干旱胁迫对不同种源文冠果幼苗水分生理特性及渗透调节物质的影响1)
2017-06-28赵雪张秀珍
赵雪 张秀珍
(河北农业大学,保定,071000)(青岛冠中生态股份有限公司)
牟洪香 孙晓 贺红月 杨可伟 单媛媛 李春友
(河北农业大学)
干旱胁迫对不同种源文冠果幼苗水分生理特性及渗透调节物质的影响1)
赵雪 张秀珍
(河北农业大学,保定,071000)(青岛冠中生态股份有限公司)
牟洪香 孙晓 贺红月 杨可伟 单媛媛 李春友
(河北农业大学)
以6个种源文冠果一年生幼苗作为研究对象,采用盆栽称质量控水的方法设置3个水分梯度,研究了不同种源文冠果幼苗在不同水分梯度上的水分生理特征及渗透调节变化规律,利用模糊数学隶属函数法对不同种源文冠果幼苗进行抗旱性综合评价。结果表明:随着干旱胁迫的加剧,不同种源的文冠果幼苗叶片含水量降低,渗透调节物质含量增加;通过对不同种源文冠果幼苗抗旱性的综合评价,种源间抗旱性由大到小依次为内蒙乌丹、新疆察县、山西汾阳、河北新华区、辽宁建平、河南陕州区。
干旱胁迫;文冠果;水分生理;渗透调节
水分缺乏是植物生长过程中时常遭受的逆境之一,伴随着全球气候变化,水分胁迫影响着越来越多的地区,水资源的短缺也将成为限制植物生长的主要因素[1],栽种抗旱性强的树种对干旱地区植被修复具有重要意义,为此抗旱树种的选择成为植物领域研究的热点[2]。植物应对水分胁迫的方式有两种:一方面通过形态学的变化来提高自身水分的吸收并降低水分的丧失;另一方面则是通过生理生化反应来抵抗干旱逆境[3]。研究表明,植物在水分胁迫下,通过细胞主动积累大量溶质,降低渗透势,保证植物能继续从外界吸收水分,维持细胞膨压,保证生理过程的正常进行[4]。研究水分胁迫下叶片的水分生理特征以及渗透调节物质的变化规律,对于了解植物的耐旱原因,揭示植物的耐旱机制,为干旱半干旱地区造林树种的选择都具有重要的指导意义。
文冠果(XanthocerassorbifoliaBunge.)为无患子科,文冠果属落叶灌木或小乔木,又名文官果、僧灯毛道等[5],分布北纬28°34′~47°20′,东经73°20′~120°25′[6],其果实具有较高的营养价值与药用价值[7],种子含油量高,是我国北方特有的木本油料树种[8]。以往一些学者对文冠果的研究侧重于分布规律、繁殖技术、开发利用等领域[9-12],而对于水分胁迫下文冠果的研究相对较少。本文通过盆栽控水试验,研究不同水分条件下文冠果幼苗水分生理及其渗透调节机制,采用模糊隶属函数法[13]对抗旱性进行综合评价,为干旱半干旱地区树种的选择及其植被恢复提供理论依据。
1 材料与方法
试验地位于河北省保定市河北农业大学标本园内,属暖温带半湿润大陆季风气候,年平均气温12 ℃,年平均降水529.7 mm,无霜期165~210 d。试验用土为壤土。选取辽宁省朝阳市建平县、内蒙古赤峰市翁牛特旗乌丹镇、新疆维吾尔自治区伊利哈萨克自治州伊宁市察布查尔锡伯自治县(以下简称“察县”)、河南省三门峡市陕州区、山西省吕梁市汾阳市、河北省石家庄市新华区6地种源种子繁育的文冠果幼苗,6种源采种地气候特征及地理位置见表1。