dev Biol 379:123-138.
Hegarty SV, Sullivan AM, O’Keeffe GW (2016) The Epigenome as a therapeutic target for Parkinson’s disease. Neural Regen Res 11:1735.
Hirsch EC, Faucheux B, Damier P, Mouatt-Prigent A, Agid Y (1997) Neuronal vulnerability in Parkinson’s disease. In: Advances in Research on Neurodegeneration, pp 79-88. Springer Science + Business Media.
Hofstetter CP, Schwarz EJ, Hess D, Widenfalk J, El Manira A, Prockop DJ, Olson L (2002) Marrow stromal cells form guiding strands in the injured spinal cord and promote recovery. Proc Natl Acad Sci 99:2199-2204.
In`t Anker PS, Scherjon SA, Kleijburg-van der Keur C, Noort WA, Claas FHJ, Willemze R, Fibbe WE, Kanhai HHH (2003) Amniotic fluid as a novel source of mesenchymal stem cells for therapeutic transplantation. Blood 102:1548.
Isacson O, Bjorklund LM, Schumacher JM (2003) Toward full restoration of synaptic and terminal function of the dopaminergic system in Parkinson’s disease by stem cells. Ann Neurol 53:S135-S148.
Jiang Y, Jahagirdar BN, Reinhardt RL, Schwartz RE, Keene CD, Ortiz-Gonzalez XR, Reyes M, Lenvik T, Lund T, Blackstad M, Du J, Aldrich S, Lisberg A, Low WC, Largaespada DA, Verfaillie CM (2002) Pluripotency of mesenchymal stem cells derived from adult marrow. Nature 418:41-49.
Joyce N, Annett G, Wirthlin L, Olson S, Bauer G, Nolta JA (2010) Mesenchymal stem cells for the treatment of neurodegenerative disease. Regen Med 5:933-946.
Kang EJ, Lee YH, Kim MJ, Lee YM, Mohana Kumar B, Jeon BG, Ock SA, Kim HJ, Rho GJ (2013) Transplantation of porcine umbilical cord matrix mesenchymal stem cells in a mouse model of Parkinson’s disease. J Tissue Eng Regen Med 7:169-182.
Karahuseyinoglu S, Cinar O, Kilic E, Kara F, Akay GG, Demiralp DÖ, Tukun A, Uckan D, Can A (2007) Biology of stem cells in human umbilical cord stroma: in situ and in vitro surveys. Stem Cells 25:319-331.
Kim SM, Lim JY, Park SI, Jeong CH, Oh JH, Jeong M, Oh W, Park SH, Sung YC, Jeun SS (2008) Gene therapy using TRAIL-secreting human umbilical cord blood–derived mesenchymal stem cells against intracranial glioma. Cancer Res 68:9614-9623.
Kitada M, Dezawa M (2012) Parkinson’s disease and mesenchymal stem cells: potential for cell-based therapy. Parkinsons Dis 2012:1-9.
Ko TL, Fu YY, Shih YH, Lin YH, Ko MH, Fu TW, Lin TY, Hsiao HS, Chu PM, Fu YS (2015) A high e ffi ciency induction of dopaminergic cells from human umbilical mesenchymal stem cells for the treatment of hemiparkinsonian rats. Cell Transplant 24:2251-2262.
Kuznetsov SA, Friedenstein AJ, Gehron Robey P (1997) Factors required for bone marrow stromal fi broblast colony formation in vitro. Br J Haematol 97:561-570.
Levy YS, Bahat-Stroomza M, Barzilay R, Burshtein A, Bulvik S, Barhum Y, Panet H, Melamed E, O ff en D (2008) Regenerative e ff ect of neural-induced human mesenchymal stromal cells in rat models of Parkinson’s disease. Cytotherapy 10:340-352.
Li JF, Yin HL, Shuboy A, Duan HF, Lou JY, Li J, Wang HW, Wang YL (2013) Di ff erentiation of hUC-MSC into dopaminergic-like cells aer transduction with hepatocyte growth factor. Mol Cell Biochem 381:183-190.
