微生物甾醇酯酶的研究进展
2017-07-18任楠楠王晓辉迟乃玉乔慧张庆芳
任楠楠,王晓辉,迟乃玉,乔慧,张庆芳*
(1.大连大学生命科学与技术学院,辽宁大连116622;2.辽宁省海洋微生物工程技术研究中心,辽宁大连116622)
微生物甾醇酯酶的研究进展
任楠楠1,2,王晓辉1,2,迟乃玉1,2,乔慧1,2,张庆芳1,2*
(1.大连大学生命科学与技术学院,辽宁大连116622;2.辽宁省海洋微生物工程技术研究中心,辽宁大连116622)
甾醇酯酶(EC 3.1.1.13)作为一种高活性的生物催化剂,在医疗检测、食品、造纸工业及环保等领域具有极其广泛的应用价值。该文综述了甾醇酯酶的微生物来源、分离纯化、克隆表达、三维结构与应用方面的研究进展,以期为甾醇酯酶的研究开发及产业化应用奠定基础。
甾醇酯酶;微生物;克隆表达;分离纯化;应用
甾醇酯酶(EC 3.1.1.13)又称为胆固醇酯酶,广泛存在于生物体内(包括人在内的高等动物与微生物)[1]。甾醇酯酶可在水相条件下催化甾醇酯水解生成甾醇和脂肪酸,在有机溶剂中通过酯化或酯交换进行催化酯合成反应[2]。甾醇酯酶作为一种重要的酶制剂,可帮助机体消化吸收油脂类营养物质,因其与人体脂质代谢及胆固醇吸收有关,甾醇酯酶是检测血液中总胆固醇含量的重要酶制剂之一[3]。还有解决机械制浆造纸工艺中树脂障碍问题的潜力[4],由于其广泛的底物特异性和稳定性,在食品、医药、纸浆工业及化工行业等领域发挥重要作用[5]。自然界中,微生物生产的甾醇酯酶具有生产成本低、工艺简单、不易污染、不受季节影响及易形成规模化生产等优点,已成为目前研究热点[6]。本文概述了甾醇酯酶的微生物来源,介绍了酶制剂的分离纯化、基因的克隆表达和蛋白的三维结构方面的研究进展,并对其应用前景进行了展望,旨在全面深入地了解甾醇酯酶,为其理论研究与产业化应用提供参考。
1 甾醇酯酶的微生物来源
对微生物甾醇酯酶的首次报道来自UWAJIMA T等[7]的研究,之后陆续有科研人员在微生物中检测到甾醇酯酶活性。据报道[6],多种微生物都能分泌甾醇酯酶:在细菌中约几十种菌能产生甾醇酯酶,有假单胞菌属(Pseudomonas sp.)、不动杆菌属(Acinetobactersp.)、金黄杆菌属(Chryseobacteriumsp.)、蜡样芽孢杆菌属(Bacillussp.)、红球菌属(Rhodococcussp.)和伯克霍尔德氏菌属(Burkholderia sp.)等;放线菌的链霉菌属(Streptomycessp.)的菌株也能分泌甾醇酯酶;沙眼衣原体(Chlamydia trachomatis)CT149分泌的甾醇酯酶专一水解胆固醇亚油酸酯并在Hela细胞中成功表达[8];真菌中的镰刀菌属(Fusariumsp.)、木霉属(Trichodermasp.)和线嘴壳菌属(Ophiostomasp.)等的菌株均可产甾醇酯酶。不同的微生物甾醇酯酶都易形成多聚体,但其结构特点及底物特异性等差异较大。目前,对产甾醇酯酶菌株的筛选主要采用以甾醇酯为唯一碳源的方法,同时以产物中的酸为依据添加指示剂进行筛选。研究表明,微生物源的甾醇酯酶底物范围广,稳定性较好,且能通过诱导提高酶产量,纯化工艺也相对简单[9],所以目前主要针对各种微生物来源的甾醇酯酶进行研究和利用[10]。
1.1 细菌来源的甾醇酯酶
