基于染色体核型的玉米地方品种聚类分析
2017-07-05姚启伦陈发波何章帅
姚启伦,陈发波*,何章帅,方 平
(1.长江师范学院 生命科学与技术学院,重庆 408100; 2.长江师范学院 武陵山片区绿色发展协同创新中心,重庆 408100)
基于染色体核型的玉米地方品种聚类分析
姚启伦1,2,陈发波1,2*,何章帅1,方 平1
(1.长江师范学院 生命科学与技术学院,重庆 408100; 2.长江师范学院 武陵山片区绿色发展协同创新中心,重庆 408100)
以40个西南地区玉米地方品种为材料,通过细胞学鉴定分析其主要核型参数,并采用最长距离法对其进行聚类分析。结果表明,40个玉米地方品种的全组染色体长度、最长染色体长度、最短染色体长度、长度比、平均着丝点指数、平均臂比和核不对称系数平均值分别为89.56 μm、5.84 μm、2.64 μm、2.46、35.62%、1.48和59.06%,其对应变幅分别为61.51~103.82 μm、3.62~7.50 μm、0.98~4.33 μm、1.41~5.21、26.93%~40.33%、1.05~2.43和51.20%~70.90%,其中,17份玉米地方品种存在B染色体;40个玉米地方品种的平均遗传距离为10.05,变幅为3.09~46.57。基于7个核型参数可将40个玉米地方品种在遗传距离15.98处划分为6类,第Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ类分别包含3、9、4、14、8、2个品种。40个玉米地方品种供试材料间存在较大的核型差异,这在细胞水平上表明西南地区玉米地方品种具有丰富的遗传变异。
玉米地方品种; 核型; 聚类分析
玉米地方品种是在适应当地变化多端的气候环境下,经过长期人工选择和栽培驯化形成的品种群体,其中蕴藏着丰富的遗传变异[1],是重要的育种基础材料。前人[1-5]已在群体、个体、细胞和分子水平对玉米地方品种的遗传多样性进行了深入研究,但鲜见有对其核型研究的报道。染色体是遗传物质的载体,由于染色体数目和形态结构在物种中的相对稳定性,不同物种和同一物种的不同品种存在染色体核型差异,因而,染色体核型可作为区分和聚类品种的依据[6-7]。