2015年4月将文冠果种子用50 ℃温水进行催芽处理,待1/3种子露白时,播种于营养杯中。2015年6月将长势一致的幼苗栽植到无孔的塑料花盆(盆高25 cm,口径31 cm,底径21 cm),每盆2株幼苗,每种源各24盆。设置3个水分胁迫梯度:适宜水分(土壤含水量为14.76%~15.81%,CK);中度干旱(土壤含水量为9.49%~10.54%);重度干旱(土壤含水量为6.32%~7.38%),同种源每个水分处理8个重复。待达到预设水分含量,采用电子秤称质量法,持续干旱胁迫28 d后进行各项指标的测定。
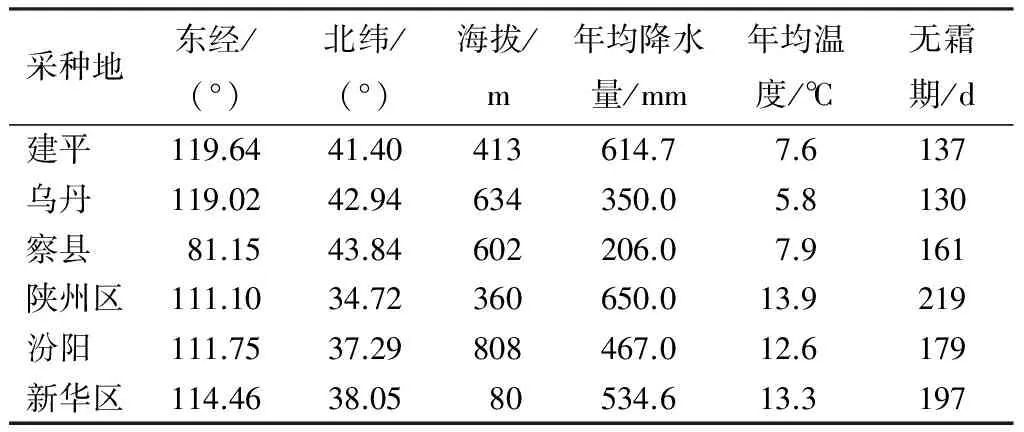
表1 文冠果采种地基本情况
1.1 测定项目及方法
叶片含水量测定采用烘干法进行测定[14];水分饱和亏测定采用称质量法[15];叶片保水力测定采用自然脱水法,用每次称质量叶片含水量占初始含水量的百分比表示;可溶性糖质量分数的测定采用蒽酮比色法[16];可溶性蛋白质质量分数的测定采用考马斯亮蓝比色法[16];K+质量分数测定采用火焰光度计法。
1.2 抗旱评价方法及数据处理与分析
试验数据处理采用Microsoft Excel 2003进行整理与计算,运用SPSS17.0进行数据分析。抗旱性综合评价采用隶属函数法,计算公式为:
U(Xi)=(Xi-Xmin)/(Xmax-Xmin),i=1、2、3、…、n;
(1)
U(Xi)=1-(Xi-Xmin)/(Xmax-Xmin),i=1、2、3、…、n。
(2)
式中,Xi指的是某一指标的测定值,Xmax与Xmin分别指的是该对应指标的最大值与最小值。若测定指标与抗旱性呈正相关关系则根据公式(1)计算隶属函数值;若测定指标与抗旱性呈负相关关系则根据公式(2)计算。
2 结果与分析
2.1 干旱胁迫对不同种源文冠果水分生理的影响
植物叶片含水量与其代谢活动有着紧密的关系[17],是反映植物忍耐干旱胁迫能力的重要指标之一[18]。表2显示随干旱程度的加深,6个种源文冠果幼苗叶片含水量都逐渐降低,但种源间下降幅度并不相同。重度干旱胁迫下,文冠果幼苗叶片含水量河南陕州区下降最大,为适宜水分状态下的8.75%;内蒙乌丹下降最小,为适宜水分状态下的5.52%。同一水分条件下,新疆察县文冠果始终能保持相对较高的叶片含水量。方差分析表明,适宜水分下,各种源间叶片含水量差异不显著;在重度干旱胁迫下,新疆察县叶片含水量显著高于辽宁建平的。
水分饱和亏是反映植物实际含水量距离饱和含水量的差值。从表2可以看出,随着水分胁迫的加重,各种源叶片水分饱和亏值逐渐增加;在重度干旱胁迫下,内蒙乌丹、河北新华区两种源水分亏缺值相对最小;方差分析表明,叶片水分饱和亏除内蒙乌丹各水分胁迫之间差异不显著外,其他种源在不同水分胁迫之间表现出显著差异;而在重度干旱胁迫下,各种源间水分亏缺值并未存在显著差异。

表2 干旱胁迫对不同种源文冠果幼苗叶片含水量及水分饱和亏的影响
注:表中数据为平均值±标准误;不同大写字母表示相同干旱胁迫下不同种源间差异显著(P<0.05);不同小写字母表示种源内不同干旱胁迫间差异显著(P<0.05)。