UWAJIMA T等[7]于1975年首次报道的具有水解甾醇酯活性的酶,由荧光假单胞菌(P.fluorescens)KY395分泌,分子质量为129 ku,该酶也具有脂肪酶活性,但对长链甾醇酯有显著的偏好性;最适温度55℃,在pH 5~12范围内均有活性。1976年,在相同菌种的菌株ATCC 21156中分离纯化出两种甾醇酯酶,这两种酶不具有脂肪酶活性,对甾醇酯具有绝对特异性,最适底物为胆固醇亚油酸酯。来自该菌株的酶产品由于其高产量,已获得专利保护并实现商品化。1994年,NISHIMURA M等[11]在门多萨假单胞菌(P.mendocina)3121分泌的脂肪酶中检测到了甾醇酯酶活性,酶分子质量为30 ku;2002年,SUGIHARA A等[12]描述了来自铜绿假单胞菌(P.aeruginosa)菌株的甾醇酯酶,等电点(isoelectric point,pI)为3.2,分子质量约为53 ku,其热稳定性和pH稳定性范围与来自荧光假单胞菌(P.fluorescens)的甾醇酯酶相同。
从系统发育上看,伯克霍尔德氏菌(Burkholderiasp.)与假单胞菌(Pseudomonassp.)进化关系最近,但是直到2006年,TAKEDA Y等[13]才首次从洋葱伯克霍尔德氏菌(B.cepacia)ST200中筛选出真正意义上的甾醇酯酶,该酶不具有脂肪酶活性,仅具有芳香酯酶活性。其分子质量约为37 ku,有较好的温度耐受性(4~65℃)和pH稳定性(5.5~12.0)。
国内对产甾醇酯酶微生物的研究始于20世纪80年代。杨光礼等[14]从实验室保藏的菌株中筛选出一株产甾醇酯酶能力较强的菌株,经鉴定为假单胞菌属(Pseudomonassp.),命名为SIPI-215,并对菌株发酵条件和酶学性质进行了研究;黄武华等[15]从土壤中分离出一株产甾醇酯酶能力较强的假单胞菌(Pseudomonassp.),得到菌株最适产酶条件;谭晓晶[16]首次从马红球菌(Rhodococcussp.)中分离得到甾醇酯酶,对粗酶液进行了一系列纯化,纯化后的酶分子质量58.0 ku,样品纯度达到94%,酶活10.56 U/mg,分别以胆固醇亚油酸酯和胆固醇油酸酯为底物时,酶的米氏常数(Km)值分别为2.2×105mol/L和3.1×105mol/L。曾诚等[17]以植物甾醇酯为底物,首次采用荧光透明圈法从金黄杆菌属(Chryseobacteriumsp.)中筛选出产甾醇酯酶菌株,通过分离纯化最终得到分子质量大小为39.0 ku的蛋白。该酶比酶活为2.443IU/mg,最适pH值为7.5,最适作用温度40℃,在45℃以下,pH 5.0~9.0的范围内稳定性较好。
1.2 放线菌来源的甾醇酯酶
KAMEIT等[18]报道了来自淡紫灰链霉菌(Streptomyces lavendulae)H-646-SY的甾醇酯酶,首次设计纤维素亲和层析的纯化方法,分离出具有甾醇酯酶活性的蛋白,分子质量为12.0 ku,这与NISHIMURA M等[11]由其核苷酸序列推导的结果类似。之后又从链霉菌属的不同菌株中分离出三种新的甾醇酯酶,这三种酶的底物特异性范围广,水解不同长度的甾醇酯,优选胆固醇亚油酸酯、对硝基苯基酯和甘油三酯。
1.3 真菌来源的甾醇酯酶
真菌来源的甾醇酯酶研究起步较晚。目前,研究较热门的是从线嘴壳菌(O.piceae)的甾醇酯酶。CALERO-RUEDAO等[19]首次在从线嘴壳菌(O.piceae)CECT 20416菌株中通过疏水层析的单一步骤纯化得到分子质量为56.5ku的甾醇酯酶,该酶底物特异性范围广,除了水解甾醇酯外,还能水解不同长度的脂肪酸酯,在酸性条件下24 h仍能保持50%的活性,但当pH>8或温度>60℃时稳定性较差。该酶催化效率远高于目前商业用的真菌甾醇酯酶(来自皱褶假丝酵母(C.rugosa)家族),其商业化应用目前正在开发中。