东方粘虫 U6启动子的克隆及功能验证
2017-07-18王高振吴文君祁志军
王高振,金 朵,吴文君,祁志军
(西北农林科技大学 植物保护学院农药研究所,陕西杨凌 712100)
东方粘虫 U6启动子的克隆及功能验证
王高振,金 朵,吴文君,祁志军
(西北农林科技大学 植物保护学院农药研究所,陕西杨凌 712100)
为了持续获得大量短片段干扰RNA(short interference RNA, siRNA),通过染色体步移技术克隆得到东方粘虫 U6 snRNA 基因5′-端侧翼启动子序列1 426 bp。经生物信息学分析,该序列含SPH元件、八聚体序列(OCT)、远端序列元件(DSE)、近端序列元件(PSE)及TATA box等RNA聚合酶Ⅲ(RNA polymerase Ⅲ,RNA Pol Ⅲ) U6启动子的特征元件。通过构建RNA干扰(RNA interference, RNAi)载体的方式,对增强型绿色荧光蛋白EGFP基因进行RNAi,证明克隆得到的东方粘虫 U6启动子可成功驱动shEGFP表达,具有 U6启动子功能。为后续构建以东方粘虫V-ATP酶H亚基为靶基因的沉默载体奠定基础。
东方粘虫;RNA干扰; U6启动子;功能验证
RNA干扰(RNA interference, RNAi)是近年来快速发展起来的一种序列特异的转录后基因沉默技术[1],在功能基因组研究[2-3]、疾病防治[4-5]及新药研发[6-7]等方面具有广阔的应用前景。其基本原理是短片段干扰RNA(short interference RNA, siRNA)与细胞内同源的mRNA结合导致其降解,从而使目的基因表达沉默[8]。获得siRNA的方法主要有化学合成法、Dice酶切割法和短片段发卡RNA(short-hairpin RNA, shRNA)表达载体法[9]等。其中,构建shRNA表达载体的方法是唯一适合长期研究的方法,而且由于质粒可以复制扩增,相比起化学合成来说,能够显著降低制备siRNA的成本[10]。目前,介导siRNA表达的启动子主要是RNA聚合酶Ⅲ(RNA polymerase Ⅲ, RNA pol Ⅲ)启动子,常用人源和鼠源的 U6启动子及人的H1启动子等[11]。这类启动子可以促进快速合成大量shRNA,可以实现对细胞内基因的快速沉默[12]。RNA pol Ⅲ启动子序列上包含明确的起始和终止序列,转录出的RNA形成发夹结构会在3′-端形成2个突出的尿嘧啶,这类似于天然的siRNA,因而有利于诱发RNA沉默[13]。目前,通过构建基于 U6启动子的基因沉默载体在不同的细胞和生物体内已经取得了良好的基因沉默效果。例如,由家蚕 U6启动子驱动的shRNA表达载体转染家蚕BmN细胞后,显著降低家蚕核多角体病毒对细胞的伤害[14];河豚双 U6启动子双链RNA(double-stranded, dsRNA)表达载体转染鲑鱼胚胎CHSE-214细胞后,有效抑制败血症病毒在该细胞内的扩散[15];CMV-U6混合启动子shRNA表达载体介导斑马鱼相关基因的沉默[16]。由此可见,基于 U6启动子的RNAi技术已经成为基因沉默的主要手段。
