川西高寒森林溪流好氧不产氧光合细菌多样性
2017-06-28黄春萍吴福忠杨万勤四川农业大学生态林业研究所林业生态工程重点实验室四川成都630四川师范大学生命科学学院四川成都60068
黄春萍,吴福忠,杨万勤,岳 楷,张 川,张 健*(.四川农业大学生态林业研究所,林业生态工程重点实验室,四川 成都 630;2.四川师范大学生命科学学院,四川 成都 60068)
川西高寒森林溪流好氧不产氧光合细菌多样性
黄春萍1,2,吴福忠1,杨万勤1,岳 楷1,张 川1,张 健1*(1.四川农业大学生态林业研究所,林业生态工程重点实验室,四川 成都 611130;2.四川师范大学生命科学学院,四川 成都 610068)
为了探明好氧不产氧光合菌(AAPB)在川西高寒森林溪流极端环境中的生态分布和群落结构及其环境影响因子,本实验结合同步温度动态和水环境特征监测,采用实时荧光定量PCR和克隆文库的方法,检测了川西高寒森林溪流季节性冻融循环3个关键时期(冻结前期、深冻期和融化期)AAPB的丰度和多样性变化及其与环境因子之间的相关性.结果表明:AAPB丰度在冻结前期非常低,仅(2.80±0.19)×104bp/mL;深冻期迅速增加到(4.65±0.30)×104bp/mL;融化期达到(5.79±0.19)×104bp/mL.系统发育分析显示,冻结前期AAPB类群主要为α-变形菌纲细菌;而深冻期和融化期以β-变形菌纲细菌占据主要地位.相关性分析表明,在季节性冻融循环过程中,叶绿素a、可溶性有机碳和温度是AAPB菌群丰度和多样性的主要影响因子.实验结果表明冬季的川西高寒森林溪流中,AAPB类群的丰度低,但是它们高度动态变化,并且与其他淡水环境中的该类群的同源性低.
好氧不产氧光合菌;丰度和多样性;高寒森林溪流;季节性冻融循环
好氧不产氧光合菌(AAPB)是一类含有细菌叶绿素 a,且能同时进行光合代谢和有机物质代谢吸收能量的原核生物[1-2].海洋中的AAPB已经被深入研究,这类原核生物在自然环境中占据着广泛的生态位[2-5].然而,淡水环境中的这一类群被关注的相对较少[6-7].另一方面,AAPB的丰度只维持极小的比例,并在不同的环境中差异极大[8-9]. AAPB具有灵活的代谢活性, 并广泛分布在水体透光层,AAPB在水生食物链和整个生物地球化学循环过程中具有十分重要的作用[10].已有研究表明,绝大多数的AAPB都归属于α-、β-和 γ-变形菌纲,并且在不同的环境中,α-或者 γ-变形菌纲占据群落优势地位,群落结构组成也随着不同的生态环境而发生变化[11].但是,在极端的高寒森林溪流环境中相关研究比较匮乏.
光、可溶性有机碳(DOC)、盐、硝态氮和氨等,都可能影响水体中AAPB的分布、丰度和群落组成[10-11].在河口和海洋中的研究结果表明AAPB与叶绿素a的含量呈显著正相关性[12].同时,基于pufM基因对AAPB季节性变化的研究结果表明,AAPB会受到环境因子、温度、DOC、叶绿素 a以及光照的深刻影响从而表现出高度的季节性动态特征[13].但是,在温带的高寒森林中,关于环境因子对AAPB类群的组成结构及多样性的调控研究非常缺乏,致使对AAPB这一类群在高寒森林溪流环境中的分布、丰度、活性及其季节性动态的理解也变得薄弱.
川西高山/亚高山森林地处青藏高原东缘和长江上游地区,是长江流域的水源涵养地,为全球气候变化最敏感的区域之一[14],她也是长江流域最重要的“水塔”,在水源涵养、指示全球气候变化、生物多样性保育等方面具有十分重要且不可替代的作用和地位,其结构和功能直接关系到岷江流域乃至整个长江上游的生态安全[15].课题组在研究区域海拔3600~3700m典型高寒森林范围内,沿主河道两岸一共找到 18条森林溪流汇入其中[16],这些不同性质的溪流将森林、河岸和河流(湖泊)连接成为一个巨大的水源涵养体.这些溪流作为开放的生态系统并且广泛分布,鲜有人为干扰,低盐寡营养、低温以及具有高寒森林典型的特点,包括从冬季雪被的完全覆盖到夏季高辐射强度的季节性变化.本文结合同步温度动态和水环境特征监测,采用实时荧光定量PCR和克隆文库的方法检测了川西高寒森林溪流冬季冻融循环3个关键时期(冻结前期、深冻期和融化期)AAPB的丰度、结构组成和多样性及其季节性变化,探究影响AAPB群落结构季节性变化的主要环境因子,以期为深入研究全球气候变暖下,高寒森林溪流环境对微生物群落的影响,理解气候变暖情景下高寒森林生态系统的区域响应提供一定科学理论依据.