玉米体细胞内除有10对正常染色体(A染色体)外,也可能有超过正常染色体数目的B染色体存在,已有研究表明,B染色体在玉米群体、个体和细胞中的分布频率有极大差异,从而对染色体核型有一定影响[8-10]。本研究以40个玉米地方品种为材料,通过对染色体鉴定和核型分析,利用主要核型参数对玉米地方品种进行遗传多样性分析,以期为玉米地方品种的分类及利用提供依据。
1 材料和方法
1.1 供试材料
供试材料为西南地区40个不同类型的玉米地方品种,由长江师范学院玉米育种课题组提供。材料编号、采集地及籽粒特征见表1。
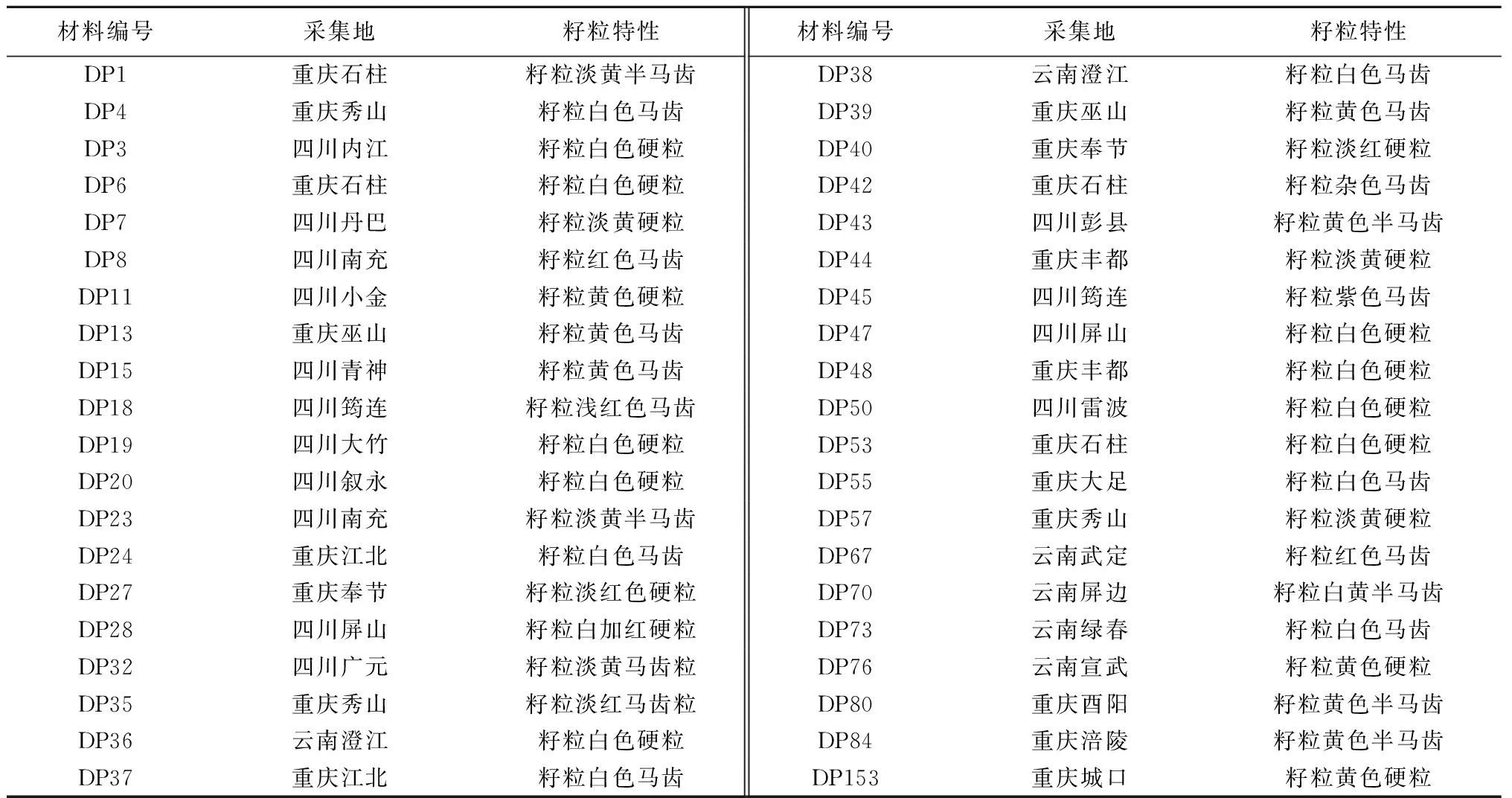
表1 玉米地方品种编号、采集地及籽粒特征
1.2 试验方法
1.2.1 浸种催芽 室温下将40个玉米地方品种的种子分别浸种1~2 h,置于25 ℃ 恒温培养箱中培养,待种子胚根长至1.5~2 cm,分别于8:00和14:00取材,此时中期分裂相细胞较多,便于染色体形态的观察。
1.2.2 预处理 将清洗干净并吸干水分的根尖放入α-溴萘饱和溶液,在4 ℃低温条件下暗处理3.5 h。
1.2.3 固定 将预处理后的根尖转移至培养皿,清洗根尖并吸干水分,置入盛有固定液(V甲醇或乙醇∶V冰乙酸=3∶1)的4 mL离心管中,在4 ℃ 低温条件下固定24 h左右。
1.2.4 解离 切取根尖分生组织,置入盛有酶液(2%纤维素酶∶2%果胶酶=1∶1)的0.2 mL离心管内,在35 ℃ 恒温培养箱中酶解4 h,在凹玻皿中用蒸馏水吹洗酶解后的根尖2~3次,最后用去离子水低渗处理根尖30 min。
1.2.5 制片 将根尖置于洁净载玻片上,滴加卡宝品红染色液,载玻片置于酒精灯上微热处理1 min,最后进行压片操作。
1.2.6 镜检及拍照 在光学显微镜下观察、鉴定单细胞染色体,拍照记录细胞染色体中期分裂相图像,用作染色体核型分析。
1.3 数据统计与分析
参照李懋学等[11]的方法,测量玉米地方品种各染色体的长度,进行核型分析。参照刘慧民等[12]和李梦寒等[13]的方法,选取染色体全长、最长染色体长度、最短染色体长度(包括B染色体)、长度比(最长染色体/最短染色体)、平均臂比(臂比=长臂长度/短臂长度)、平均着丝点指数(着丝点指数=染色体的短臂长度/染色体全长)和核不对称系数(长臂总长/全组染色体长度)7个核型参数,利用DPS软件,进行标准化转化,采用欧氏距离,利用最长距离法对40个玉米地方品种进行聚类分析。