目前,人们对 U6启动子的结构已经研究较为清楚,发现在转录起始位点上游包含3个启动子元件:DSE、PSE和TATA box[17]。Lambeth等[18]研究发现 U6启动子各个元件的间距、序列和结构之间存在种属差异;Wise等[19]通过比较同一载体上的鼠源和鸡 U6启动子,发现不同启动子的序列和结构对启动效率有很大影响。也就是说某一物种的 U6启动子对其他物种体内RNA pol Ⅲ的驱动能力可能降低甚至丧失。因此,要获得理想的启动效果,利用物种本源 U6启动子是最佳选择。
东方粘虫 (Mythimnaseparata(Walker)) 是一种鳞翅目害虫,其幼虫主要危害玉米和小麦等禾本科作物,具有远距离迁飞、暴食和集中危害等特点。严重发生时,短时间内就可将成片的玉米、谷子、水稻吃成根茬,防治不及时会造成严重损失,甚至绝产[20]。目前,防治东方粘虫以喷洒杀虫剂为主,但大面积使用化学农药,不但造成农药残留、杀伤害虫天敌,而且会导致害虫产生抗药性。近年来,随着基因沉默技术的发展,基于RNAi的害虫防治策略逐渐受到人们的关注[21],已经在玉米根叶甲(DiabroticavirgiferavirgiferaLeConte)[22]和烟粉虱(Bemisiatabaci)[23]等害虫防治上获得成功。在前期研究中,本实验室克隆出东方粘虫V-ATP酶H亚基,设计合成dsRNA并微量注射3龄幼虫,试虫出现生长发育受阻和死亡等典型症状[24]。为了持续获得大量的siRNA,本研究克隆出东方粘虫的 U6启动子并对其进行功能验证,为后续构建以东方粘虫V-ATP酶H亚基为靶基因的沉默载体奠定基础。
1 材料与方法
1.1 供试材料与试剂
东方粘虫,在温度为(22±1) ℃,相对湿度为60%~70%,光周期为L∶D=16 h∶8 h,光强为30 000 lx的条件下室内人工饲养,试验中选取5龄初蜕幼虫。293 T细胞系由西北农林科技大学农药研究所提供,并在含胎牛血清(φ=10%)、0.1 mg/mL硫酸链霉素和100 U/mL青霉素的DMEM高糖培养基中置于37 ℃、φ=5% CO2生化培养箱中培养,取对数生长期的细胞进行试验。
Genome walker universal试剂盒及pEGFP-N1载体购自Clontech公司,T4DNA连接酶、pGEM-T载体等购自Promega公司,ExTaqDNA聚合酶(5 U/μL)、10×ExTaqbuffer (20 mmol/L, Mg2+plus)、dNTP Mixture(2.5 mmol/L)、DNA marker、pMD18-T载体、DH5α感受态细胞等购自TaKaRa公司;限制性内切酶EcoRⅠ、BamHⅠ、HindⅢ购自NEB公司;脂质体Lipofectamine 2000、DMEM高糖培养基、胎牛血清等购自Invitrogen公司;凝胶回收纯化试剂盒、质粒提取试剂盒、昆虫基因组DNA提取试剂盒、无内毒素质粒提取试剂盒等购自Omega公司;24孔细胞培养板购自ThermoFisher scientific公司;引物及测序均由上海英骏生物技术有限公司完成。
1.2 东方粘虫基因组DNA的提取
基因组DNA的提取按照昆虫基因组DNA提取试剂盒说明书进行。
1.3 染色体步移引物设计与合成