CEDILLO V B等[20]将其在毕赤酵母(P.pastoris)中重组表达,因在重组蛋白的N-末端掺入了额外的6~8个氨基酸,蛋白的聚集性降低,酶活显著提高。VAQUERO M E等[21]将从线嘴壳菌(O.piceae)甾醇酯酶在酿酒酵母(S.cerevisiae)中重组表达,其催化效率介于毕赤酵母(P.pastoris)中表达的重组蛋白和天然蛋白之间。目前已经评估了该酶的粗制备物在行业应用中的效率,在毕赤酵母中产生的重组O.piceae甾醇酯酶在实验室条件下结果最好[22],有望用于生产应用。图1显示了对不同来源的甾醇酯酶的系统发育分析。
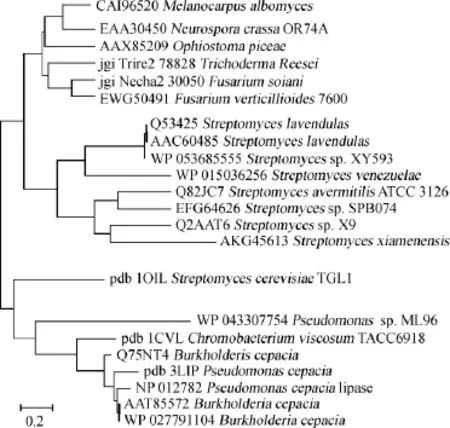
图1 不同微生物来源的甾醇酯酶的系统发育分析Fig.1 Phylogenetic analysis of sterol esterase from different microorganism's origin
2 甾醇酯酶的分离纯化及结构
2.1 甾醇酯酶的分离纯化
酶的分离纯化是研究酶的分子结构、生理功能以及应用的基础。至今国内使用的甾醇酯酶主要依赖进口,价格昂贵。究其原因,除了酶源问题外,酶的分离纯化也未从根本上得到解决。甾醇酯酶是一种疏水性较强的蛋白,容易形成聚集体,不利于酶纯化。添加表面活性剂能减轻蛋白之间的聚集作用,有助于甾醇酯酶的纯化,一般多采用胆汁酸盐或TritonX-100作为活化剂。根据文献报导[23],在以往的纯化研究中,超滤、透析、萃取、离子交换层析、凝胶层析、疏水层析、亲和层析等常规技术相继得到应用。其中,疏水层析技术显示出了极大的优越性,尤其是对于工程菌产甾醇酯酶的纯化。VAQUERO M E等[24]对在毕赤酵母(P.pastoris)中重组表达的甾醇酯酶采取单一的疏水层析(Octyl-Sepharose柱)进行纯化并取得理想的结果。不同来源的甾醇酯酶纯化步骤及相关数据见表1,目前这方面研究仍是一大热点。

表1 几种甾醇酯酶纯化步骤Table 1 Purification steps of several kinds of sterol esterase
2.2 甾醇酯酶的结构
与大部分酯水解酶一样,甾醇酯酶在结构上属于α/β水解酶家族,具有构成酶活性中心的催化三联体结构(丝氨酸Ser、组氨酸His和天冬氨酸Asp/谷氨酸Glu)[25]。在一级结构上,甾醇酯酶除了三联体氨基酸序列和“GXSXG”的五肽保守序列外,不同微生物的甾醇酯酶氨基酸序列同源性较低[26];在二级结构上,α/β水解酶折叠结构为酶的活性位点提供了一个稳定的支架,不同甾醇酯酶的二级结构的差异体现在α-螺旋和β-折叠数量、β-折叠扭曲的角度、α-螺旋的空间分布位置等;在三级结构上,脂肪酶分子在酶催化活性中心的上方,有一个α-螺旋形成的盖子(lid)结构,而酯酶分子的α-螺旋远离活性中心的上方。这一结构差异正好与“脂肪酶具有界面激活而酯酶却不具界面激活”的催化性质相吻合。JAEGER K E等[27]详细阐述了酶催化酯键断裂的过程,将其分为4个阶段:结合底物,形成瞬间的中间态,形成共价中间体以及酰基释放。因此,催化胆固醇酯水解的机制类似于胰凝乳蛋白酶那样的丝氨酸蛋白酶,这是典型的酯酶的特点。
XIANG H等[28]研究发现,来自链霉菌的甾醇酯酶缺失了“GXSXG”的保守五肽序列,催化活性中心的三联体结构中缺乏保守的组氨酸(His),但是存在两个保守的半胱氨酸(Cys)残基,可形成二硫桥以稳定三级结构。同时它也不受丝氨酸蛋白酶抑制剂苯甲基磺酰氟(phenylmethanesulfonyl fluoride,PMSF)的影响。由此推断,放线菌的甾醇酯酶与已知的酯酶相关性很小,可能存在不同的催化水解机制。