2 结果与分析
2.1 玉米地方品种的核型分析
玉米地方品种染色体数目为2n=2x=20+xB(图1)。40个玉米地方品种的主要核型参数见表2。从表2可知,40个玉米地方品种全组染色体总长度介于61.51~103.82 μm,平均值为89.56 μm。最长染色体长度介于3.62~7.50 μm,平均值为5.84 μm。最短染色体长度介于0.98~4.33 μm,平均值为2.64 μm。长度比介于1.41~5.21,平均值为2.46。平均着丝点指数介于26.93%~40.33%,平均值为35.62%。平均臂比介于1.05~2.43,平均值为1.48。核不对称系数介于51.20%~70.90%,平均值为59.06%。由此可知,40份供试材料的各核型参数变幅大。40个玉米地方品种中有17个品种含B染色体,分别为DP1、DP3、DP6、DP7、DP11、DP13、DP15、DP28、DP35、DP37、DP39、DP40、DP47、DP55、DP57、DP73和DP84,其中有2个品种含2条B染色体,分别为DP35和DP73,所含B染色体的玉米地方品种在40份供试材料中所占的比例为42.5%,含2条B染色体的占5%。B染色体出现频率较大。
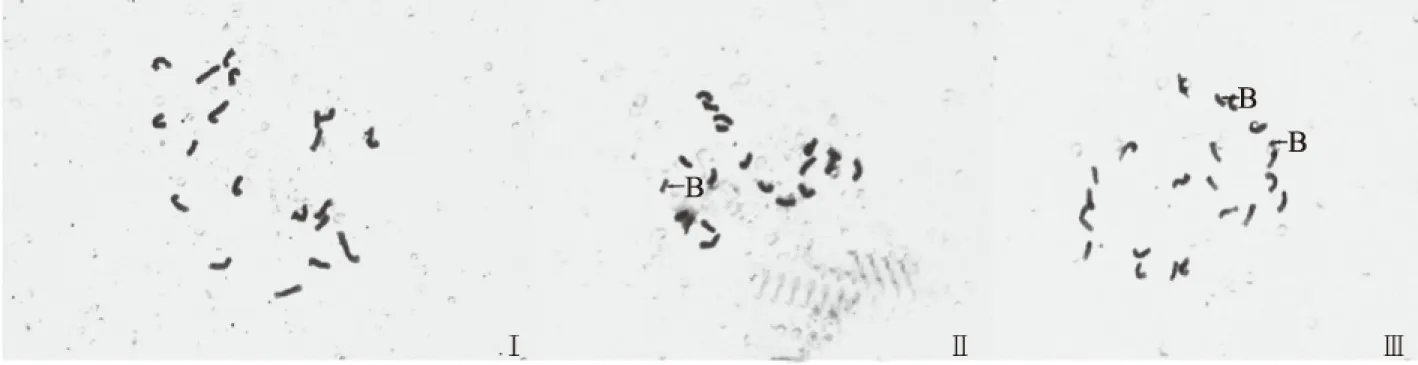
Ⅰ.DP42,含有0条B染色体;Ⅱ.DP15,含有1条B染色体;Ⅲ.DP73,含有2条B染色体
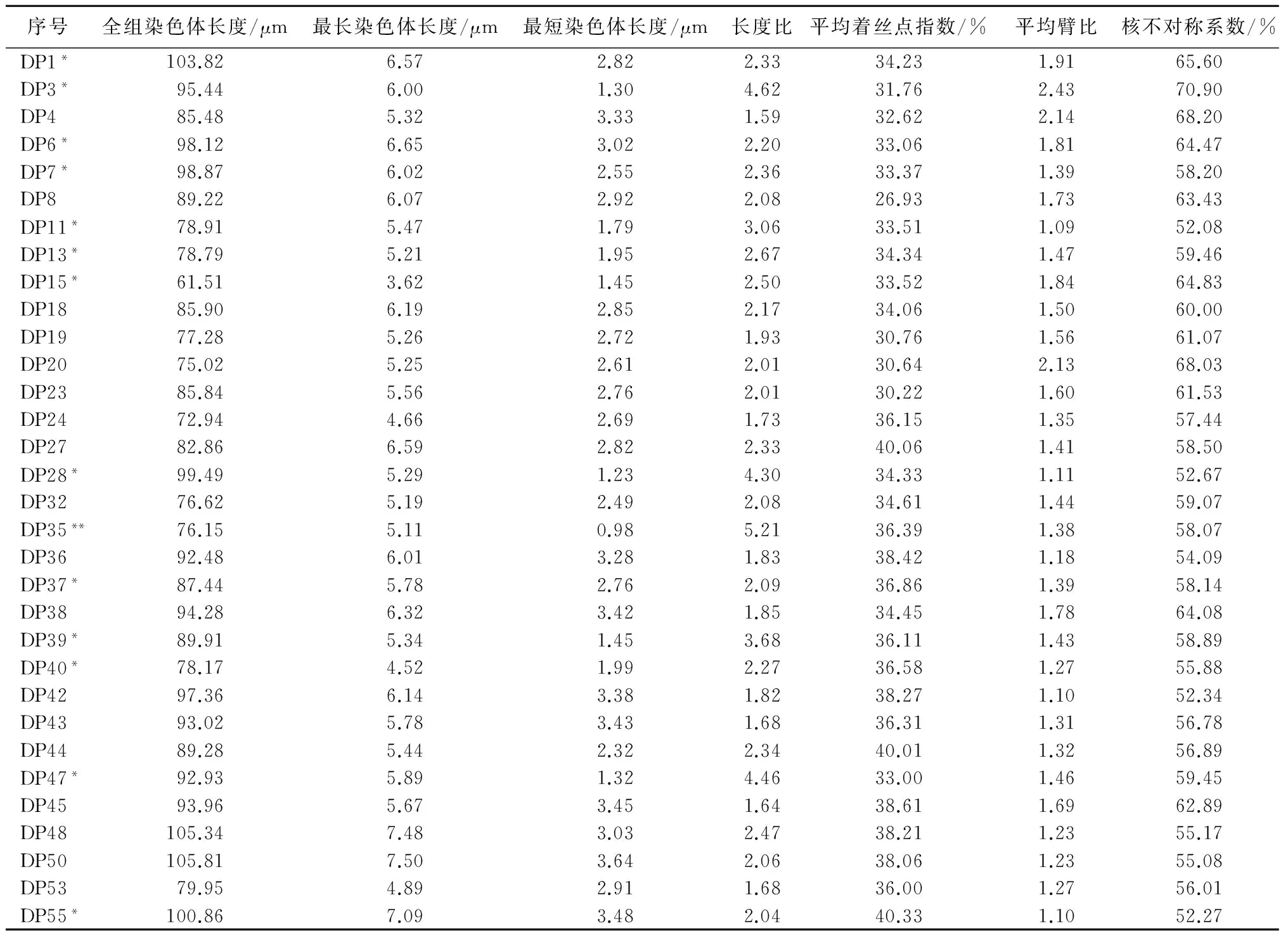
序号全组染色体长度/μm最长染色体长度/μm最短染色体长度/μm长度比平均着丝点指数/%平均臂比核不对称系数/%DP1*103.826.572.822.3334.231.9165.60DP3*95.446.001.304.6231.762.4370.90DP485.485.323.331.5932.622.1468.20DP6*98.126.653.022.2033.061.8164.47DP7*98.876.022.552.3633.371.3958.20DP889.226.072.922