将家蚕 U6 snRNA基因全序列(AY649381.1)与果蝇的 U6 snRNA基因全序列(AH004871.1)经在线软件Clustal Omega(http://www.ebi.ac.uk/tools/msa/clustalo/)比对,分析它们的同源区域并设计两条下游引物U6R1 和 U6R2。上游两条引物使用Genome walker universal试剂盒中的AP1和AP2。
引物序列如下:
AP1:5′-GTAATACGACTCACTATAGG-GC-3′;AP2:5′-ACTATAGGGCACGCGTGG- T-3′;U6R1:5′-CGATTTTGCGTGTCATCCTTGCGCAG-3′;U6R2:5′-TGCCTGGCTGTAAGTGCTCAGATTCC-3′。
在用户担心余额宝的自身潜在风险的时,阿里巴巴正在努力加强用户的资金提供安全和保障。例如发表承诺,如果客户账户资金被盗,支付宝方面将承诺全额赔偿。
1.4 染色体步移文库的建立
将已测定质量浓度的东方粘虫基因组DNA经4种限制性内切酶DraⅠ、EcoR Ⅴ、StuⅠ和PvuⅡ过夜酶切,消化基因组DNA,回收纯化消化产物。将纯化后的DNA与Genome walker 接头于16 ℃过夜连接,70 ℃下5 min终止反应。用TE缓冲液稀释至一定质量浓度作为PCR扩增的模板。
1.5 U6 snRNA基因上游5′-端侧翼序列的克隆
以构建好的染色体步移文库为模板,分别以AP1/U6R1和AP2/U6R2为引物,进行两轮巢式PCR。PCR程序按照染色体步移试剂盒说明书进行。将4个不同文库对应的两轮PCR产物经10 g/L琼脂糖凝胶电泳分析,并将目的条带用凝胶回收试剂盒回收纯化,再将其连接至pGEM-T载体上,转化至大肠杆菌DH5α感受态细胞中,对阳性克隆测序分析。
1.6 U6启动子的shEGFP重组载体的构建
1.6.1 pMD18-N载体的构建 根据pMD18-T载体图谱,通过T-A克隆导入一段不包含EcoRⅠ、BamHⅠ及HindⅢ酶切位点的片段N(515 bp),经双酶切检测并测序。
1.6.3 pMD18-U6与pMD18-shEGFP载体的构建 根据染色体步移法克隆得到的 U6 snRNA上游5′-端侧翼序列,设计上游引物F1:GGATCCCTAGTGCACGGTTCCAAATTGGT(斜体为BamHⅠ酶切位点),下游引物R1 :AAGCTTCATGAAAACTTTACTTGCGCGA(斜体为Hind Ⅲ酶切位点),以东方粘虫基因组DNA为模板,克隆得到上游序列1 360 bp,即为包含BamHⅠ、Hind Ⅲ酶切位点的 U6启动子。将得到的pMD18-N载体经BamHⅠ、Hind Ⅲ酶切,并用凝胶回收试剂盒回收,回收产物分别与 U6启动子及获得的双链shRNA连接,得到pMD18-U6与pMD18-shEGFP载体,并送上海英骏生物技术有限公司测序。
1.6.4 pMD18-U6-shEGFP载体的构建 按照“1.6.3”方法,设计上游引物F2:5′-GAATTCCTAGTGCACGGTTCCAAATTGGT-3′(斜体为EcoRⅠ酶切位点),下游引物R2 :5′-GGATCCCATGAAAACTTTACTTGCGCGA-3′(斜体为BamHⅠ酶切位点),克隆得到包含EcoRⅠ、BamHⅠ酶切位点的 U6启动子。将pMD18-N载体经EcoRⅠ、BamHⅠ酶切,并用凝胶回收试剂盒回收,回收产物与 U6启动子连接,得到pMD18-U6-N质粒;将pMD18-U6-N经BamHⅠ、HindⅢ酶切,凝胶回收后与shRNA连接,得到 pMD18-U6-shEGFP载体,并送上海英骏生物技术有限公司测序。