叶片保水力反映的是植物叶片防止水散失和组织抗脱水的能力[19]。6个种源文冠果在不同水分条件下叶片保水力如表3所示,各种源叶片保水力均在重度干旱胁迫下最高。其中,辽宁建平、新疆察县和河北新华区的文冠果叶片保水力在不同干旱胁迫下差异不大;而内蒙乌丹和河南陕州区两种源文冠果在重度干旱胁迫下,叶片保水力显著高于其他两个水分条件下的保水力。

表3 干旱胁迫下各种源文冠果幼苗叶片保水力
2.2 干旱胁迫对不同种源文冠果幼苗叶片渗透调节物质的影响
渗透调节作为植物抵抗干旱胁迫的一种重要方式,一直是抗旱生理研究的重要领域之一[20]。不同植物,甚至是同一种植物的不同品种其渗透调节能力也会存在一定的差异,大量研究表明,抗旱性强的植物渗透调节能力也较强[21]。一般认为,可溶性糖、可溶性蛋白质等物质以及K+等在渗透调节中扮演着重要的角色[22]。
可溶性糖质量分数:表4显示,随着水分胁迫的加剧,各种源叶片中可溶性糖质量分数均呈现上升趋势,且在重度干旱胁迫下达到最大值。与对照相比,可溶性糖质量分数增加幅度最大的为新疆察县(1.47倍),最小的为山西汾阳(0.67倍)。方差分析表明,不同水分条件下,各种源可溶性糖质量分数存在显著差异;在水分胁迫下,新疆察县文冠果幼苗叶片中可溶性糖质量分数显著高于其他种源。初步分析可得,可溶性糖的增加可能对提高其渗透调节有重要作用。
表4 干旱胁迫下各种源文冠果幼苗叶片可溶性糖质量分数的变化
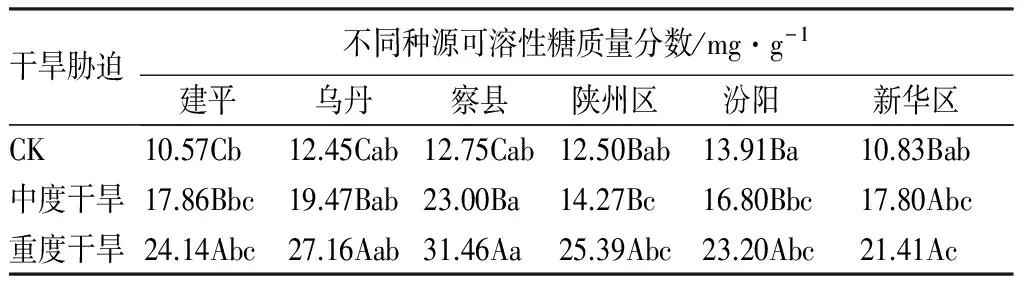
干旱胁迫不同种源可溶性糖质量分数/mg·g-1建平乌丹察县陕州区汾阳新华区CK10.57Cb 12.45Cab12.75Cab12.50Bab13.91Ba10.83Bab中度干旱17.86Bbc19.47Bab23.00Ba14.27Bc16.80Bbc17.80Abc重度干旱24.14Abc27.16Aab31.46Aa25.39Abc23.20Abc21.41Ac
注:不同大写字母表示不同干旱胁迫下的差异性(P<0.05);不同小写字母表示种源间的差异性(P<0.05)。
可溶性蛋白质质量分数:表5显示,在重度干旱胁迫下,山西汾阳和河北新华区种源叶片中可溶性蛋白质质量分数下降,其他种源可溶性蛋白质质量分数均达到最大值。重度干旱胁迫下可溶性蛋白质质量分数与对照相比,增加最多的为新疆察县种源(67.21%),最小的为山西汾阳种源(6.93%)。种源辽宁建平、内蒙乌丹、新疆察县和河南陕州区可溶性蛋白质质量分数在中度及重度干旱胁迫下差异不显著,但都与对照呈现显著差异;在中度干旱胁迫下,山西汾阳种源可溶性蛋白质质量分数显著大于辽宁建平、新疆察县、河南陕州区种源。由此可知,在中度干旱胁迫下,山西汾阳和河北新华区种源文冠果具有较高的渗透调节能力,而在重度干旱胁迫下,其渗透调节能力下降。
表5 干旱胁迫下各种源文冠果幼苗叶片可溶性蛋白质质量分数的变化

