中国涡虫新纪录科微口涡虫属一新种(大口虫目,微口虫科)及其分子系统学分析
2017-07-31林逸涛庄洁怡陈龙钟林宏汪安泰
林逸涛, 庄洁怡, 陈龙, 钟林宏, 汪安泰
(深圳大学生命与海洋科学学院,深圳市海洋生物资源与生态环境重点实验室,广东深圳518060)
中国涡虫新纪录科微口涡虫属一新种(大口虫目,微口虫科)及其分子系统学分析
林逸涛, 庄洁怡, 陈龙, 钟林宏, 汪安泰*
(深圳大学生命与海洋科学学院,深圳市海洋生物资源与生态环境重点实验室,广东深圳518060)
本文报道中国涡虫新纪录科,中国微口涡虫,新种Microstomumsinensisn. sp.。标本采自中国广东省深圳市南山区蛇口码头海边。该新种具有横裂殖和有性生殖个体。所有个体均具雄性生殖器官,其交配刺骨质,整体呈钩状,全长60~75 μm,基部呈漏斗状,从基部到末端刺管直径逐渐缩小。交配刺后1/3弯曲位于基部所在平面之外,弯曲程度接近半圆,末端开口呈尖叶状。另对其16S rDNA和28S rDNA进行了分子系统学分析,确定了其分类地位。
大口虫目; 微口虫科; 微口涡虫属; 新种; 形态学; 分子系统发生
微口涡虫属Microstomum隶属于大口虫目Macrostomorpha微口虫科Microstomidae,迄今全球共记录37种(http://turbellaria.umaine.edu/),亦有称39种(http://macrostomorpha.info/)。微口涡虫属的研究始于1848年(Schmidt,1848),研究数据较为贫乏,缺少影像资料,分子系统学仅7个物种有数据(16S rDNA和28S rDNA)(Janssenetal.,2015)。中国仅见无脊椎动物教材有简要介绍(江静波,1965),尚未见微口虫科的多样性研究报道。微口涡虫属物种的特征是全身被纤毛,消化道盲端达到口前,存在有性和无性两种生殖方式,雌孔位于雄孔之前。
本研究于2017年2月在中国深圳湾海域采集到微口涡虫属1种,通过个体形态、组织学、精子形态及分子系统学的详细研究,确定其新种地位。最后,对中国涡虫的研究现状进行了讨论。
1 材料方法
1.1采集与培养
在深圳湾潮汐低位时,用扫把刷洗礁石表面,同时用200目的水生生物网于水底及沉水礁石间扫网。网内物洗入2 350 mL保鲜盒,带回实验室镜检。分离鉴定后的涡虫置于培养皿,10只涡虫加入生境水50 mL,于室温(25 ℃±1 ℃)下培养,每日定时加入适当蒸馏水以补充蒸发水分。
1.2标本制备、观察及数据处理
使用含5%乙醇的生境水麻醉标本,Bouin氏液(甲醛14 mL,冰乙酸6 mL,饱和苦味酸80 mL)固定5 h,水洗2 d,切片制作使用莱卡切片机(Leica RM2235),切片厚6 μm。切片与整装片标本使用苏木精-伊红染色,常规组织学方法制片。乳酸酚溶液(乳酸:苯酚=1∶1,v/v)用于骨质交配刺分离。
使用莱卡体视显微镜(Leica,EZ4)分离涡虫个体;骨质交配刺、精子、整装片与组织切片标本在奥林巴斯微分干涉显微镜(Olympus BX51 PA,USA)下观测与分析,专业数码相机(Olympus DP72 PA,USA)拍照;奥林巴斯专业软件(Olympus,DP 2-BSW,版本2.2)偶联用于拍摄活体、切片、精子和交配刺的照片,并用其测量分类性状;Photoshop(CS6)用于模式图的绘制;PCR使用美国应用生物系统PCR仪(Applied Biosystems 2720);BIofriend蓝光切胶仪(BD-BGC1)用于切胶回收。
1.3分子系统学分析