1 材料和方法
1.1 研究区域概况
研究区域位于四川省理县毕棚沟高山森林生 态 系统 定 位站(31°14′~31°19′N,102°53′~102°57′E,海拔 2458~4619m),地处青藏高原东缘与四川盆地的过渡带.区域气候属丹巴-松潘半湿润气候,随着海拔上升,气候呈现暖温带、温带、寒温带、亚寒带和冰冻带的垂直分布规律.该区域年降水量随着海拔高度的变化而变化,大约为801~850mm,降雨主要分布在 6~9月,年均气温2~4℃,最高气温(7月)23℃,最低气温(1月)-18℃.研究区域土壤季节性冻结期为每年 11月初~次年4月中旬,且季节性冻融期长达5~6个月,冻融作用明显.区域集水区系岷江上游的杂谷脑河第二大支流梭罗沟的支流,集水区面积 179km2,河长31km,年平均流量3.7m3/s.高寒森林由四川红杉和方枝柏构成冠层优势物种;康定柳和高山杜鹃构成了高寒森林河岸优势植物[14,16-17].
1.2 样品采集
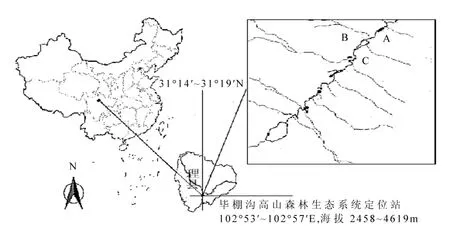
图1 研究样地溪流采样位点(A, B和C)Fig.1 A pattern diagram showing the experimental streams (A, B and C) in the study site
基于前期调查的结果[26],本实验在研究区域内沿主河道海拔 3582m 的位置选择3条典型的高寒森林溪流作为重复采样位点(图 1).分别于2013年12月23日(冻结前期)、2014年3月10日(深冻期)和2014年4月23日(融化期)在各采样点用灭菌的安剖瓶采集表层10~20cm的溪流水,低温保存带回实验室.在每次采样时即时测定采样点水样pH值和流速,并且在各采样点放置1个纽扣式温度计,同步记录采样点水温每两小时变化情况(图2).