.0826.931.7363.43DP11*78.915.471.793.0633.511.0952.08DP13*78.795.211.952.6734.341.4759.46DP15*61.513.621.452.5033.521.8464.83DP1885.906.192.852.1734.061.5060.00DP1977.285.262.721.9330.761.5661.07DP2075.025.252.612.0130.642.1368.03DP2385.845.562.762.0130.221.6061.53DP2472.944.662.691.7336.151.3557.44DP2782.866.592.822.3340.061.4158.50DP28*99.495.291.234.3034.331.1152.67DP3276.625.192.492.0834.611.4459.07DP35**76.155.110.985.2136.391.3858.07DP3692.486.013.281.8338.421.1854.09DP37*87.445.782.762.0936.861.3958.14DP3894.286.323.421.8534.451.7864.08DP39*89.915.341.453.6836.111.4358.89DP40*78.174.521.992.2736.581.2755.88DP4297.366.143.381.8238.271.1052.34DP4393.025.783.431.6836.311.3156.78DP4489.285.442.322.3440.011.3256.89DP47*92.935.891.324.4633.001.4659.45DP4593.965.673.451.6438.611.6962.89DP48105.347.483.032.4738.211.2355.17DP50105.817.503.642.0638.061.2355.08DP5379.954.892.911.6836.001.2756.01DP55*100.867.093.482.0440.331.1052.27
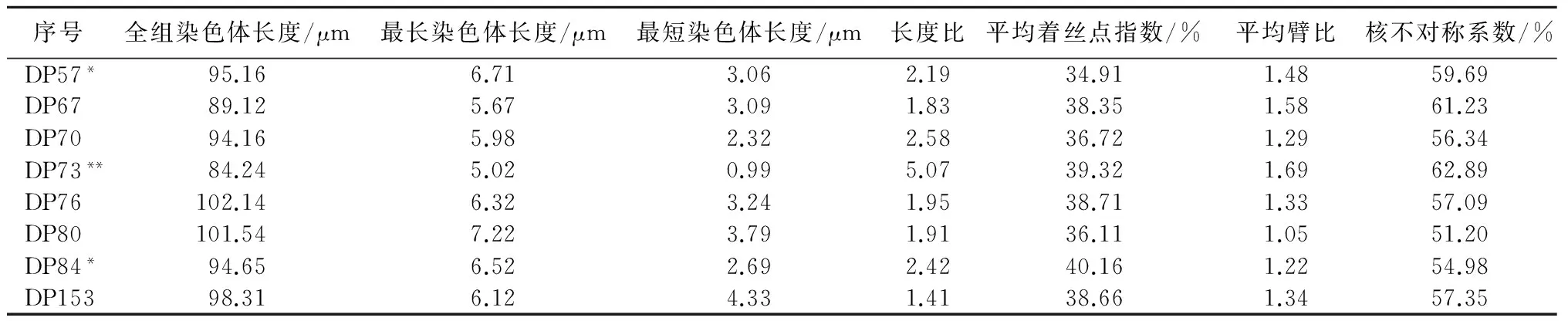