个体饥饿处理3 d,液氮处理15 s。使用E.Z.N.ATMMollusk DNA Isolation Kit(Omega,Norcross,GA,USA)提取涡虫DNA。16S rDNA引物为Mac-16SF和Mac-16SR(Janssenetal.,2015),28S rDNA引物为ZX-1a和1200R(Schäreretal.,2011)(表1)。以上引物由上海生工生物工程技术服务有限公司合成。PCR产物切胶回收纯化,得到的产物连接、克隆到pMDTM19-T Vector Cloning Kit (TaKaRa,中国大连),然后转化到制备成感受态的大肠杆菌EscherichiacoliTop 10中。涂板、挑取单克隆,培养后形成的菌液由北京基因组研究所进行测序(华大基因测序,中国深圳)。分子系统学分析所用序列均来源于GenBank(Schäreretal.,2011;Laumer & Giribet,2014;Janssenetal.,2015)(表2),本文所得序列均上传至GenBank。MEGA 7.0用于序列分析。
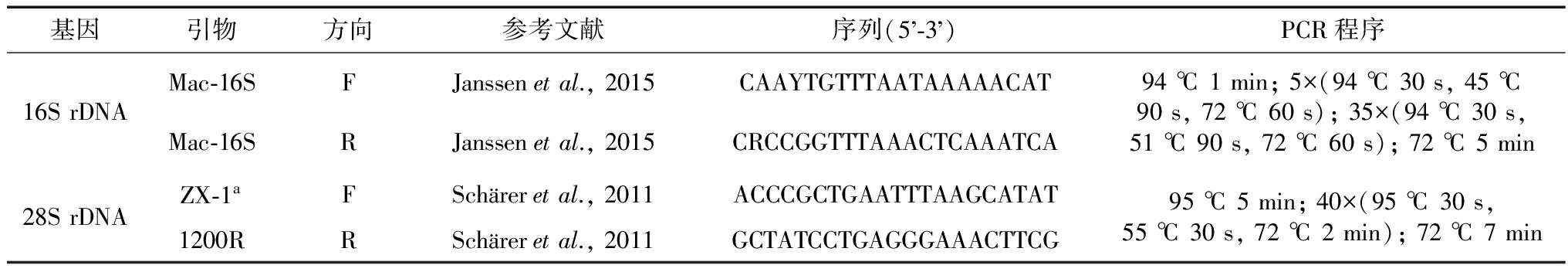
表1 本文所用引物及PCR程序Table 1 Primers and PCR protocols

表2 本文所用物种基因序列的GenBank登录号Table 2 GenBank accession numbers used in the study
1.4缩写词
ad.黏附腺adhesive glands;b.脑brain;eg.辐射状腺osinophilic glands;fg.雌性生殖孔female gonopore;fs.裂殖缝fragmentation seam;i.肠intestines;m.口mouth;mg.雄性生殖孔male gonopore;o.卵巢ovary;ps. 交配刺penis stylet;pg.咽腺pharyngeal glands;rh. 杆状体rhabdites;t.精巢testis;vg.颗粒囊vesicula granulorum。
2 结果
2.1中国微口涡虫,新种Microstomumsinensisn.sp.Wang&Lin(图1~图5)
模式标本:正模PLA-Mi001,整装片。2017年采自中国广东省深圳市南山区蛇口码头海边(113°54′48.50″E,22°28′58.30″N),水温18~20 ℃,盐度17‰~21‰,采集人林逸涛。副模PLA-Mi002~004,整装片;PLA-Mi005~007,连续组织切片;采集记录同正模。所有标本均保存于中国科学院动物研究所国家动物博物馆标本馆。
2.2形态学特征
虫体无色素,游动时呈棒状。体长800~1 200 μm(n=5),宽200~250 μm(n=5)(图1:A)。全身被密集纤毛,长2~3 μm(n=6)。背部表皮内分布有稀疏的杆状体束(长13~42 μm,1~4根/束,n=9),深入到皮下组织,头部的杆状体略多(图2:A~D)。吻端呈长圆弧形(图1:C)。脑位于虫体头部腹侧(图2:D,图3:A)。口位于脑后腹侧,咽部分布有咽腺(图1:A~C,图2:D,图3:A)。消化道与口连接,延伸至虫体前后两端,约占虫体的90%(图1:A~B,图3:A)。虫体分为裂殖体和有性生殖体,两者均具备雄性生殖系统(图1:A~B,图2:A~D,图3:A)。
裂殖体无雌性生殖系统,由前后2个裂殖段组成,由裂殖缝连接(图1:A、D,图2:A,图3:A-Ⅰ)。个体通过裂殖缝进行横断裂无性生殖。