干旱胁迫不同种源可溶性蛋白质质量分数/mg·g-1建平乌丹察县陕州区汾阳新华区CK1.28Bb1.47Bb1.25Bb1.33Bb1.91Ba1.33Cb中度干旱1.80Ab2.12Aab1.89Ab1.94Ab2.30Aa2.01Aab重度干旱2.03Aab2.24Aa2.10Aa1.97Aab2.04ABab1.74Bb
注:不同大写字母表示不同干旱胁迫下的差异性(P<0.05);不同小写字母表示种源间的差异性(P<0.05)。
K+质量分数:表6显示,K+质量分数变化与可溶性蛋白质质量分数的变化基本一致。内蒙乌丹、河南陕州区和山西汾阳种源文冠果K+质量分数随着干旱胁迫的加剧,呈现出先增加后降低的趋势,但均大于对照组;重度干旱胁迫下K+质量分数与对照相比,增加最多为辽宁建平种源(62.29%),最小的为山西汾阳种源(10.72%)。方差分析表明,水分胁迫对K+质量分数表现出显著影响,两种程度干旱胁迫下K+质量分数均显著高于对照组;在中度干旱胁迫下,辽宁建平种源K+质量分数显著低于内蒙乌丹、新疆察县、山西汾阳、河北新华区种源;在重度干旱胁迫下,辽宁建平和河南陕州区种源K+质量分数显著低于其他种源。表明K+质量分数在文冠果水分胁迫下发挥着重要的渗透调节作用。
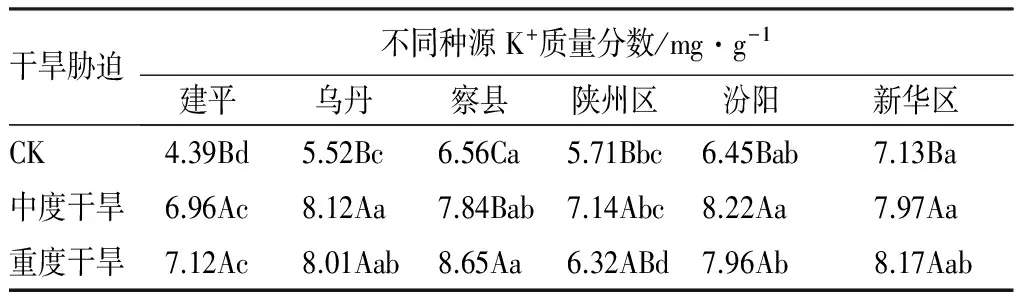
表6 干旱胁迫下各种源文冠果幼苗叶片K+质量分数的变化
注:不同大写字母表示不同干旱胁迫下的差异性(P<0.05);不同小写字母表示种源间的差异性(P<0.05)。
2.3 叶片含水量与渗透调节物质的关系
叶片含水量与3种渗透调节物质的相关性分析如表7所示。辽宁建平种源叶片含水量与可溶性糖呈显著负相关,其中新疆察县和河南陕州区种源表现出极显著负相关;辽宁建平、内蒙乌丹种源叶片含水量与可溶性蛋白质呈显著负相关;新疆察县、山西汾阳和河北新华区种源的K+质量分数与叶片含水量呈显著负相关。由此可以说明,文冠果幼苗叶片含水量与可溶性糖及K+质量分数相关性较强,二者之间存在着一定的联系;而山西汾阳、河北新华区种源叶片含水量与可溶性蛋白质、可溶性糖质量分数并未表现出显著相关性,而与K+质量分数表现出显著负相关关系。
表7 不同种源文冠果幼苗叶片含水量与可溶性糖、可溶性蛋白质、K+相关性
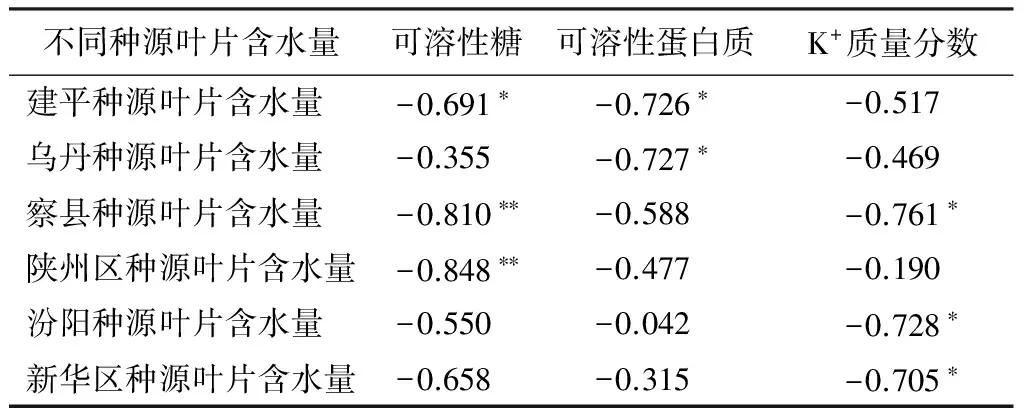
不同种源叶片含水量可溶性糖可溶性蛋白质K+质量分数建平种源叶片含水量-0.691∗-0.726∗-0.517乌丹种源叶片含水量-0.355-0.727∗-0.469察县种源叶片含水量-0.810∗∗-0.588-0.761∗陕州区种源叶片含水量-0.848∗∗-0.477-0.190汾阳种源叶片含水量-0.550-0.042-0.728∗新华区种源叶片含水量-0.658-0.315-0.705∗
注:*表示差异显著(P<0.05);** 表示差异极显著(P<0.01)。
2.4 不同种源文冠果幼苗抗旱性综合评价
基于多项指标测定的基础上,采用隶属函数综合评价的方法能提高评价的准确性[23]。对文冠果幼苗6个种源的抗旱性综合评价如表4所示,利用测定的叶片含水量、水分饱和亏、可溶性糖、可溶性蛋白质和K+这5项指标综合评价,结果发现,6个种源文冠果幼苗的抗旱性由大到小依次为内蒙乌丹、新疆察县、山西汾阳、河北新华区、辽宁建平、河南陕州区。