Li M, Zhang SZ, Guo YW, Cai YQ, Yan ZJ, Zou Z, Jiang XD, Ke YQ, He XY, Jin ZL (2010) Human umbilical vein-derived dopaminergic-like cell transplantation with nerve growth factor ameliorates motor dysfunction in a rat model of Parkinson’s disease. Neurochem Res 35:1522-1529.
Lin YT, Chern Y, Shen CK, Wen HL, Chang YC, Li H, Cheng TH, Hsieh-Li HM (2011) Human mesenchymal stem cells prolong survival and ameliorate motor de fi cit through trophic support in Huntington’s disease mouse models. PLoS One 6:e22924.
Lindvall O, Björklund A (2004) Cell therapy in Parkinson’s disease. Neurotherapeutics 1:382-393.
Liu C, Zhao X (2009) MicroRNAs in adult and embryonic neurogenesis. Neuromolecular Med 11:141-152.
Liu XS, Li JF, Wang SS, Wang YT, Zhang YZ, Yin HL, Geng S, Gong HC, Han B, Wang YL (2014) Human umbilical cord mesenchymal stem cells infected with adenovirus expressing HGF promote regeneration of damaged neuron cells in a Parkinson’s disease model. Biomed Res Int 2014:909657.
Maccallini C, Amoroso R (2016) Targeting neuronal nitric oxide synthase as a valuable strategy for the therapy of neurological disorders. Neural Regen Res 11:1731-1734.
Madhavan L, Daley BF, Paumier KL, Collier TJ (2009) Transplantation of subventricular zone neural precursors induces an endogenous precursor cell response in a rat model of Parkinson’s disease. J Comp Neurol 515:102-115.
Mathieu P, Roca V, Gamba C, del Pozo A, Pitossi F (2012) Neuroprotective e ff ects of human umbilical cord mesenchymal stromal cells in an immunocompetent animal model of Parkinson’s disease. J Neuroimmunol 246:43-50.
McLaughlin D, Tsirimonaki E, Vallianatos G, Sakellaridis N, Chatzistamatiou T, Stavropoulos-Gioka C, Tsezou A, Messinis I, Mangoura D (2006) Stable expression of a neuronal dopaminergic progenitor phenotype in cell lines derived from human amniotic fluid cells. J Neurosci Res 83:1190-1200.
Meng X, Sun B, Xue M, Xu P, Hu F, Xiao Z (2016) Comparative analysis of microRNA expression in human mesenchymal stem cells from umbilical cord and cord blood. Genomics 107:124-131.
Meyer W, Rumpelt H, Yao A, Lind J (1978) Structure and closure mechanism of the human umbilical artery. Eur J Pediatr 128:247-259.
Mezey E, Chandross KJ, Harta G, Maki RA, McKercher SR (2000) Turning blood into brain: cells bearing neuronal antigens generated in vivo from bone marrow. Science 290:1779-1782.
Mitchell KE, Weiss ML, Mitchell BM, Martin P, Davis D, Morales L, Helwig B, Beerenstrauch M, Abou-Easa K, Hildreth T (2003) Matrix cells from Wharton’s jelly form neurons and glia. Stem cells 21:50-60.
Mohapel P, Frielingsdorf H, Häggblad J, Zachrisson O, Brundin P (2005) Platelet-derived growth factor (PDGF-BB) and brain-derived neurotrophic factor (BDNF) induce striatal neurogenesis in adult rats with 6-hydroxydopamine lesions. Neuroscience 132:767-776.
Murrell W, Wetzig A, Donnellan M, Féron F, Burne T, Meedeniya A, Kesby J, Bianco J, Perry C, Silburn P (2008) Olfactory mucosa is a potential source for autologous stem cell therapy for Parkinson’s disease. Stem Cells 26:2183-2192.
Nakao N, Shintani-Mizushima A, Kakishita K, Itakura T (2004) The ability of graed human sympathetic neurons to synthesize and store dopamine: a potential mechanism for the clinical effect of sympathetic neuron autografts in patients with Parkinson’s disease. Exp Neurol 188:65-73.
Nikoletopoulou V, Papandreou M, Tavernarakis N (2015) Autophagy in the physiology and pathology of the central nervous system. Cell Death Di ff er 22:398-407.