有性生殖体具雌性生殖系统,无裂殖缝(图1:B,图2:C~D,图3:A-Ⅱ)。雌性生殖系统由卵巢和雌孔构成。卵巢位于虫体中部,长180~200 μm,宽150~180 μm(n=3)。雌孔位于卵巢后方,其腹侧环绕着大量放射状腺体,受精卵通过雌孔产出体外(图2:C~D,图3:A~B)。

图1 中国微口涡虫Microstomum sinensis n. sp.活体图、骨质交配刺和成熟精子Fig. 1 Microstomum sinensis n. sp. living specimen, penis stylet, and mature sperm
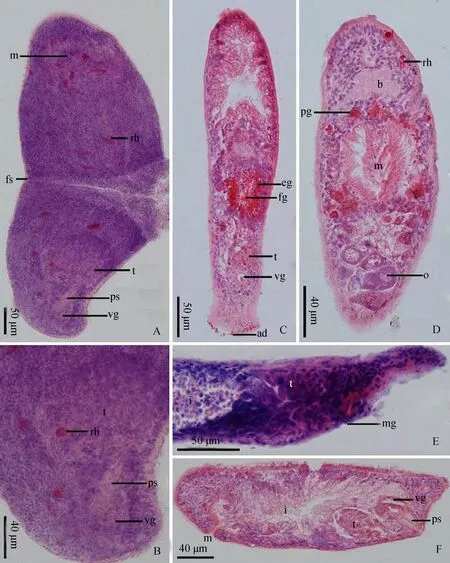
图2 中国微口涡虫Microstomum sinensis n. sp.整装片和连续组织切片Fig. 2 Microstomum sinensis n. sp. mounted specimens and serial sections
雄性生殖系统位于虫体尾部腹侧,由精巢、颗粒囊、骨质交配刺和雄孔组成(图1:E,图2:B、F,图3:A~C)。精巢呈椭球形,长120~150 μm,宽60~80 μm,其后连接颗粒囊(n=3)。颗粒囊位于精巢后方,呈球形,直径为35~45 μm(n=3),其后端连接骨质交配刺。骨质交配刺位于颗粒囊腹侧,朝向前方。交配刺整体呈钩状,全长60~75 μm(n=3),基部呈漏斗状,直径22~28 μm(n=3),从基部到末端刺管直径逐渐缩小。交配刺后1/3弯曲位于基部所在平面之外,弯曲程度接近半圆。末端开口呈尖叶状,直径2.5~3.5 μm(n=3),朝向腹前侧的雄孔(图1:F,图2:E,图3:D)。雄孔位于虫体腹侧,交配刺前方。精子呈梭形,全长30~35 μm(n=5),头部较尖,尾部有1根尾毛,长7~9 μm(n=3),有摆动功能(图1:G,图3:E)。
2.3分类学比较
中国微口涡虫,新种Microstomumsinensisn. sp.全身被纤毛,消化道盲端达到口前,雌孔位于雄孔之前,存在有性和无性2种生殖方式,故隶属于大口虫目微口虫科微口涡虫属。
微口涡虫属物种除交配刺外均为肉质,骨质交配刺是最重要的鉴定特征。
在微口涡虫属中,与本文新种在形态学上相似的物种有5种,其骨质交配刺均指向虫体前方,分别是:M.papillosum(Graff, 1882)、M.jenseni(Riedel, 1932)、M.bispiralis(Stirewalt, 1937)、M.trichotum(Marcus, 1950)和M.spiculifer(Faubel, 1974)。

图3 中国微口涡虫Microstomum sinensis n. sp.模式图Fig. 3 Microstomum sinensis n. sp. patterns
M.jenseni、M.spiculifer和M.trichotum的裂殖体形态存在多个裂殖段,前两者裂殖体无雄性生殖系统。M.jenseni交配刺整体呈波浪状,全长60 μm。M.spiculifer交配刺呈獠牙状。M.trichotum交配刺整体呈短钩状。本文新种裂殖体具雄性生殖系统,交配刺整体呈钩状,后端弯曲接近半圆,全长60~75 μm,故与上述物种差异极为显著。
M.bispiralis的裂殖体形式存在2个裂殖段,无雄性生殖系统;有性生殖体交配刺整体呈闪电状,与精巢距离较远。而本文新种裂殖体可见雄性生殖器官,交配刺整体呈钩状,精巢与交配刺位置接近,故两者差异明显。