3 甾醇酯酶的克隆表达
随着分子生物学的发展,不同来源的微生物甾醇酯酶相继被克隆,并在大肠杆菌(E.coli)、毕赤酵母(P.pastoris)、克鲁维酵母(Kluyveromyces lactis)、曲霉(Aspergillussp.)等宿主中表达[21]。
XIANG H等[28]利用反转录聚合酶链反应(reverse tran scriptase-polymerase chain reaction,RT-PCR)技术克隆了来自链霉菌(Streptomycessp.)X9的完整的甾醇酯酶基因片段,共1 336 bp,其中开放阅读框(open reading frame,ORF)包括654 bp,编码217个氨基酸残基,分子质量为18.8 ku。在起始密码子上游5 bp处有Shine-Dalgarno序列(AAGGAGG),前38个氨基酸为信号肽部分,与来自阿维链霉菌(S.avermitilis)JCM5070的甾醇酯酶基因有64%的同源性。CALERO-RUEDA O等[19]将从线嘴壳菌O.piceaeCECT 20416的甾醇酯酶基因克隆并在毕赤酵母(P.pastoris)GS115中进行异源表达,分别以pGEM-T Easy和pPIC9质粒(Invitrogen TM AOX 1启动子)作为克隆载体和表达载体,获得具有较高甾醇酯酶活性的重组蛋白。该酯酶基因的DNA序列和推导的氨基酸序列在GenBank/EBI数据库中的编号为AY899847。该基因的ORF为1647bp,不含内含子序列,编码549个氨基酸,分子质量为58.2 ku,前12个氨基酸为信号肽部分,其N末端的氨基酸序列与之前从野生菌株中纯化获得的蛋白质的氨基酸序列一致。
KONTKANEN H等[29]将嗜热真菌Melanocarpus albomycesVTTD-96490的甾醇酯酶STE1转入pHAK4/77,在里氏木霉(T.reesei)中进行表达(纤维二糖水解酶启动子),通过凝胶过滤层析,测得其分子质量为120 ku,以二聚体形式存在,而野生菌株中的甾醇酯酶以四聚体形式存在,虽然两者都有5%的N连接的糖基化,但是两者的糖基化模式可能有轻微不同。氨基酸序列显示其重组SET1疏水性氨基酸残基达41.1%。推测重组STE1内部隐藏的C端His标签能降低疏水性,导致二聚体结构的形成。
4 微生物甾醇酯酶的应用
4.1 临床上测定胆固醇含量
心脑血管疾病是危害人类健康的一种常见疾病[30],而胆固醇含量是诊断和预防心脑血管疾病的重要指标。目前,临床上已经形成了多种血液胆固醇的检测方法(化学法、酶法、同位素稀释法、质谱分析法等)[31],其中,胆固醇的酶法检测是一种重要的方法。
自ALLAIN C C等[32]于1974年首次提出胆固醇的的酶法检测后,胆固醇酯酶的实际应用正式被开创。检测原理是:胆固醇酯依次在胆固醇酯酶(cholesterol esterase,COE)和胆固醇氧化酶(cholesteroloxidase,COD)的作用下生成脂肪酸,△4-胆甾烯(cholest-4-en-3-one)和H2O2。常规酶法检测是通过用Tninder显色系统检测终点产物H2O2含量(波长500 nm处用比色法检测所得的吸光度值与样品中的总胆固醇浓度对应的线性关系,可以通过预先制作的标准曲线求出对应的总胆固醇浓度)来计算胆固醇酯含量。具体反应如下:

随着科学的发展,对常规酶法检测不断改进,出现了许多新方法(如胆固醇脱氢酶法、荧光法、电极法、热敏电阻法等)[33]。虽然方法不断改进,但基本原理不变,反应第一步都需要胆固醇酯酶的参与,这使胆固醇酯酶在测定胆固醇含量上起着不可替代的作用。因此研发高质量的胆固醇酯酶制剂具有重要意义。
4.2 酶法合成植物甾醇酯
植物甾醇是一类普遍存在于植物体中的微量活性成分,具有降低血液中胆固醇、防治心血管疾病、抗氧化等功效,被誉为“生命的钥匙”[34-35]。2010年,我国卫生部批准植物甾醇和甾烷醇作为新资源食品[36],植物甾醇成为功能性食品研发的热点。但是,植物甾醇在油相中溶解度低,在食品体系中添加时经常会出现结晶现象,稳定性也不及甾醇酯,因此考虑用将植物甾醇改性转变成甾醇酯的方式添加到食品中,制成功能性食品。其主要合成方法有化学法和酶法,酶法具有合成条件温和、副产物少、安全性高等优点,因此具有很好的应用前景。罗日明[37]通过酶法催化合成的甾醇酯(肉豆蔻酸植物甾醇酯和月桂酸植物甾醇酯)在油脂中的溶解度比植物甾醇提高了3~7倍。且有研究显示各类甾醇酯均具有良好的降胆固醇功能[36]。
4.3 制浆造纸工业,解决树脂障碍
造纸工业术语“树脂障碍”中的树脂主要指木材原料中各种天然的疏水性有机物质,包括甘油三酯、甾醇酯、树脂酸等[38]。在造纸生产过程中,这些化合物可以单独或者与不溶性填料、无机盐和涂布剂等作用产生沉积,严重影响造纸产品的质量及产量,被统称为“树脂障碍”。早在20世纪90年代初,日本Juji公司率先提出用酶法水解甘油三酯减少树脂障碍。目前已将一些商业化的酶制剂(如resinase)用于解决这一问题,但是这些酶制剂缺乏甾醇酯酶活性,不能水解木材甾醇酯。因此,提出将甾醇酯酶与脂肪酶结合使用的设想,共同解决树脂障碍问题,以期生产出稳定性和韧性更好的纸张。CALERO-RUEDA O等[19]最先使用由从线嘴壳菌(O.piceae)分泌的甾醇酯酶降解木浆中的甾醇酯,并取得理想效果。叶聿程等[39]以天然树脂为唯一碳源筛选出一株降解树脂能力较强的粉尘肠杆菌(Enterobacter pulveris)Z57,降解率为6%。ZENG C[40]以植物甾醇酯为唯一碳源,从土壤中筛选出一株高产甾醇酯酶的金黄杆菌(Chryseobacteriumsp.)jx-39,其甾醇酯酶对不同碳链长度的脂肪酸胆固醇酯和对硝基苯酚酯具有广泛的底物特异性,显示该酶具有解决树脂障碍的潜力。
除此之外,甾醇酯酶也可作为清洁催化剂应用于化工、纺织等行业,具有很大的应用潜力,因此挖掘稳定的、催化效率高的甾醇酯酶在实际的工业化生产中尤其重要。
5展望
目前研究报道的甾醇酯酶活力较低,底物专一性范围较宽,它们中的大多数也可以同时水解不同长度的脂肪酸链,这并不利于专一性地水解甾醇酯,因此限制了该酶的广泛应用。科研工作者不断努力筛选底物特异性强的高产甾醇酯酶菌株、优化提高甾醇酯酶的生产水平且降低生产成本。除此之外,随着分子生物学、基因组学、蛋白质组学等学科的发展,提高甾醇酯酶的异源表达水平,改善甾醇酯酶的酶学性质,研究甾醇酯酶与底物的作用机理将是今后的科研热点和趋势。这将为甾醇酯酶在不同领域的开发应用奠定基础。
[1]熊缨,李伯良.人细胞内胆固醇酯化酶ACAT[J].生命的化学,2014,34(3):337-345.
[2]徐毅,陈晨,吴小梅.立体选择性羧酸酯酶产生菌的筛选及发酵优化[J].上海应用技术学院学报:自然科学版,2016,16(2):112-116.
[3]黄泓轲,罗健玮.胆固醇测定生物传感器研究进展[J].科技资讯,2015,13(3):234-234.