Odekerken VJ, van Laar T, Staal MJ, Mosch A, Ho ff mann CF, Nijssen PC, Beute GN, van Vugt JP, Lenders MW, Contarino MF (2013) Subthalamic nucleus versus globus pallidus bilateral deep brain stimulation for advanced Parkinson’s disease (NSTAPS study): a randomised controlled trial. Lancet Neurol 12:37-44.
Olson SD, Pollock K, Kambal A, Cary W, Mitchell GM, Tempkin J, Stewart H, McGee J, Bauer G, Kim HS (2012) Genetically engineered mesenchymal stem cells as a proposed therapeutic for Huntington’s disease. Mol Neurobiol 45:87-98.
Peister A (2004) Adult stem cells from bone marrow (MSCs) isolated from di ff erent strains of inbred mice vary in surface epitopes, rates of proliferation, and di ff erentiation potential. Blood 103:1662-1668.
Pires AO, Mendes-Pinheiro B, Teixeira FG, Anjo SI, Ribeiro-Samy S, Gomes ED1, Serra SC, Silva NA, Manadas B, Sousa N, Salgado AJ (2016) Unveiling the di ff erences of secretome of human bone marrow mesenchymal stem cells, adipose tissue-derived stem cells, and human umbilical cord perivascular cells: a proteomic analysis. Stem Cells Dev 25:1073-1083.
Pountos I, Giannoudis PV (2005) Biology of mesenchymal stem cells. Injury 36:S8-12.
Prashanth L, Fox S, Meissner WG (2011) l-Dopa-induced dyskinesia-clinical presentation, genetics, and treatment. Int Rev Neurobiol 98:31-54.
Schwinger W, Urban C, Lackner H, Benesch M, Kerbl R, Dornbusch H, Sovinz P, Kogler G (1998) CASE REPORTS-Unrelated 5/6-locus matched umbilical cord blood transplantation in a 23-monthold child with hemophagocytic lymphohistiocytosis. Bone Marrow Transplant 22:393-396.
Secco M, Moreira YB, Zucconi E, Vieira NM, Jazedje T, Muotri AR, Okamoto OK, Verjovski-Almeida S, Zatz M (2009) Gene expression pro fi le of mesenchymal stem cells from paired umbilical cord units: cord is di ff erent from blood. Stem Cell Rev 5:387-401.
Sherer TB, Chowdhury S, Peabody K, Brooks DW (2012) Overcoming obstacles in Parkinson’s disease. Mov Disord 27:1606-1611.
Storch A, Sabolek M, Milosevic J, Schwarz SC, Schwarz J (2004) Midbrain-derived neural stem cells: from basic science to therapeutic approaches. Cell Tissue Res 318:15-22.
Subramanian T, Marchionini D, Potter EM, Cornfeldt ML (2002) Striatal xenotransplantation of human retinal pigment epithelial cells attached to microcarriers in hemiparkinsonian rats ameliorates behavioral de fi cits without provoking a host immune response. Cell Transplant 11:207-214.
Taghizadeh R, Cetrulo K, Cetrulo C (2011) Wharton’s Jelly stem cells: future clinical applications. Placenta 32:S311-315.
Thomson JA (1998) Embryonic stem cell lines derived from human blastocysts. Science 282:1145-1147.
Tracy E, Aldrink J, Panosian J, Beam D,acker J, Reese M, Kurtzberg J (2008) Isolation of oligodendrocyte-like cells from human umbilical cord blood. Cytotherapy 10:518-525.
Vessoni AT, Muotri AR, Okamoto OK (2011) Autophagy in stem cell maintenance and di ff erentiation. Stem Cells Dev 21:513-520.
Wang HS, Hung SC, Peng ST, Huang CC, Wei HM, Guo YJ, Fu YS, Lai MC, Chen CC (2004) Mesenchymal stem cells in the Wharton’s jelly of the human umbilical cord. Stem cells 22:1330-1337.
Wei X, Peng G, Zheng S, Wu X (2012) Di ff erentiation of umbilical cord mesenchymal stem cells into steroidogenic cells in comparison to bone marrow mesenchymal stem cells. Cell Prolif 45:101-110.