M.papillosum的形态特征与本文新种近似,其骨质交配刺整体呈C形,全长80 μm。另外其精巢呈长条形;卵巢位于虫体中后部;雄孔位于虫体尾部腹面左侧。而中国微口涡虫,新种M.sinensisn. sp.的骨质交配刺整体呈钩状,全长60~75 μm,其精巢呈椭球形,卵巢位于虫体中部,雄孔位于交配刺前方,虫体中部,故两者差异十分明显。
综上所述,中国微口涡虫新种是成立的。
2.4分子系统学分析
16S rDNA为线粒体核糖体基因,28S rDNA为核糖体大亚基基因,这2个基因具有较高的保守性,在系统进化的研究上具有重要意义,并可以据此推断物种间的亲缘关系。利用MEGA软件将本文新种3个样本与6种微口涡虫属物种、5种大口虫目其他科属涡虫、1外群Haplopharynxrostratus的16S rDNA和28S rDNA基因序列(表2)分别匹配,以基于距离的邻接法(NJ法)和基于统计学的最大似然法(ML法)建立系统发育树(图4~图5)。
NJ法和ML法构建的发育树结果一致(图4:A~B,图5:A~B),16S rDNA与28S rDNA结果一致。由系统树可知,本文新种3个个体(图4~图5:“▲”标号)的序列集中于一支,位于微口涡虫属,并与Bradynectessterreri、Dolichomacrostomumuniporum、Karlingialutheri和Myozonariafissipara汇为一支,共同属于Dolichomicrostomida,分类地位明确。另外,M. sp. C与本文新种亲缘关系较近。
3 讨论
江静波(1965)曾在教材中介绍一种淡水微口涡虫Microstomumsp.捕食水螅Hydra,并利用水螅刺丝囊作为自身防御武器的观察记录。迄今,尚未见中国的微口涡虫多样性研究的记录。本研究首次在中国海域发现微口虫科微口涡虫属新种,即中国微口涡虫Microstomumsinensisn. sp.,这是微口虫科在中国的首次报道。世界微口涡虫属的研究较为缓慢,分子系统学方面仅有M.papillosum、M.lineare、M. sp.A~D(Janssenetal.,2015)有相关研究。本文对中国微口涡虫做了16 S rDNA和28S rDNA的分子系统发生分析,丰富了世界微口涡虫属分子系统学研究数据,为相关研究提供重要的参考数据。
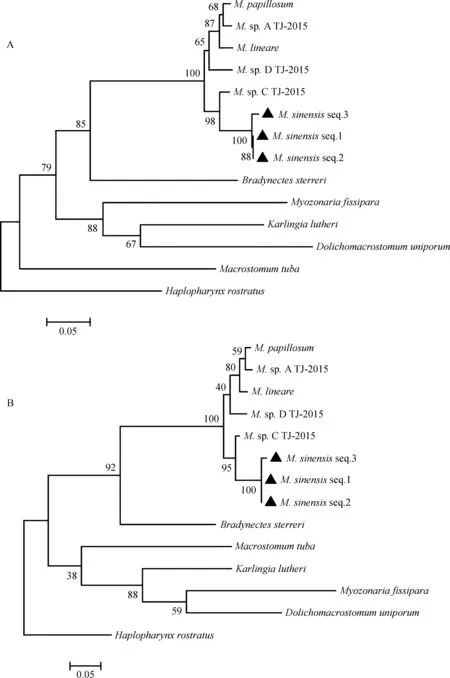
图4 16S rDNA发育树Fig. 4 Phylogenetic tree of 16S rDNA

图5 28S rDNA发育树Fig. 5 Phylogenetic tree of 28S rDNA
中国的涡虫多样性研究进展缓慢,20世纪中国学者仅记录淡水涡虫21种(孙婷等,2016),均分布于古北界。东洋界的淡水涡虫仅记录日本三角涡虫Dugesiajaponica(Ichikawa & Kawakatsut,1964)。2004年以来,在中国发现分布于东洋界的涡虫物种达39种(汪安泰等,2004;汪安泰,罗振国,2004;汪安泰,2004,2005;汪安泰,李慧,2005;汪安泰,吴海龙,2005a,2005b,2008;汪安泰,邓利,2006;彭杉等,2007;张小英等,2010;赵鹏飞等,2011;汪安泰,孙源,2011;Laietal.,2013;Luetal.,2013;Yuetal.,2013;马柳安等,2014;孙婷,汪安泰,2014;Xiaetal.,2014;Zhangetal.,2014;Chen XMetal.,2015 Chen YHetal.,2015;Sunetal.,2015;Fangetal.,2016;容粗徨等,2016;Heetal.,2016;张吕等,2017)。