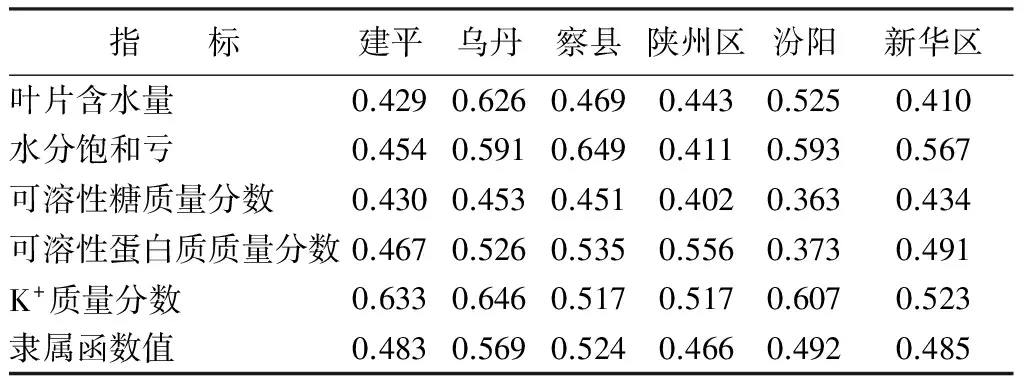
表8 不同水分条件下不同种源文冠果各指标隶属函数值
3 结论与讨论
不同水分条件下,文冠果幼苗叶片水分生理及渗透调节物质的变化,都是其对干旱环境适应性的反应。研究表明,具有较强保水能力的树种,其对干旱胁迫的耐受力也较高[24]。植物在幼苗期对干旱胁迫反应最为敏感,因此,研究幼苗期植物的抗旱能力具有一定的意义。本试验选取叶片含水量、水分饱和亏、叶片保水力及可溶性糖、可溶性蛋白质及K+质量分数各项指标,研究不同水分条件下对文冠果6个不同种源幼苗水分生理及渗透调节物质的影响,并对抗旱性进行综合评价。
植物自身水分状况与其抗旱性有着密切的关系,叶片含水量是植物体内水分状况综合指标之一[25]。本研究表明,随着干旱胁迫的加剧,各种源文冠果幼苗叶片含水量逐渐不同程度降低,水分饱和亏呈现不同程度的上升。其中新疆察县种源在重度干旱胁迫下叶片含水量最高,依据赵一鹤等[26]研究表明,在相同条件下,叶片含水量高的植物,在生理、代谢等功能上相对旺盛,其适应干旱的能力相对较强,初步可以推断新疆察县种源文冠果具有较强的抗旱性。
渗透调节是植物应对水分胁迫一种重要生理机制,通过降低水势减少水分的损失并继续从外界吸收水分,来保证各项生理活动正常进行[27]。可溶性蛋白具有较强的亲水胶体性质,能够提高植物体内细胞的亲水能力[28];可溶性糖作为植物体内重要的能源与碳源,在干旱环境下可作为小分子物质来参与渗透调节,提高植物吸水与保水能力;无机离子K+也是参与渗透调节的重要溶质。对于不同植物渗透调节物质的研究,仍存在一定的差异。王海珍等[29]研究表明,在干旱条件下,白刺花叶片内会积累大量K+和可溶性糖,来增强细胞的保水,维持膨压;王琰等[30]研究表明,随着土壤含水量的降低,绝大多数种源的油松可溶性蛋白质含量呈现出先升高后降低的趋势;而杜金伟[31]研究表明,随着干旱胁迫的加剧,可溶性蛋白质含量呈现下降的趋势。本试验研究表明,不同种源文冠果幼苗叶片可溶性糖质量分数均随着干旱胁迫的加深呈现不同程度的上升趋势;大部分种源文冠果幼苗叶片中可溶性蛋白质质量分数随着干旱胁迫加剧呈上升趋势,只有山西汾阳和河北新华区种源表现出先增加后降低的趋势。当胁迫程度超过了植物渗透调节机制的范围后,渗透调节能力必将会减弱甚至丧失,此刻植物将要启动其他抗旱机制来应对不良环境,由此可以推测,重度干旱胁迫下山西汾阳和河北新华区种源文冠果可溶性蛋白质已无法完成其渗透调节能力,因此出现下降的现象。
大量研究表明,植物的抗旱性是多个因素共同影响所决定的,同一植物的不同种源间也存在着较大的差异,单一的指标并不能体现其抗旱性,因此要用多个指标综合评价。本试验采用模糊数学隶属函数法对6个不同种源的文冠果幼苗进行综合评价,结果表示6个不同种源的文冠果幼苗抗旱性由大到小为内蒙乌丹、新疆察县、山西汾阳、河北新华区、辽宁建平、河南陕州区。