续表2 40个玉米地方品种的主要核型参数
注: *表示该品种存在1条B染色体,**表示该品种存在2条B染色体。
2.2 玉米地方品种的遗传距离及分布频率分析
基于核型参数,利用DPS软件进行标准化转化,采用欧氏距离,计算各品种间的遗传距离,40个玉米地方品种间遗传距离变幅为3.09~46.57,平均距离为10.05,遗传距离变化范围较大。其中DP32和DP13间的遗传距离最小,为3.09;DP11与DP1间的遗传距离最大,为46.57。从遗传距离分布情况(图2)可知,遗传距离在3.09~4.00的频率为7.7%,遗传距离在4.00~7.00的频率为35.9%,遗传距离在7.00~10.00的频率为25.6%,遗传距离在14.00~19.00的频率和19.00~26.00的频率都为7.7%,遗传距离在26.00~47.00的频率为2.6%。可以看出,遗传距离在4.00~10.00的分布较多,占61.5%,其中有92.3%的遗传距离大于4.00,说明40份供试材料间存在一定的遗传多样性。
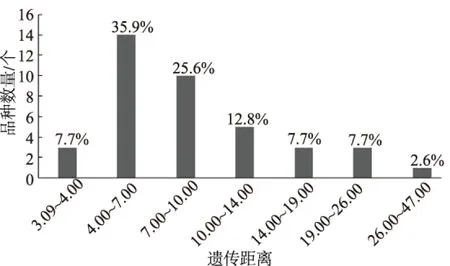
图2 40个玉米地方品种遗传距离频率分布
2.3 基于核型参数的玉米地方品种聚类分析
基于全组染色体长度、最长染色体长度、最短染色体长度、长度比、平均臂比、平均着丝点指数和核型不对称系数7个核型参数,对玉米地方品种进行聚类分析(图3),在遗传距离15.98处可以将40个玉米地方品种分为6类。第Ⅰ类包含DP1、DP3和DP6 3个品种,占总供试材料的7.5%,均含有B染色,核型不对称系数在64.47%~70.90%,平均臂比在1.81~2.43,籽粒都为马齿型;第Ⅱ类包含9个品种,分别为DP7、DP76、DP153、DP28、DP80、DP42、DP55、DP48和DP50,占总供试材料的22.5%,核型不对称系数在51.20%~58.20%,平均臂比在1.05~1.39,最长染色体长度介于5.29~7.50 μm,其中,DP7、DP28和DP55含有B染色体;第Ⅲ类共4个品种,分别为DP4、DP18、DP23和DP8,占总供试材料的10.0%,这类中最明显的特征是都不含B染色体,全组染色体长度在85.48~89.22 