其中与本文新种同属于大口虫目的物种11种,分别是管大口涡虫Macrostomumtuba(汪安泰等,2004)、厦门大口涡虫Mac.xiamensis(汪安泰,罗振国,2004)、中国大口涡虫Mac.sinensis、针大口涡虫Mac.acus、钝大口涡虫Mac.obtuse(汪安泰,2005)、帆大口涡虫Mac.saifunicum(赵鹏飞等,2011)、异尾大口涡虫Mac.bicaudatum、河源大口涡虫Mac.heyuanensis、东源大口涡虫Mac.dongyuanensis(Sunetal.,2015)、珠江大口涡虫Mac.zhujiangensis、宝安大口涡虫Mac.baoanensis(Fangetal.,2016)。动物地理区划对于物种分布的了解具有重要指导意义。中国横跨古北界和东洋界,湖泊河流等水系与地貌结构多样性丰富,物种资源十分丰富(Chen,2004)。本团队探索的水域目前仅限于东洋界的部分淡水水域。2013年以后开始咸淡水环境的涡虫多样性研究,发现了深圳米罗涡虫Miroplanashenzhensis(Yuetal.,2013)、格氏肠口涡虫Enterostomulagraffi(马柳安等,2014)、澳洲异尾涡虫Heterochaerusaustralis(孙婷,汪安泰,2014)(无肠目Acoela现已移出扁形动物门Platyhelminthes,孙婷等,2016)、中国五肠涡虫Pentacoelumsinensis(Heetal.,2016)、珠江大口涡虫Mac.zhujiangensis和宝安大口涡虫Mac.baoanensis(Fangetal.,2016)、铲形杰氏涡虫Gieysztoriaknipovici(张吕等,2017),全部分布在深圳沿海。中国的海栖类涡虫多样性研究刚起步,分子系统学相关研究较为鲜见。中国的海岸线很长,跨越东洋界与古北界,提示许多未知的涡虫物种有待发现与描述。
致谢:2015级冯伟涛同学协助查阅文献,在此衷心感谢!
江静波. 1965. 无脊椎动物学[M]. 北京: 高等教育出版社: 118.
马柳安, 容粗徨, 汪安泰. 2014. 中国涡虫一新纪录科肠口涡虫属一新纪录种格氏肠口涡虫(原卵黄目, 柱口科)[J]. 动物学杂志, 49(2): 244-252.
彭杉, 刘笑媚, 汪安泰, 等. 2007. 中国涡虫一新纪录目一新纪录种(卵黄上皮目,原吻科)[J]. 动物分类学报, 32(2): 433-437.
容粗徨, 刘旭坤, 赵佳琪, 等. 2016. 达氏科(扁形动物门, 单肠目) 涡虫二新种[J]. 四川动物, 35(1): 97-103.
孙婷, 何一, 方楚玉, 等. 2016. 华南常见涡虫类群的的分布与鉴别[J]. 生物学通报, 51(3): 47-51.
孙婷, 汪安泰. 2014. 澳洲异尾涡虫(无肠目, 盘旋科)新纪录及描述[J]. 动物学杂志, 49(3): 415-422.
汪安泰, 邓利. 2006. 杰氏涡虫属一新种及中国一新纪录种(扁形动物门, 单肠目, 达氏科)[J]. 动物分类学报, 31(1): 120-124.
汪安泰, 胡好远, 罗振国. 2004. 管大口涡虫生物学特性的观察[J]. 动物学杂志, 39(4): 55-58.
汪安泰, 李慧. 2005. 旋涡虫属一新种(单肠目, 隐头吻亚目, 多囊科)[J]. 动物分类学报, 30(4): 721-724.