1.7 细胞转染
将处于对数期的293 T细胞铺至24孔板内培养,在倒置显微镜下观察,当细胞融合度达70%左右时进行下一步试验。将构建好的pMD18-U6-shEGFP、pMD18-U6、pMD18-shEGFP载体分别与pEGFP-N1载体共转染293 T细胞。具体方法见表1。
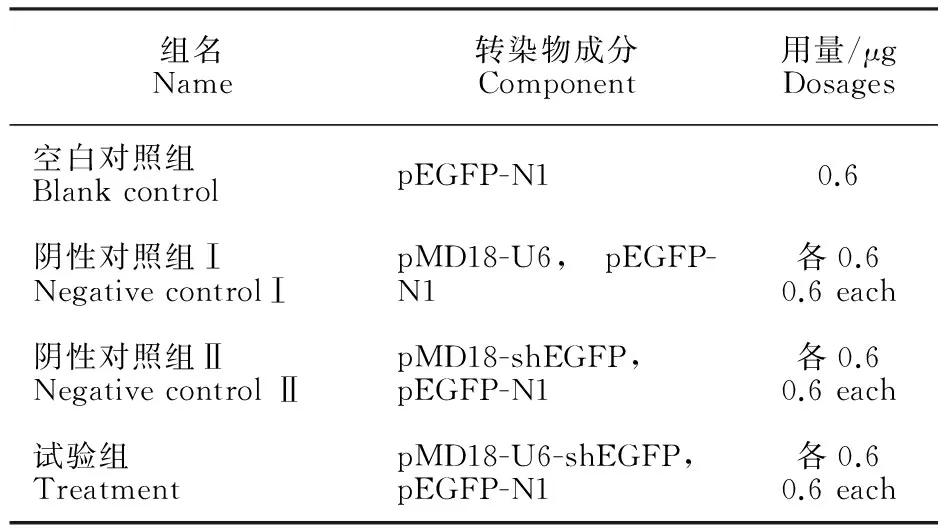
表1 转染组及转染成分 Table 1 Transfected group and compositions
2 结果与分析
2.1 U6 snRNA基因上游5′-端侧翼基因克隆与序列分析
采用染色体步移法获得东方粘虫 U6 snRNA基因上游5′-端侧翼序列并测序,该序列长1 426 bp。经NCBI数据库序列比对发现,该序列下游3′-端与家蚕(Bombyxmori) U6 snRNA基因序列(AY649381)相似度为100%,与黑腹果蝇(Drosophilamelanogaster) U6 snRNA基因序列(M24605)相似度为96%,与豹纹蝶(Melitaeacinxia) U6 snRNA基因序列(JX878560.1)相似度为96%。由此推断这些相似序列为东方粘虫 U6 snRNA基因序列。经启动子在线分析软件NNPP v. 2. 2 (http://www.fruitfly.org/ seq_tools/ promoter.html) 预测,东方粘虫 U6 snRNA 基因上游5′-端侧翼序列有4个可能的转录起始位点(图1),其转录起始位点上游存在着 TATA box,符合真核生物基因转录起始位点所具有的特征。
参照Domitrovich等[25]的方法寻找东方粘虫 U6启动子元件,结果发现,该序列中包含RNA聚合酶Ⅲ启动子的典型基序,包括远端序列元件DSE,近端序列元件PSE及TATA box,DSE由1个能够与转录因子Oct-1相结合的八聚体序列(OCT),和1个与刺激反式作用因子(stimulated trans-acting factor, STAF)相结合的SPH元件组成(图2)。
2.2 东方粘虫 U6启动子的克隆
根据东方粘虫 U6 snRNA基因上游5′-端侧翼序列,设计两对引物F1/R1、F2/R2,以基因组为模板,得到分别含BamHⅠ、Hind Ⅲ与含EcoRⅠ、BamHⅠ的 U6启动子(图3)。
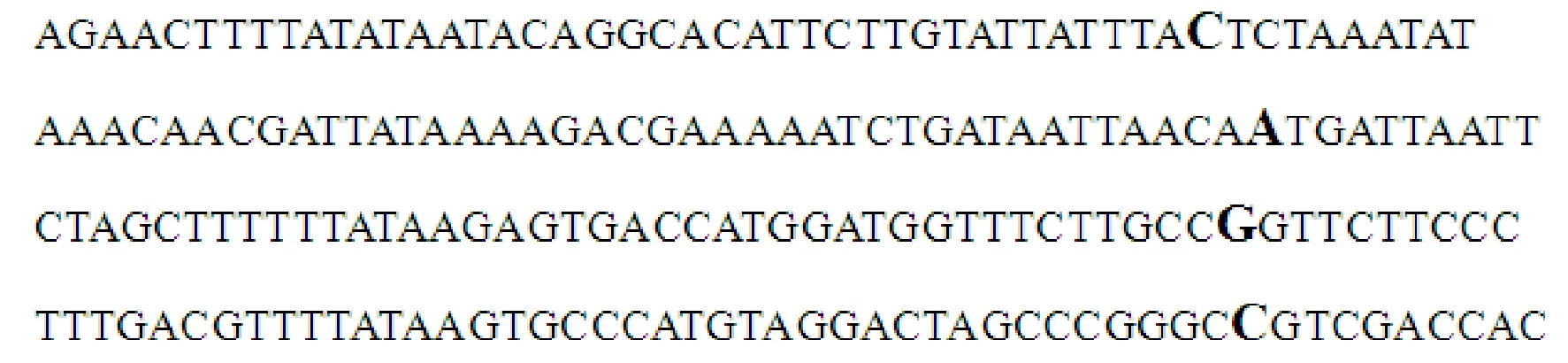
加粗字母代表可能的转录起始位点 The bold letters mean prediction of transcription start site

OCT, SPH, PSE, TATA-box都用下划线标出,预测的转录起始位点作为+1,并被框出 OCT, SPH, PSE, TATA box were underlined, and start site of the predicted transcription were recorded as +1, and was out of the box.