μm,平均臂比在1.50~2.14,核不对称系数介于60.00%~68.20%,籽粒都为马齿型或半马齿型;第Ⅳ类共含有14个玉米地方品种DP27、DP73、DP37、DP39、DP44、DP67、DP36、DP43、DP70、DP84、DP38、DP45、DP47和DP57,占总供试材料的35%,这类平均臂比介于1.18~1.78,核型不对称系数介于54.09%~64.08%,其中,有5个玉米地方品种含有B染色体,DP73含有2条B染色体;DP11、DP40、DP53、DP13、DP32、DP19、DP24和DP35聚为第Ⅴ类,占总供试材料的20%,平均臂比在1.09~1.56,核型不对称系数在52.08%~61.07%,这一类含B染色体的品种出现频率为50.0%,B染色体出现频率相对较高;DP15和DP20聚为第Ⅵ类,占总数的5%,其中DP15含B染色体,平均臂比分别为1.84和2.13,核型不对称系数分别为64.83%和68.03%。
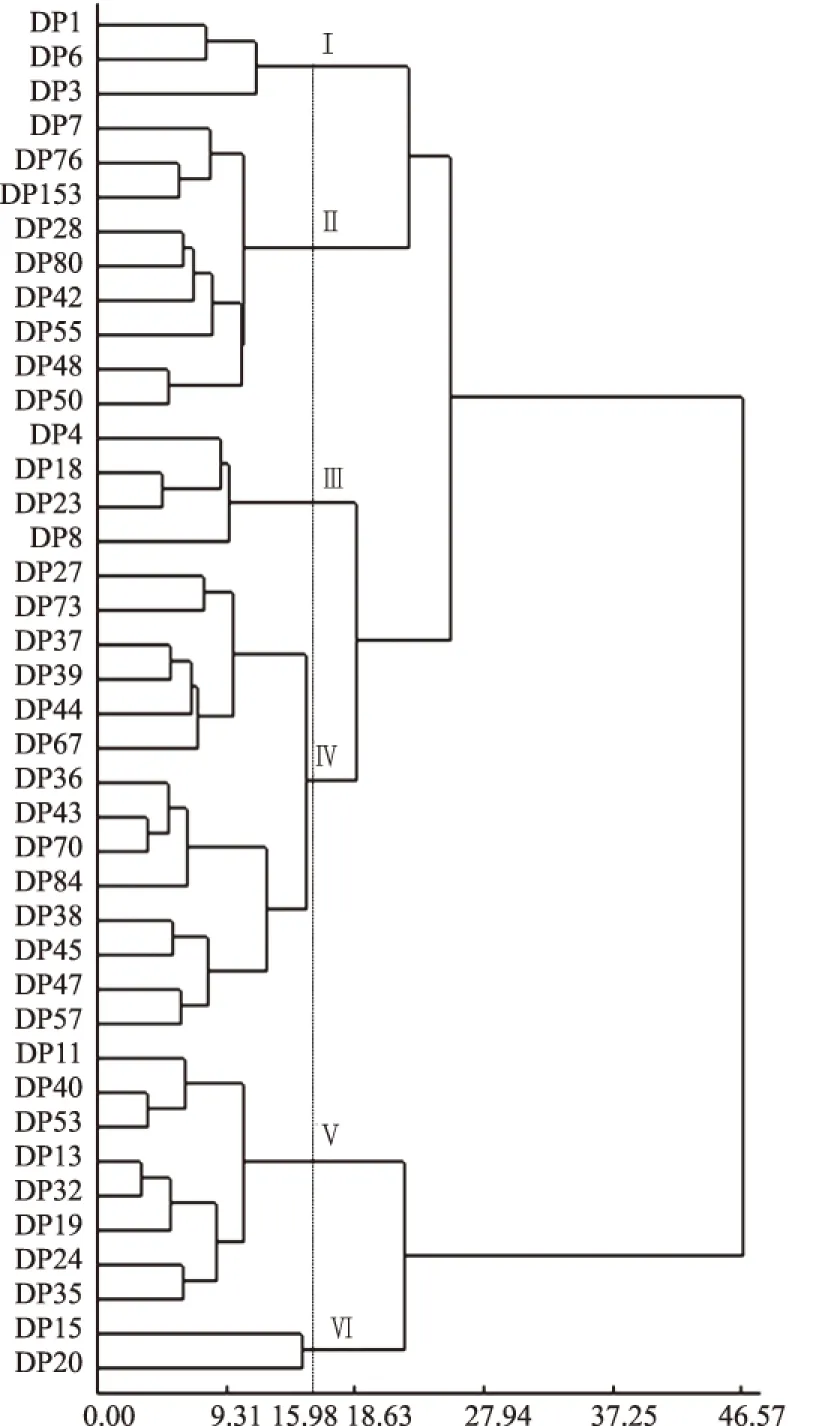
图3 基于核型参数的40个玉米地方品种聚类分析
3 结论与讨论
在作物遗传多样性和亲缘关系分析的研究中,常用的方法有分子标记、表型参数和生理生化标记等。而染色体核型是了解植物系统发育和进化的一个重要参数,可以为植物分类与鉴定提供可靠的细胞学依据[12]。张海洋等[14]选择染色体数目、染色体相对长度极差、相对长度方差、臂比值倒数均值、臂比值倒数方差、着丝点指数均值、着丝点指数方差等8个核型参数指标进行核型似近系数和进化距离分析,将14份芝麻分为2组。李璇等[15]利用相对长度、平均臂比和核型不对称系数三因子对12个泸定百合进行聚类分析,最终将12个居群分为3类。刘慧民等[12]认为,核型参数的重要性排序依次为核型不对称系数、平均臂比、臂比大于2的比率、染色体最长(最短)值、核型类型、着丝点指数均值。基于核型参数系统聚类,李梦寒等[13]将8个西藏白菜型黄籽油菜地方品种分为3组,马衣努尔姑·吐地等[16]将24个新疆野苹果分为3 类。但鲜见基于核型参数的玉米亲缘关系研究。本研究结果表明,选用的染色体全长、最长染色体长度、最短染色体长度、长度比、平均臂比、平均着丝点指数和核型不对称系数7个核型参数在不同玉米地方品种间均存在较大的差异,系统聚类将40个玉米地方品种分为6类,各类之间存在明显的差异。说明核型参数可以作为重要的细胞学标记,用于玉米地方品种分类、鉴定及遗传多样性研究。