汪安泰, 孙源. 2011. 中国涡虫一新纪录科(扁形动物门, 单肠目, 盲扁虫科)背睾涡虫属一新种[J]. 动物分类学报, 36(1): 159-164.
汪安泰, 吴海龙. 2005a. 达氏科中国一新纪录属及三新种(扁形动物门, 单肠目, 达氏亚目)[J]. 动物分类学报, 30(2): 300-308.
汪安泰, 吴海龙. 2005b. 达氏科中国一新纪录属一新种(扁形动物门, 单肠目, 达氏亚目)[J]. 动物分类学报, 30(3): 516-519.
汪安泰, 吴海龙. 2008. 小达氏涡虫属一新种(扁形动物门, 单肠目, 达氏科)[J]. 动物分类学报, 33(1): 123-127.
汪安泰, 罗振国. 2004. 中国大口涡虫属一新种记述(大口虫目, 大口虫科)[J]. 动物分类学报, 29(4): 700-703.
汪安泰. 2004. 中国涡虫一新纪录科达氏涡虫属一新种(单肠目,达氏科)[J]. 动物分类学报, 29(4): 697-699.
汪安泰. 2005. 中国大口涡虫属三新种(扁形动物门, 大口虫目, 大口虫科)[J]. 动物分类学报, 30(4): 714-720.
张吕, 蔡慧, 鲍思全, 等. 2017. 铲形杰氏涡虫(单肠目, 达氏科)新纪录的形态学描述及其分子系统学分析[J]. 四川动物, 36(1): 65-74.
张小英, 李逸, 汪安泰. 2010. 小达氏涡虫属(扁形动物门, 单肠目, 达氏科)亚洲一新纪录种[J]. 四川动物, 29(6): 950-953.
赵鹏飞, 罗慧琳, 汪安泰. 2011. 大口涡虫属中国一新纪录(扁形动物门, 大口涡虫)及其分类性状[J]. 四川动物, 30(1): 39-44.
Chen L. 2004. The precise biogeographic division of Palaearctic and Oriental Realm in the east of China: based on data of amphibians[J]. Zoological Research, 25(5): 369-377.
Chen XM, Chen YH, Wu CC,etal. 2015. A new species of the genusGirardia(Tricladida: Dugesiidae) from China[J]. Zoological Systematics, 40(2): 166-178.
Chen YH, Chen XM, Wu CC,etal. 2015. A new species of the genusDugesia(Tricladida: Dugesiidae) from China[J]. Zoological Systematics, 40(3): 237-249.
Fang CY, Wang L, Zhang Y,etal. 2016. Two new species of brackish-waterMacrostomum(Platyhelminthes, Macrostomida) from southern China[J]. Zootaxa, 4170(2): 298-310.
Faubel A. 1974. Macrostomida (Turbellaria) von einem Sandstrand der Nordseeinsel Sylt[M]. Mikrofauna des Meeresboden, 45: 1-32 [339-370]. 349-352.
Graff L. 1882. Monographie der Turbellarien I. Rhabdocoelida [J]. Verlag Wilhelm Engelmannm, Leipzig, (2): 252.
He Y, Zhao JQ, Ning WR,etal. 2016. A new species of the genusBothrioplana(Platyhelminthes: Bothrioplanida: Bothrioplanidae) and a new species of the genusPentacoelum(Tricladida: Bdellouridae) from southern China[J]. Zootaxa, 4179(2): 209-224.
Ichikawa A, Kawakatsu M. 1964. A new freshwater planarian,Dugesiajaponica, commonly but erroneously known asDugesiagonocephala(Duges)[J]. Annotationes Zoologicae Japonenses, 37: 185-194.
Janssen T, Vizoso DB, Schulte G,etal. 2015. The first multi-gene phylogeny of the Macrostomorpha sheds light on the evolution of sexual and asexual reproduction in basal Platyhelminthes[J]. Molecular Phylogenetics & Evolution, 92: 82-107.
Lai XT, Lu YH, Wang AT. 2013. A new species of the genusGieysztoria(Platyhelminthes, Rhabdocoela, Dalyelliidae) from China[J]. Acta Zootaxonomica Sinica, 38(2): 251-256.