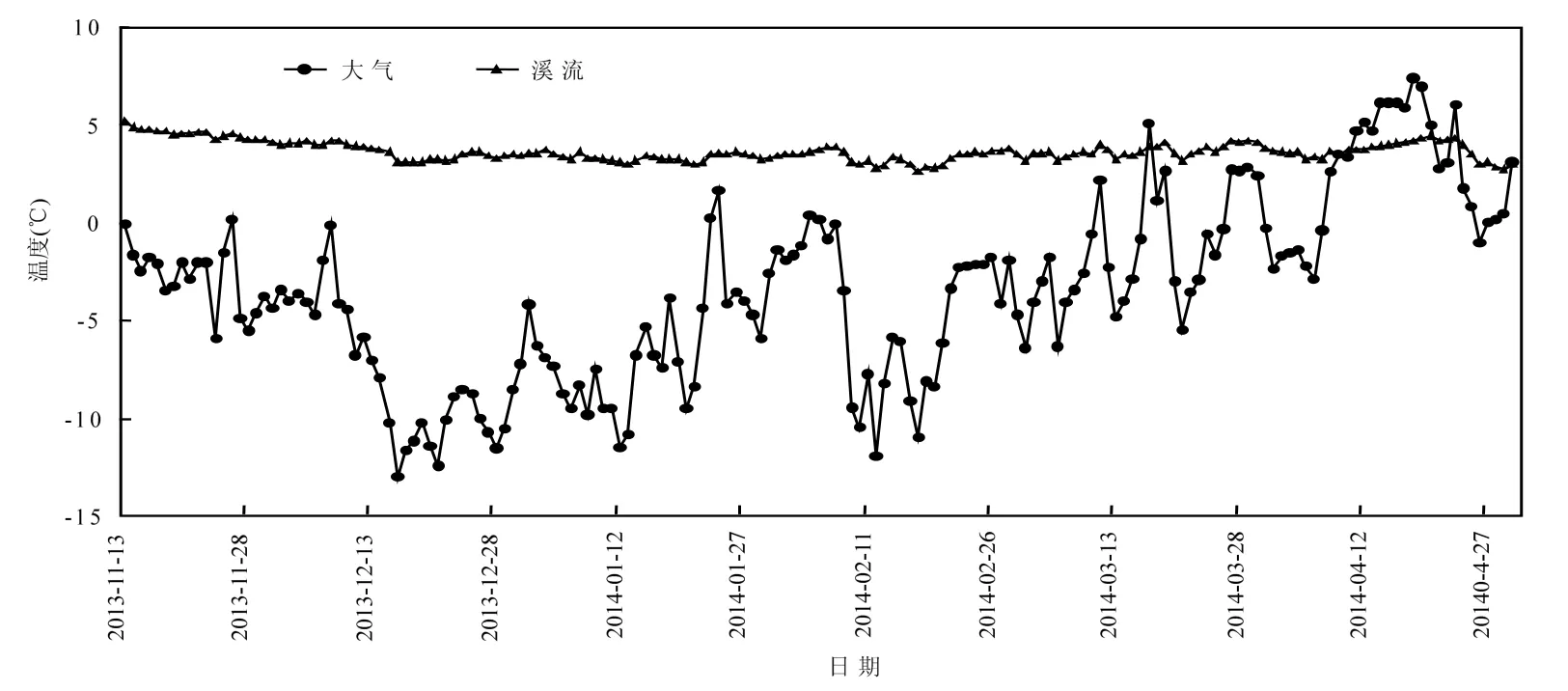
图2 2013-11-13~2014-04-30高寒森林溪流和大气温度动态Fig.2 Temperature dynamics of the stream and the atmosphere in the alpine fir forest from Nom 13, 2013 to Aug 30, 2014)
1.3 样品分析
将各水样500mL用0.22µm的微孔滤膜过滤,滤液采用双指示剂中和法测定碳酸氢根(HCO32)含量,紫外分光光度法测定磷酸根(PO43)含量,离子色谱测定铵态氮(NH4+N)和硝态氮(NO3N),multi N/C 2100, analytikjena测定水溶性有机碳(DOC)等[18],结果见表1.
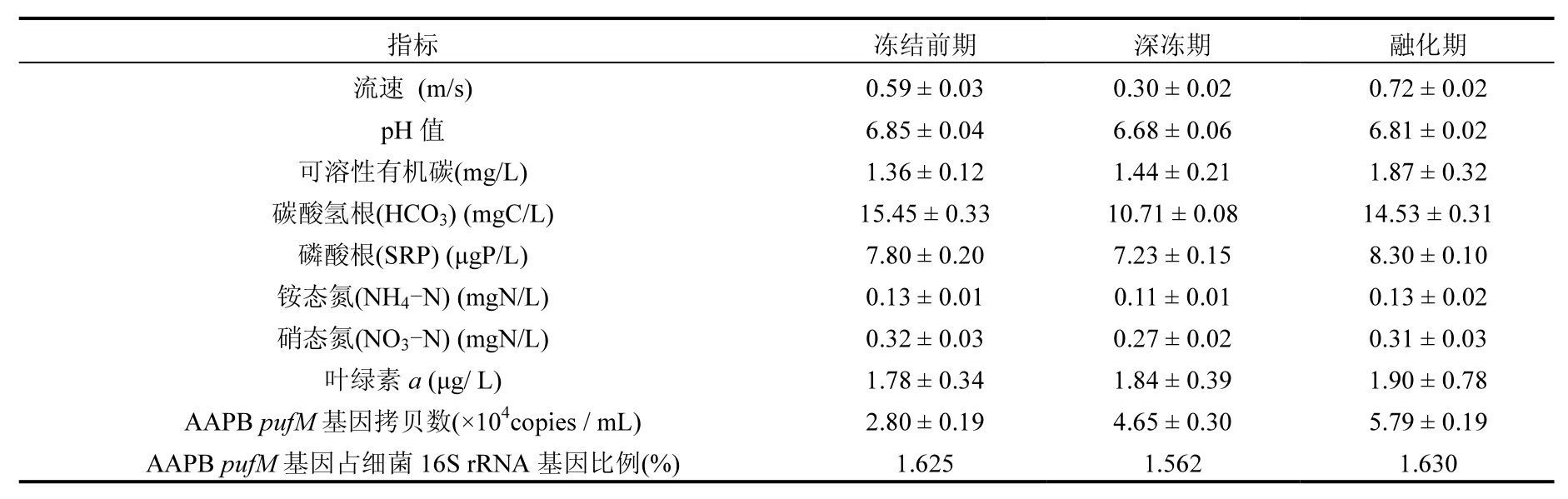
表1 采样期溪流水特征、AAPB pufM基因丰度及pufM基因所占细菌16S rRNA基因的比例Table 1 Stream water physicochemical characteristics, AAPB puf M gene numbers and % pufM gene of bacterial 16S rRNA gene in the researched area during each sampling time
滤膜采用-80℃冷冻干燥[19],一部分按照文献[20-21]的方法,采用高效液相色谱法检测叶绿素a和细菌叶绿素 a的含量.为了保证色素不被光降解,样品均在暗光条件下处理[21],在采样期内均未检测到细菌叶绿素a,结果见表1.