核型分析表明,含B染色体的品种占供试材料的42.5%,西南地区玉米地方品种B染色体出现频率较高。各核型参数的变化范围较大,其中核不对称系数介于51.20%~70.90%,表明供试材料进化程度参差不齐。玉米地方品种是玉米传入我国后,经过长期地自然选择和人工培养驯化而形成的适应当地复杂环境的品种群体,加之农民在自留种过程中选择的标准不同,使玉米地方品种进化程度参差不齐[1]。遗传距离分析表明,供试玉米地方品种间遗传距离变幅为3.09~46.57,平均距离为10.05,其中有92.3%的玉米地方品种遗传距离大于4.00,聚类分析将40个玉米地方品种分为6类,且6类各具特色,说明供试玉米地方品种间存在较丰富的遗传多样性,这与姚启伦等[4]、陈发波等[17]、吴元奇等[1]和刘志斋等[5]等对玉米地方品种遗传多样性的研究结果相同。
[1] 吴元奇,郑灵,荣廷昭.西南地区白玉米地方种质资源分布及遗传多样性[J].草业学报,2013,22(4):160-169.
[2] Li Y,Shi Y S,Cao Y S,etal.Establishment of a core collection for maize germplasm preserved in Chinese National Genebank using geographic distribution and characterization data[J].Genetic Resources and Crop Evolution,2005,51(8):845-852.
[3] Li Y,Shi Y S,Cao Y S,etal.A phenotypic diversity analysis of maize germplasm preserved in China[J].Maydica,2002,47(2):107-114.
[4] 姚启伦,陈发波,刘红芳,等.基于Glb1基因序列的西南地区玉米地方品种的系统进化[J].作物学报,2015,41(7):998-1006.