[1] KLICH M G. Leaf variations in elaeagnus angustifolia related to environmental heterogeneity[J]. Environmental & Experimental Botany,2000,44(3):171.
[2] 安玉艳,梁宗锁,郝文芳.杠柳幼苗对不同强度干早胁迫的生长与生理响应[J].生态学报,2011,31(3):716-725.
[3] 刘艳,陈贵林,蔡贵芳,等.干旱胁迫对甘草幼苗生长和渗透调节物质含量的影响[J].西北植物学报,2011,31(11):2259-2264.
[4] KHALIL A M, GRACE J. Acclimation to drought in acer pseudoplatanus L. (Sycamore) seedlings[J]. Journal of Experimental Botany,1992,43(12):1591-1602.
[5] YANG X, CHEN J. Plant litter quality influences the contribution of soil fauna to litter decomposition in humid tropical forests, southwestern China[J]. Soil Biology & Biochemistry,2009,41(5):910-918.
[6] 牟洪香.木本能源植物文冠果(XanthocerassorbifoliaBunge)的调查与研究[D].北京:中国林业科学研究院,2006.
[7] 高述民,马凯,杜希华,等.文冠果(Xanthocerassorbifolia)研究进展[J].植物学通报,2002,19(3):296-301.
[8] 齐鑫淼,李晓宇,丁蕾,等.文冠果研究进展[J].内蒙古林业科技,2013,39(4):37-41.
[9] 牟洪香,于海燕,侯新村.木本能源植物文冠果在我国的分布规律研究[J].安徽农业科学,2008,36(9):3626-3628.
[10] 常月梅,张彩红.文冠果嫁接繁殖技术[J].经济林研究,2013,31(2):154-156.
[11] 莫保儒,王多锋,戚建莉,等.文冠果不同营养器官扦插繁殖试验研究[J].甘肃林业科技,2014,39(1):18-55.
[12] 陈鸥,董凤亮,马海渊,等.文冠果油的萃取及理化性质和脂肪酸组成分析[J].北京农学院学报,2013,28(1):78-80.
[13] 李清河,江泽平.白刺研究[M].北京:中国林业出版社,2011.
[14] 孙群,胡景江.植物生理学研究技术[M].杨凌:西北农林科技大学出版社,2005.
[15] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2002.
[16] 李合生.植物生理生化实验原理与技术[M].北京:高等教育出版社,2000.