另一部分滤膜参照文献[22]的方法对总DNA进行提取与纯化.AAPB pufM 基因引物为pufM-557F (5-TACGGSAACCTGTWCTAC-3)/ pufM-750R (5-CCATSGTCCAGCGCCAGAA-3),细菌 16S rRNA 基因引物为 338f (5-CCTACGGGAGGCAGCAG-3)/518r(5-ATTACCGCGGCTGCTGG-3)[2,6,23].采用经过琼脂糖凝胶电泳回收纯化后的总DNA 10~100ng作模板,进行PCR扩增.扩增得到的pufM基因产物纯化后与pUC19载体进行连接,连接产物转化感受态细胞DH5α,在含有Ampicillin、IPTG和X-Gal的LB琼脂平板培养基上37℃培养24h,挑选阳性克隆.共创建 3个 pufM 基因克隆文库(S-FP、S-FPD和S-TP).用M13-47和RV-M引物对进行菌落PCR扩增后,选取阳性克隆并提取质粒供qPCR反应的标准曲线使用.质粒浓度采用ScanDrop100超微量核酸蛋白测定仪测定,然后分别进行10倍梯度系列稀释以作为定量PCR扩增的标准品进行扩增,根据所得标准曲线计算得出样品中的基因拷贝数,最后以每克样品(干重)中的基因拷贝数为单位进行分析.PCR反应效率和标准曲线的相关系数分别为:细菌,107.5%和r2= 0.998; AAPB,103.3%和r2= 0.995.
1.4 数据处理与分析
测序结果采用DNAstar和Cluster软件进行序列分析,下载最相似的菌株序列作为系统发育树的参考序列.采用 MEGA软件,Neighborjoining法构建系统发育树,自展数为 1000.克隆文库的 Shannon-Weaver等指数采用 PAST program基于OTU水平计算.基因丰度和多样性指数采用SPSS 19.0统计软件进行单因素方差分析(ANOVA),多重比较采用最小显著差异法(LSD),AAPB多样性和基因丰度与环境因子的相关关系采用Pearson法分析,AAPB群落与理化因子的冗余分析(RDA)采用Canoco for Windows 4.5软件.数据整理、计算与作图均采用Microsoft Excel 2013软件.测序所得的基因序列在GenBank中的登录号为KT279202 ~KT279344.
2 结果与分析
2.1 AAPB pufM基因丰度
荧光定量PCR结果显示,冬季不同冻融时期的溪流水中均检测到 AAPB pufM基因.从冻结前期到融化期,AAPB pufM基因丰度呈显著增加趋势,冻结前期、深冻期和融化期每毫升溪流水中的拷贝数分别为(2.80±0.19)×104、(4.65±0.30)× 104和(5.79±0.19)×104;各个时期AAPB pufM基因所占细菌16S rRNA基因的比例则表现出先降低再升高的趋势,3个时期分别为 1.625%、1.562%和1.630%(表1).