Weiss ML, Medicetty S, Bledsoe AR, Rachakatla RS, Choi M, Merchav S, Luo Y, Rao MS, Velagaleti G, Troyer D (2006) Human umbilical cord matrix stem cells: preliminary characterization and effect of transplantation in a rodent model of Parkinson’s disease. Stem cells 24:781-792.
Xiong N, Cao X, Zhang Z, Huang J, Chen C, Zhang Z, Jia M, Xiong J, Liang Z, Sun S (2010) Long-term e ffi cacy and safety of human umbilical cord mesenchymal stromal cells in rotenone-induced hemiparkinsonian rats. Biol Blood Marrow Transplant 16:1519-1529.
Xiong N, Zhang Z, Huang J, Chen C, Jia M, Xiong J, Liu X, Wang F, Cao X, Liang Z (2011) VEGF-expressing human umbilical cord mesenchymal stem cells, an improved therapy strategy for Parkinson’s disease. Geneer 18:394-402.
Yan M, Sun M, Zhou Y, Wang W, He Z, Tang D, Lu S, Wang X, Li S, Wang W (2013) Conversion of human umbilical cord mesenchymal stem cells in Wharton’s jelly to dopamine neurons mediated by the Lmx1a and neurturin in vitro: potential therapeutic application for Parkinson’s disease in a rhesus monkey model. PLoS One 8:e64000.
Yang S, Sun HM, Yan JH, Xue H, Wu B, Dong F, Li WS, Ji FQ, Zhou DS (2013a) Conditioned medium from human amniotic epithelial cells may induce the di ff erentiation of human umbilical cord blood mesenchymal stem cells into dopaminergic neuron-like cells. J Neurosci Res 91:978-986.
Yang S, Xue DD, Wu B, Sun HM, Li XS, Dong F, Li WS, Ji FQ, Zhou DS (2013b) Pleiotrophin is involved in the amniotic epithelial cell-induced di ff erentiation of human umbilical cord blood-derived mesenchymal stem cells into dopaminergic neuron-like cells. Neurosci Lett 539:86-91.
Zhang Y, Wang J, Ren M, Li M, Chen D, Chen J, Shi F, Wang X, Dou J (2014) Gene therapy of ovarian cancer using IL-21-secreting human umbilical cord mesenchymal stem cells in nude mice. J Ovarian Res 7:8.
Zhao C, Li H, Zhao XJ, Liu ZX, Zhou P, Liu Y, Feng MJ (2016) Heat shock protein 60 a ff ects behavioral improvement in a rat model of Parkinson’s disease grafted with human umbilical cord mesenchymal stem cell-derived dopaminergic-like neurons. Neurochem Res 41:1238-1249.
Zhuang H, Zhang R, Zhang S, Shu Q, Zhang D, Xu G (2015) Altered expression of microRNAs in the neuronal di ff erentiation of human Wharton’s Jelly mesenchymal stem cells. Neurosci Lett 600:69-74.
Zuk PA, Zhu M, Mizuno H, Huang J, Futrell JW, Katz AJ, Benhaim P, Lorenz HP, Hedrick MH (2001) Multilineage cells from human adipose tissue: implications for cell-based therapies. Tissue Eng 7:211-228.
Copyedited by Li CH, Song LP, Zhao M
Mossa Gardaneh, Ph.D., mossabenis65@gmail.com.
10.4103/1673-5374.211201
*< class="emphasis_italic">Correspondence to: Mossa Gardaneh, Ph.D., mossabenis65@gmail.com.
orcid: 0000-0003-3036-2929 (Mossa Gardaneh) 0000-0002-1987-4563 (Mahdi Eskandarian Boroujeni)
杂志排行

中国神经再生研究(英文版)的其它文章
- SoxC transcription factors in retinal development and regeneration
- Recovery of sympathetic nerve function after lumbar sympathectomy is slower in the hind limbs than in the torso
- Targeting 14-3-3 adaptor protein-protein interactions to stimulate central nervous system repair
- RACK1 regulates neural development
- Schwann cell development, maturation and regeneration: a focus on classic and emerging intracellular signaling pathways
- BDNF pro-peptide: a novel synaptic modulator generated as an N-terminal fragment from the BDNF precursor by proteolytic processing