[17] 张梅花,张建生,李云霞.干旱胁迫下5种园林地被植物叶片保水力、质膜相对透性和脯氨酸含量的变化[J].甘肃科技,2010,26(10):145-148.
[18] 吴芹,张光灿,裴斌,等.3个树种对不同程度土壤干旱的生理生化响应[J].生态学报,2013,33(12):3648-3656.
[19] 李雪华,蒋德明,阿拉木萨,等.科尔沁沙地4种植物抗旱性的比较研究[J].应用生态学报,2002,13(1):1385-1388.
[20] 姜雪昊,穆立蔷,王晓春,等.3种护坡灌木对干旱胁迫的生理响应[J].草业科学,2013,30(5):678-686.
[21] 武玉叶,李德全.土壤水分胁迫对冬小麦叶片渗透调节及叶绿体超微结构的影响[J].华北农学报,2001,16(2):87-93.
[22] 种培芳,苏世平,李毅,等.不同地理种源红砂幼苗对PEG胁迫的生理响应[J].草业学报,2013,22(1):183-192.
[23] 张智猛,万书波,戴良香,等.花生抗旱性鉴定指标的筛选与评价[J].植物生态学报,2011,35(1):100-109.
[24] 孙存华,李扬,贺鸿雁,等.藜对干旱胁迫的生理生化反应[J].生态学报,2005,25(10):2556-2561.
[25] KOTSIRAS A, OLYMPIONS C M, DROSOPOULOS J, et al. Effects of nitrogen form and concentration on the distribution of ions within cucumber fruits[J]. Scientia Horticulturae,2002,95(3):175-183.
[26] 赵一鹤,李建宾,杨时宇,等.干旱胁迫下3个甜角品种幼苗的生理生化响应及抗旱性评价[J].林业科学研究,2012,25(5):569-575.
[27] 陈少瑜,郎南军,贾利强,等.干早胁迫下坡柳等抗早树种幼苗膜脂过氧化及保护酶活性的影响[J].植物研究,2006,26(1):88-92.
[28] 桑子阳,马履一,陈发菊.干旱胁迫对红花玉兰幼苗生长和生理恃性的影响[J].西北植物学报,2011,31(1):109-115.
[29] 王海珍,梁宗锁,郝文芳,等.白刺花适应土壌干旱的生理学机制[J].干旱地区农业研究,2005,23(1):106-110.
[30] 王琰,陈建文,狄晓艳.水分胁迫下不同油松种源SOD、POD、MDA及可溶性蛋白化较研究[J].生态环境学报,2011,20(10):1449-1453.
[31] 杜金伟.水分胁迫对山杏抗旱生理指标及解剖结构的影响[D].呼和浩特:内蒙古农业大学,2009.
Effect of Drought Stress on the Water Physiological Characteristics and the Osmotic Regulation Substances ofXanthocerassorbifoliaSeedings from Different Provenances//
Zhao Xue
(Agricultural University of Hebei, Baoding 071000, P. R. China);
Zhang Xiuzhen
(Qingdao Guanzhong Ecology Co., Ltd.);
Mu Hongxiang, Sun Xiao, He Hongyue, Yang Kewei, Shan Yuanyuan, Li Chunyou
(Agricultural University of Hebei)//Journal of Northeast Forestry University,2017,45(6):17-21.
Drought stress;Xanthocerassorbifolia; Water physiology characteristics; Osmotic regulation
1)河北省高等学校科学技术项目项目(QN2016153);河北农业大学林学学科创新基金项目(LXXK2014-1)。
赵雪,女,1990年6月生,河北农业大学林学院,硕士研究生。E-mail:zhaoxue_rain@163.com。
牟洪香,河北农业大学林学院,副教授。E-mail:mouhongxiang@hebau.edu.cn。
2017年1月13日。
S722.5
责任编辑:潘 华。
We studied the drought-resistance of six varieties ofXanthocerassorbifoliaBunge. accessions from different provenances, designed three water gradients to study the water physiology and osmotic regulation response to water stress, and then valued it integrative with subordinate function. The seedling leaf water content was decreased and the osmotic regulation substances content was increased with the decrease of soil relative water content. By subordinate function analysis, the drought-resistance of six varieties was in the descending order of Wudan, Neimeng, Chaxian, Xinjiang, Fenyang, Shanxi, Xinhua area, Hebei, Jianping, Liaoning, and Shanzhou area, Henan.