2.2 AAPB pufM 基因多样性
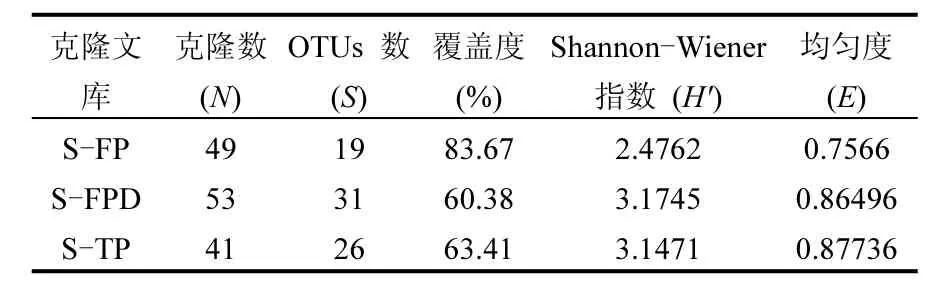
表2 不同克隆文库pufM 基因多样性Table 2 Diversity of pufM gene in different clone libraries
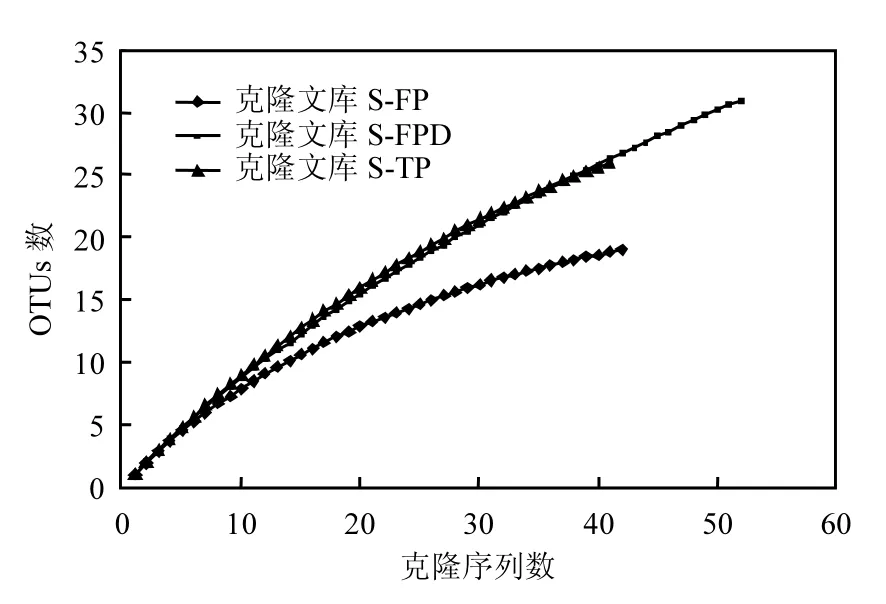
图3 AAPB pufM基因克隆文库稀释曲线Fig.3 Rarefaction curve analysis of the AAPB pufM genes
采用克隆文库的方法检测了 3个不同时期的AAPB群落结构,构建了3个克隆文库(S-FP、S-FPD和S-TP).总共获得了143 条pufM 基因序列用于系统发育分析,其中S-FP 49条,S-FPD 53条,S-TP 41条.这些条带在各自库中的覆盖度分别为 S-FP 83.67%,S-FPD 60.38%及 S-TP 63.41%(表2).稀释曲线(图3)显示3个克隆文库并未饱和,说明在 3个关键时期依然存在更丰度的AAPB物种.基于Zeng等的研究,pufM基因序列以cutoff = 0.06和0.15分别在种和属水平进行同源性和系统发育分析[24],本实验将得到的 143条序列,以cutoff = 0.06分为78个OTUs,主要归属于 α-变形菌纲(18 OTUs,占所有序列的23.68%)和 β-变形菌纲(19 OTUs,占所有序列的24.36%)(图 4).α-变形菌纲主要包括红假单胞菌属(Rhodopseudomonas sp.)、 甲基杆菌 属(Methylobacterium sp.)和 玫 瑰 杆 菌 属(Roseobacter sp.)的细菌,而 β-变形菌纲主要包括红长命菌属(Rubrivivax sp.)、纤毛菌属(Leptothrix sp.)和3个其他类群的细菌.另外,还有7个OTUs属于γ-变形菌纲.通过分析3个克隆文库,从冻结前期到融化期,占优势地位的 AAPB类群发生一定的改变,冻结前期以 α-变形菌纲为主要类群,而β-变形菌纲在深冻期和融化期占据优势地位.