图2 东方粘虫 U6 snRNA基因5′-端侧翼序列分析
Fig.2 Analysis of 5′-flanking region of U6 snRNA gene inM.separata

M. Marker DL 5000; 1. 含BamHⅠ、Hind Ⅲ的 U6启动子 U6 promoter withBamHⅠ andHind Ⅲ; 2. 含EcoRⅠ、BamHⅠ的 U6启动子 U6 promoter withEcoRⅠ andBamHⅠ
图3 具有酶切位点的东方粘虫 U6启动子
Fig.3 U6 promoters ofM.separatawith restriction sites
2.3 基因沉默载体的构建与双酶切验证
为了构建基因沉默载体,首先需要对pMD18-T载体进行环化,为此设计并克隆一段515 bp的片段N,该片段不包含构建载体所需要的3个酶切位点:EcoRⅠ、BamHⅠ及Hind Ⅲ。经T-A克隆后,双酶切验证N片段成功导入pMD18-T载体中(图4)。随后分别将 U6、shEGFP及U6-shEGFP片段重组到pMD18-N,构建载体pMD18-U6、pMD18-shEGFP及pMD18-U6-shEGFP,双酶切结果表明目的片段成功转入载体(图5)。2.4 RNAi载体转染293 T细胞的荧光显微镜检测
将载体pMD18-U6、pMD18-shEGFP与pMD18-U6-shEGFP分别与pEGFP-N1共转染293 T细胞。将各孔细胞置于倒置荧光显微镜下,观察同一视野下细胞生长情况及各组荧光强度。结果如图6所示:在转染24 h、48 h与72 h后,与pEGFP-N1空白对照、pMD18-U6阴性对照和pMD18-shEGFP阴性对照相比,pMD18-U6-shEGFP的荧光强度均显著降低,表明shEGFP可以在293 T细胞中成功表达并引起荧光蛋白的表达量降低。
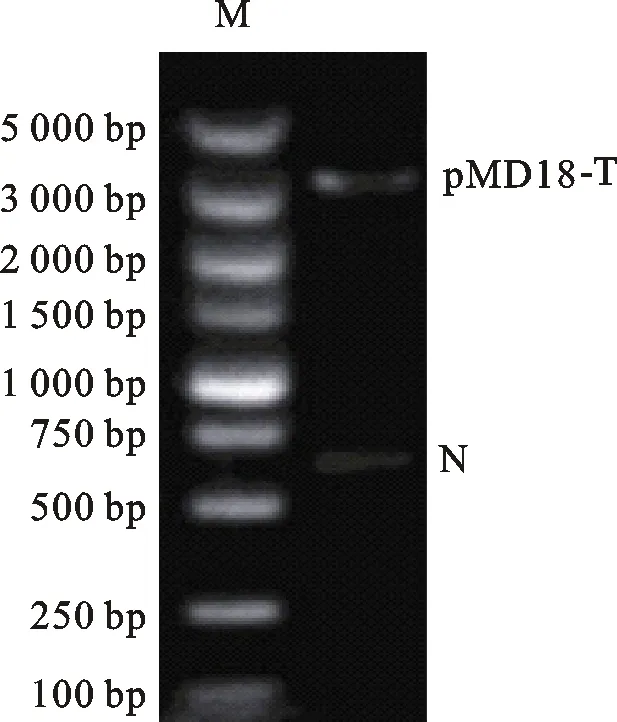
M. Marker DL 5000
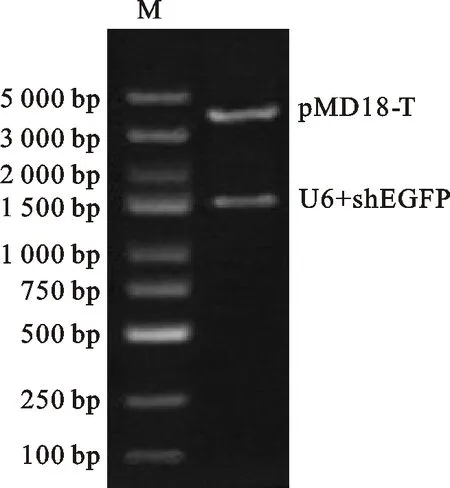
M. Marker DL 5000

图6 转染后不同时间293 T细胞荧光Fig.6 293 T cell fluorescence after transfected by different vectors with pEGFP-N1
3 讨 论
U6启动子的3′-端序列一般较为保守,但5′-端侧翼序列及其长度往往差异较大,用常规的RACE等技术很难克隆到启动子的全长序列。本研究采用染色体步移技术,首先分别用4个限制性内切酶(DraⅠ、EcoRⅤ、PvuⅡ和StuⅠ)对东方粘虫基因组DNA进行酶切,将其分成4个带相应酶切位点的DNA文库,添加特异性接头后设计上游引物,再参照近源物种家蚕和果蝇 U6 snRNA的3′-端共有序列设计下游特异性引物,用染色体步移试剂盒获得5′-端侧翼片段并逐级向前推动,最终克隆出东方粘虫 U6启动子序列。