[4]李云泽.生物酶控制树脂技术在木片磨木浆车间的应用[J].华东纸业,2014,45(3):11-14.
[5]HASAN F,SHAH A A,HAMEED A.Industrial applications of microbial lipases[J].Enzyme Microb Tech,2006,39(2):235-251.
[6]VAQUERO M E,BARRIUSO J,MARTÍNEZ M J,et al.Properties, structure,and applications of microbial sterol esterases[J].Appl Microbiol Biotechnol,2016,100(5):2047-2061.
[7]UWAJIMA T,TERADA O.Studies on sterol metabolism by microorganisms.III.Purification and properties of extracellular cholesterol ester hydrolase ofPseudomonas fluorescens[J].Agr Biol Chem,1975,39(7): 1511-1512.
[8]PETERS J,ONGURI V,NISHIMOTO S K,et al.TheChlamydia trachomatisCT149 protein exhibits esterase activityin vitroand catalyzes cholesteryl ester hydrolysis when expressed in HeLa cells[J].Microbes Infect,2012,14(13):1196-1204.
[9]SØE J B,JØRGENSEN T L.Method for producing phytosterol/phy-tostanol phospholipid esters:EP20100713521[P].2012-02-01.
[10]BROWN A W,HANG J L,DUSSAULT P H,et al.Plant sterol and stanol substrate specificity of pancreatic cholesterol esterase[J].J Nutr Biochem,2010,21(8):736-740.
[11]NISHIMURA M,SUGIYAMA M.Cloning and sequence analysis of a Streptomyces cholesterolesterase gene[J].Appl Microbiol Biotechnol, 1994,41(4):419-424.
[12]SUGIHARA A,SHIMADA Y,NOMURA A,et al.Purification and characterization of a novel cholesterol esterase fromPseudomonas aeruginosa,with its application to cleaning lipid-stained contact lenses [J].Biosci Biotechnol Biochem,2014,66(11):2347-2355.
[13]TAKEDA Y,AONO R,DOUKYU N.Purification,characterization,and molecular cloning of organic-solvent-tolerant cholesterol esterase from cyclohexane-tolerantBurkholderia cepaciastrain ST-200[J].Extremophiles,2006,10(4):269-277.
[14]杨光礼,陈壁佩,张一闵,等.假单胞菌胆固醇酯酶的生产Ⅰ.菌种筛选和发酵条件的研究[J].中国医药工业杂志,1987,18(3):17-20.
[15]黄武华,方定一.假单胞菌1299产胆固醇酯酶的研究I.适宜的产酶条件[J].微生物学通报,1990,1(5):274-277.
[16]谭晓晶.Rhodococcussp.胆固醇酯酶分离纯化与基本性质研究[D].成都:四川大学,2006.
[17]曾诚,范远鑫,丁少军.甾醇酯酶菌株Chryseobacteriumsp.的筛选及酶学性质研究[J].林产化学与工业,2013,33(3):43-49.
[18]KAMEI T,SUZUKI H,MATSUZAKI M,et al.Cholesterol esterase produced byStreptomyces lavendulae[J].Chem Pharm Bull,1978,25 (12):3190-3197.
[19]CALERO-RUEDA O,PLOU F J,BALLESTEROS A,et al.Production, isolationandcharacterizationofasterolesterasefromOphiostomapiceae [J].Biochim Et Bioph Acta,2002,1599(1-2):28-35.
[20]CEDILLOVB,PLOUFJ,MARTÍNEZMJ.Recombinant sterol esterase fromOphiostoma piceae:an improved biocatalyst expressed inPichia pastoris[J].Microb Cell Factories,2012,11(1):73-86.
[21]VAQUERO M E,BARRIUSO J,MEDRANO F J,et al.Heterologous expression of a fungal sterol esterase/lipase in different hosts:Effect on solubility,glycosylation and production[J].J Biosci Bioeng,2015,120 (6):637-643.
[22]SALAS F D,BARRIUSO J.Quorum-sensing mechanisms mediated by farnesol inOphiostoma piceae:effect on secretion of sterol esterase[J]. Appl Environ Microbiol,2015,81(13):4351-4357.
[23]刘祥义,贾宇,王雪.疏水层析技术及应用[J].煤炭与化工,2012,35(9):68-69.