Laumer CE, Giribet G. 2014. Inclusive taxon sampling suggests a single, stepwise origin of ectolecithality in Platyhelminthes[J]. Biological Journal of the Linnean Society, 111(3): 570-588.
Lu YH, Wu CC, Xia XJ,etal. 2013. Two new species ofGieysztoria(Platyhelminthes, Rhabdocoela, Dalyelliidae) from a freshwater artificial lake in Shenzhen, China[J]. Zootaxa, 3745(5): 569-578.
Marcus E. 1950. Turbellaria Brasileiros (8) [J]. Boletins da Faculdade de Pilosofia, Siencias e Letras, Universidade So Paulo (Zoologia), 15: 5-192.
Riedel G. 1932. Ergebnisse einer von E. Reisinger & O. Steinböck mit Hilfe das Rask-Orsted Fonds durchgeführten zoologischen Reise in Grönland 1926. 3. Macrostomida. 4. Dalyelliidae[J]. Videnskabelige Meddelelser Fra Dansk Naturhist orisk Forening I Kjøbenhavn, 94: 33-107.
Schärer L, Littlewood DTJ, Waeschenbach A,etal. 2011. Mating behavior and the evolution of sperm design[J]. Proceedings of the National Academy of Sciences of the United States of America, 108(4): 1490-1495.
Schmidt O. 1848. Die rhabdocoelen Strudelwürmer (Turbellaria Rhabdocoela) des süssen Wassers[J]. Friedrich Mauke, Jena, 65.
Stirewalt MA. 1937.Microstomumbispiralisn. sp[J]. Zoologischer Anzeiger, 110: 314-320.
Sun T, Zhang L, Wang AT,etal. 2015. Three new species of freshwaterMacrostomum(Platyhelminthes, Macrostomida) from southern China[J]. Zootaxa, 4012(1): 120-134.
Xia XJ, Lu YH, Wang AT. 2014. A new species of Aequales, genusGieysztoria(Rhabdocoela: Dalyelliidae) from China[J]. Zoological Systematic, 39(2): 229-235.
Yu AQ, Wang AT, Lai XT. 2013. A first recorded family uteriporidae with a new species of the genusMiroplana(Platyhelminthes, Tricladida) from China[J]. Acta Zootaxonomica Sinica, 38(2): 257-266.
Zhang H, Li YK, Wu CC,etal.2014. Two new species of the genusGieysztoria(Rhabdocoela: Dalyelliidae) from China[J]. Zoological Systematics, 39(4): 485-495.
FirstRecordofMicrostomidae(Platyhelminthes:Macrostomorpha)fromChina,andthePhylogenyofOneNewSpeciesofMicrostomum
LIN Yitao, ZHUANG Jieyi, CHEN Long, ZHONG Linhong, WANG Antai*
(Shenzhen Engineering Laboratory of Marine Algal Biotechnology, College of Life Science and Oceanography,Shenzhen University, Shenzhen, Guangdong Province 518060, China)
Here we reported the first record of Microstomidae (Platyhelminthes: Macrostomorpha) from China:Microstomumsinensisn. sp., which was collected at the Shekou port, Nanshan district, Shenzhen, Guangdong province, China (113°54′48.50″E, 22°28′58.30″N).Microstomumsinensishas 2 ways of reproduction: transverse fragmentation and sexual reproduction. All specimens have male genital organs with penis stylets. The length of the sickle-like penis stylet is 60-75 μm. The base is in funnel shape. The diameter of penis stylet gradually decreases from the base to the distal end. The semi-suborbicular bend at the posterior 1/3 position of penis stylet locates beyond the dimension of the base. The distalopening is in sharp lobed shape. The phylogeny of the new species was studied based on partial 16S rDNA and 28S rDNAsequences to support our study.
Macrostomorpha; Microstomidae;Microstomum; new species; morphology; phylogeny
2017-02-25接受日期:2017-05-09
广东省大学生科技创新培育专项资金(“攀登计划”专项资金)项目(pdjh2017a0429); 广东省大学生创新创业训练计划项目(201710590042; 201710590044; 201710590045)
林逸涛, 男, 本科生, 研究方向:动物学, E-mail:linyitao0@outlook.com
*通信作者Corresponding author, E-mail:wang118@szu.edu.cn
10.11984/j.issn.1000-7083.20170059
Q959.151
: A
: 1000-7083(2017)04-0441-09