对克隆的 U6启动子序列分析表明,该序列包含OCT,SPH, DSE, PSE及TATA-box等RNA pol Ⅲ启动子特有的序列元件,与有关文献[25-27]报道的人类、鸡及水牛等动物的 U6启动子特征元件一致。经与NCBI数据库比对发现,该 U6启动子3′-端序列与家蚕及果蝇 U6 snRNA基因部分序列均有很高的同源性(分别为100%和96%)。因此,确定此序列为东方粘虫 U6 snRNA基因,其上游5′-端侧翼序列包含东方粘虫的 U6启动子。
U6启动子能否有效驱动RNA的表达是检验 U6 snRNA是否包含整个启动子序列的关键。为了验证东方粘虫 U6启动子序列是否完整,本研究用siDicer软件对pEGFP-N1基因序列进行siRNA预测分析,随机挑选1个siRNA进行shRNA合成,并将其构建到由东方粘虫 U6启动子驱动的shRNA载体中。细胞转染证实,在24、48、72 h时,试验组荧光与空白对照组和阴性对照组均有显著差异,而空白对照组和各阴性对照组之间荧光则无明显差异。由此表明,本研究克隆获得的东方粘虫 U6 snRNA包含完整的 U6启动子序列,能成功驱动shRNA的表达并可用于后续基因沉默载体的构建。
Reference:
[1] FIRE A,XU S Q,MONTGOMERY M K,etal.Potent and specific genetic interference by double-stranded RNA inCaenorhabditiselegans[J].Nature,1998,391(6669):806-811.
[2] HARBORTH J,ELBASHIR S M,BECHERT K,etal.Identification of essential gene in cultured mammalian cells using small interfering RNAs [J].JournalofCellScience,2001,114(24):4557-4565.
[3] AZA-BLANC P,COOPER C L,WAGNER K,etal.Identification of modulator of TRAIL-induced apoptosis via RNAi-based phenotypic screening [J].MolecularCell,2003,12(3):797-801.
[4] GE Q,MCMANUS M T,NGUYEN T,etal.RNA interference of influenza virus production by directly targeting mRNA for degradation and indirectly inhibiting all viral RNA transcription [J].ProceedingsoftheNationalAcademyofSciencesoftheUSA,2003,100(5):2718-2723.