[24]VAQUERO M E,PRIETO A,BARRIUSO J,et al.Expression and properties of three novel fungal lipases/sterol esterases predicted in silico:comparison with other enzymes of theCandida rugosa-like family [J].Appl Microbiol Biotechnol,2015,99(23):10057-10067.
[25]何勇锦,周有彩,季潇炜,等.脂肪酶A的研究进展[J].食品与发酵工业,2016,42(11):255-261.
[26]VÍCTOR B C,ALICI A P,JES U′S M M.Potential ofOphiostoma piceaesterolesterase for biotechnologically relevant hydrolysis reactions[J].Bioengineered,2013,4(4):249-253.
[27]JAEGER K E,REETZ M T.Microbial lipases form versatile tools for biotechnology[J].Trends Biotechnol,1998,16(9):396-403.
[28]XIANG H Y,MASUO S,HOSHINO T,et al.Novel family of cholesterolesterasesproducedbyactinomycetesbacteria[J].Biochim Et Bioph Acta,2007,1774(1):112-120.
[29]KONTKANEN H,TENKANEN M,REINIKAINEN T.Purification and characterisation of a novel steryl esterase fromMelanocarpus albomyces [J].Enzyme Microb Tech,2006,39(2):265-273.
[30]孟磊,谭艳美,袁中华.中性胆固醇酯水解酶与动脉粥样硬化[J].中南医学科学杂志,2015,43(2):217-221.
[31]郭玮,陈方俊,潘柏申.胆固醇检测方法的发展历史及标准化[J].检验医学,2013,28(11):970-974.
[32]ALLAIN C C,POON L S,CHAN C S,et al.Enzymatic determination of total serum cholesterol[J].Clin Chem,1974,20(4):470-475.
[33]武翠玲,谭晓晶,姚明明,等.胆固醇酯酶高产菌株的筛选及动力学性质研究[J].中国酿造,2006,25(12):36-39.
[34]张蕾.植物甾醇及制品降心脑血管疾病风险的功能性评价[D].天津:天津商业大学,2014.
[35]朱婧,史云洁,王颜,等.含植物甾醇酯牛奶对高胆固醇血症患者血脂水平影响的随机对照研究[J].卫生研究,2016,49(5):718-724.
[36]程谦伟,孟陆丽,冯礼,等.响应面法优化植物甾醇硬脂酸酯制备工艺[J].食品研究与开发,2016,37(6):128-131.
[37]罗日明.脂肪酶催化合成植物甾醇酯的研究[D].广州:华南理工大学,2013.
[38]王恒,叶聿程,李祖巍,等.微生物法控制马尾松树脂障碍[J].华东纸业,2014,45(1):28-31.
[39]叶聿程,李祖巍,张向明,等.一株降解树脂细菌的鉴定及发酵条件的优化[J].华东纸业,2013,44(1):36-44.
[40]ZENG C,FAN Y X,DING S J.Screening of sterol esterase-producing strainChryseobacteriumsp.and its emzymatic properties[J].Chem Ind Forest Prod,2013,33(3):43-49.
Research advance in sterol esterase from microorganisms
REN Nannan1,2,WANG Xiaohui1,2,CHI Naiyu1,2,QIAO Hui1,2,ZHANG Qingfang1,2*
(1.College of Life Science and Biotechnology,Dalian University,Dalian 116622,China;
2
.Liaoning Technology of Marine Microbiological Engineering Research Center,Dalian 116622,China)
Sterol esterase(EC 3.1.1.13),as a high activity biocatalyst,has extremely broad application value in medical detection,food,paper industry and environmental protection,etc.The research advance of microorganism's origin,separation and purification,cloning and expression,three-dimensional structure and application of sterol esterase were summarized,which might lay foundation for research,development and industrialization application of sterol esterase.
sterol esterase;microorganism;cloning and expression;separation and purification;application
Q556.1
0254-5071(2017)06-0009-05
10.11882/j.issn.0254-5071.2017.06.002
2017-05-12
国家高技术研究发展计划‘863计划’项目(2007AA021306)
任楠楠(1992-),女,硕士研究生,研究方向为微生物与酶工程。
*通讯作者:张庆芳(1965-),女,教授,博士,研究方向为微生物与酶工程。