[5] 刘志斋,吴迅,李永祥,等.中国玉米地方品种种族的遗传变异评估[J].中国农业科学,2015,48(16):3101-3111.
[6] 蒋培东,李展,朱苏文,等.对生玉米和互生玉米染色体核型比较研究[J].激光生物学报,2005,14(3):197-201.
[7] 张赞平.玉米各亚种的核型研究[J].华北农学报,1995,10(3):60-67.
[8] 姚启伦,陈发波,刘红芳,等.西南地区玉米地方品种B染色体多态性分析[J].中国农业科学,2015,48(14):2697-2704.
[9] 姚启伦,姚强,方平,等.玉米地方品种和自交系B染色体的比较分析[J].江苏农业科学,2015,43(3):79-81.
[10] 姚启伦,陈发波,方平.西南地区玉米地方品种B染色体初探[J].玉米科学,2014,22(1):18-22.
[11] 李懋学,陈瑞阳.关于植物核型分析的标准化问题[J].植物科学学报,1985,3(4):297-302.
[12] 刘慧民,陈雅君,吕贵娥,等.17种绣线菊核型特征及核型参数分析[J].园艺学报,2010,37(9):1456-1462.
[13] 李梦寒,次仁白珍,安克杰,等.西藏白菜型黄籽油菜地方品种的核型分析[J].中国油料作物学报,2016,38(5):549-554.
[14] 张海洋,苗红梅,李春,等.芝麻染色体核型及似近系数分析[J].植物学报,2012,47(6):602-614.
[15] 李璇,段青,王祥宁,等.云南泸定百合12个野生居群的核型研究[J].园艺学报,2014,41(5):935-945.
[16] 马衣努尔姑·吐地,张延辉,秦伟,等.新疆野苹果的不同种下类型染色体核型分析[J].中国农业科学,2016,49(8):1540-1549
[17] 陈发波,姚启伦,聂术君,等.利用Waxy基因检测不同玉米地方品种的遗传差异[J].江苏农业科学,2014,42(4):18-21.
Cluster Analysis of Maize Landraces Based on Karyotype Parameters
YAO Qilun1,2,CHEN Fabo1,2*,HE Zhangshuai1,FANG Ping1
(1.Life Sciences and Technology Institute,Yangtze Normal University,Chongqing 408100,China; 2.Centre for Green Development and Collaborative Innovation in Wuling Mountain Region,Yangtze Normal University,Chongqing 408100,China)
Fourty maize landraces collected from Southwest China were used as materials,and their mainly karyotype parameters were identified using cytological technology,subsequently,cluster analysis of those maize landraces were conducted by the longest distance method.The results indicated that B chromosomes(Bs) existed in 17 maize landraces,and the total chromosome length,the longest chromosome length,the smallest chromosome length,the ratio of chromosome length,the mean centromere index,the mean arm ratio,as well as the index of karyotypic asymmetry of fourty maize landraces were on an average of 89.56 μm,5.84 μm,2.64 μm,2.46,35.62%,1.48 and 59.06%,respectively,and the corresponding amplitude variations were 61.51—103.82 μm,3.62—7.50 μm,0.98—4.33 μm,1.41—5.21,26.93%—40.33%,1.05—2.43 and 51.20%—70.90%,respectively.In addition,the mean genetic distance among those maize landraces was on average of 10.05,ranging from 3.09 to 46.57.Based on these seven karyotype parameters,40 maize landraces could distinctively be clustered into six groups under the genetic distance of 15.98,group Ⅰ,Ⅱ,Ⅲ,Ⅳ,Ⅴ and Ⅵ were consisted of 3,9,4,14,8 and 2 maize landraces,respectively.Great karyotype difference was found in 40 maize landraces,suggesting that abundant genetic variance existed in maize landraces at cytological level in Southwest China.
maize landraces; karyotype; cluster analysis
2016-12-14
国家自然科学基金项目(31371633);重庆市科委社会民生项目(cstc2016shmszx80030);重庆高校优秀成果转化项目(CGZH1633)
姚启伦(1964-),男,重庆万州人,教授,博士,主要从事作物遗传育种研究工作。E-mail:yql641@aliyun.com
*通讯作者:陈发波(1981-),男,四川冕宁人,副教授,博士,主要从事作物遗传育种研究工作。 E-mail:chenfabo963@126.com
S513
A
1004-3268(2017)06-0034-05