[5] LE H,KUMAR V.Specific inhibition of gene expression aid trans-activation functions of hepatitis B virus X protein and c-myc by small interfering RNAs [J].FEBSLetters,2004,560(1-3):210-214.
[6] SHIRANE D,SUGAO K,NAMIKI S,etal.Enzymatic production of RNAi libraries from cDNAs [J].NatureGenetics,2004,36(2):190-196.
[7] DYKXHOORN D M,LIEBERMAN J.Running interference:prospects and obstacles to using small interfering RNAs as small molecule drugs [J].AnnualReviewofBiomedicalEngineering,2006,8:377-402.
[8] TIJSTERMAN M,PLASTERK R H A.Dicers at RISC; the mechanism of RNAi [J].Cell,2004,117(1):1-3.
[9] HU S W,NI W,HAZI W,etal.Cloning and functional analysis of sheep U6 promoters[J].AnimalBiotechnology,2011,22(3):170-174.
[10] 楚莉辉,刘龙丁,马绍辉,等.利用人 U6 snRNA启动子构建RNA干扰质粒载体[J].中国生物化学与分子生物学报,2006,22(3):228-233.
CHU L H,LIU L D,MA SH H,etal.Constructing of an RNAi plasmid based on human U6 snRNA[J].ChineseJournalofBiochemistryMolecularBiology,2006,22(3):228-233(in Chinese with English abstract).
[11] MIYAGISHI M,TAIRA K. U6 promoter-driven siRNAs with four uridine 3'overhangs efficiently suppress targeted gene expression in mammalian cells [J].NatureBiotechnology,2002,20(5):497-500.
[12] ZHOU H,HUANG C,XIA X G.A tightly regulated Pol Ⅲ promoter for synthesis of miRNA genes in tandem [J].BiochimicaetBiophysicaActa,2008,1779(11):773-779.
[13] ELBASHIR S M,HARBORTH J,WEBER K,etal.Analysis of gene function in somatic mammalian cells using small interfering RNAs [J].Methods,2002,26(2):199-213.
[14] OHTSUKA D,NAKATSUKASA T,FUJITA R,etal.Use of bombyx mori U6 promoter for inducing gene-silencing in silkworm cells [J].JournalofInsectBiotechnologyandSericology,2008,77(3):125-131.
[15] KIM M S,JEE B Y,CHO M Y,etal.Fugu double U6 promoter-driven long double-stranded RNA inhibits proliferation of viral hemorrhagic septicemia virus (VHSV) in fish cell lines [J].ArchivesofVirology,2012,157(6):1029-1038.
[16] SU J G,ZHU Z Y,XIONG F,etal.Hybrid cytomegalovirus- U6 promoter-based plasmid vectors improve efficiency of RNA interference in zebrafish [J].MarineBiotechnology,2008,10(5):511-517.
[17] CARBON P,MURGO S,EBEL J P,etal.A common octamer motif binding protein is involved in the transcription of U6 snRNA by RNA polymerase Ⅲ and U2 snRNA by RNA polymeraseⅡ[J].Cell,1987,51(1):71-79.
[18] LAMBETH L S,WISE T G,MOORE R J,etal.Comparison of bovine RNA polymerase Ⅲ promoters for short hairpin RNA expression [J].AnimalGenetics,2006,37(4):369-372.
[19] WISE T G,SCHAFER D J,LAMBETH L S,etal.Characterization and comparison of chicken U6 promoters for the expression of short hairpin RNAs [J].AnimalBiotechnology,2007,18(3):153-162.
[20] RENJU T,GIRADDI R,HUNJE R,etal.Evaluation of new insecticidal poison baits againstMythimnaseparata(Walker) in sorghum[J].KarnatakaJournalofAgriculturalSciences,2009,22(4):7773-776.
[21] GU L Q,KNIPPLE D C.Recent advance in RNA interference research in insects:Implications for future insect pest management strategies [J].CropPrtection,2013,45:36-40.
[22] BAUM J A,BOGAERT T,CLINTON W,etal.Control of coleopteran insect pests through RNA interference [J].NatureBiotechnology,2007,25(11):1322-1326.
[23] THAKUR N,UPADHYAY S K,VERMA P C,etal.Enhanced whitefly resistance in transgenic tobacco plants expressing double stranded RNA of V-ATPaseAgene [J].PloSone,2014,9(3):e87235.
[24] 张淑静,王高振,刘 爽,等.利用RNAi技术沉默东方粘虫V-ATP酶H亚基研究[J].西北农业学报,2015,24(1):170-174.
ZHANG SH J,WANG G ZH,LIU SH,etal.Silencing of V-ATPase subunit H ofMythinmaseparatathrough RNA interference [J].ActaAgriculturalBoreali-occidentalisSinica,2015,24(1):170-174(in Chinese with English abstract).
[25] DOMITROVICH A M,KUNKEL G R.Multiple,dispersed human U6 small nuclear RNA genes with varied transcriptional efficiencies [J].NucleicAcidsResearch,2003,31(9):2344-2352.
[26] 陈新雨.鸡RNA聚合酶Ⅲ型启动子7SK克隆和转录shRNA活性研究[D].陕西杨陵:西北农林科技大学,2012:21-23.
CHEN X Y.Cloning of chicken RNA polymerase Ⅲ type Ⅲ promoter 7SK and characterization of its shRNA transcription activity in vitro[D].Yangling Shaanxi:Northwest A&F University,2012,21-23(in Chinese with English abstract).
[27] 张晓溪,刘庆友,邓彦飞,等.水牛RNA聚合酶Ⅲ启动子的克隆与鉴定[J].南方农业学报,2014,45(5):858-863.
ZHANG X X,LIU Q Y,DENG Y F,etal.Cloning and identification of bufalo RNA polymerase Ⅲ promoters [J].JournalofSouthernAgriculture,2014,45(5):858-863(in Chinese with English abstract).
(责任编辑:郭柏寿 Responsible editor:GUO Baishou)
Cloning and Functional Verification of U6 Promoter fromMythimnaseparata(Walker)
WANG Gaozhen, JIN Duo, WU Wenjun and QI Zhijun
(Institute of Pesticide Science, College of Plant Protection, Northwest A&F University, Yangling Shaanxi 712100,China)
In order to obtain constantly short interference RNA(siRNA) , the 1 426 bp 5′- flanking promoter sequence of U6 snRNA gene ofMythimnaseparata(Walker) was cloned by the method of Genome Walking, as well as its function was verified in this study. Based on bioinformatics analysis, the results showed that the promoter fragment contained polymerase Ⅲ core promoter elements SPH, OCT, DSE, PSE and TATA box. The observation of expression level enhanced green fluorescent protein EGFP after being transfected with RNA interference (RNA interference, RNAi) , the vectors by fluorescence microscope showed that the U6 promoter fragment drived the expression of shEGFP successfully. This study had established a foundation for further construction of RNAi vectors which can silence V-ATPase subunit H.
Mythimnaseparate(Walker); RNAi; U6 promoter; Functional verification
2016-05-19 Returned 2016-06-28
The National Natural Science Foundation of China (No. 31371958).
WANG Gaozhen, male, master student. Research area:pesticide toxicology. E-mail:wanggaozhen@nwsuaf.edu.cn
QI Zhijun, male, associate professor. Research area:pesticide molecular toxicology. E-mail:qzhij@nwsuaf.edu.cn
日期:2017-06-05
2016-05-19
2016-06-28
国家自然科学基金(31371958)。 第一作者:王高振,男,硕士研究生,从事农药毒理学研究。E-mail:wanggaozhen@nwsuaf.edu.cn 通信作者:祁志军,男,副教授,主要从事农药分子毒理学研究。E-mail:qzhij@nwsuaf.edu.cn
Q789
A
1004-1389(2017)06-0939-07
网络出版地址:http://kns.cnki.net/kcms/detail/61.1220.S.20170605.1728.036.html