图4 AAPB pufM基因序列系统发育分析Fig.4 Phylogenetic relationship (neighbor-joining tree ) of pufM gene clone sequences
2.3 AAPB群落结构与环境因子的相关性
为了解环境因子对AAPB的影响,对AAPB丰度及其群落结构与环境因子之间的相关性进行了分析,结果见表3.AAPB pufM基因丰度分别与细菌16S rRNA基因丰度,叶绿素a和温度呈极显著的正相关性(P<0.01),且与DOC呈显著正相关性(P<0.05),将环境因子与 AAPB群落结构进行冗余分析(RDA)(图 5),结果显示叶绿素 a、温度和DOC与α-变形菌纲细菌、β-变形菌纲细菌及一些未知细菌类群关系密切,呈正相关性;而叶绿素a、温度和DOC均与γ-变形菌纲细菌呈负相关性.γ-变形菌纲细菌受到水流流速、铵态氮含量、硝态氮含量、碳酸氢根含量、磷酸根含量及 pH值的深刻影响,均呈正相关性.同时这些因子均与 α-变形菌纲细菌、β-变形菌纲细菌及未知的细菌类群呈负相关性.另外,所检测的环境因子与AAPB的Shannon-Wiener多样性指数呈正相关性.
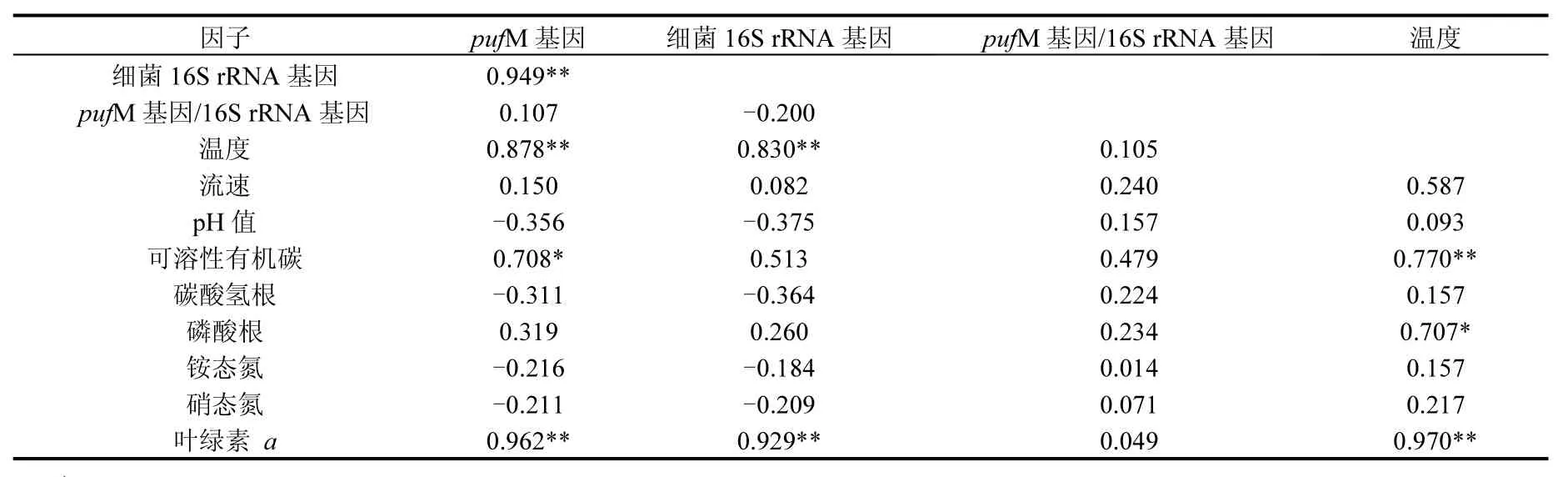
表3 AAPB pufM基因丰度、细菌16S rRNA基因丰度与环境因子之间的相关性分析Table 3 Pearson’s correlation analyses among the abundance of bacteria 16S rRNA gene and AAPB pufM gene, and environmental variables

图5 环境因子与AAPB群落结构的RDA排序Fig.5 RDA ordination diagrams showing correspondence between AAPB community structures and environmental variables
3 讨论
AAPB群落结构及其动态对微生物生态学过程十分重要,海洋和河口环境中AAPB类群的系统发育,结构组成及其与环境之间的关系都有深入的研究[12].但是,温带高寒森林溪流生境中这类菌群的研究相对较少[13].因此,本研究对冬季冻融时期高寒森林溪流 AAPB的丰度和结构动态的深入理解具有十分重要的生态学意义.
冬季土壤冻融循环的3个关键时期,川西高寒森林溪流中均检测到 AAPB,其丰度较低并呈动态变化,丰度值在已有研究的范围内[13].冻融循环是高寒森林生态系统关键的季节性转变时期[14,17],其冻融特征的差异使得微生物结构存在显著的变化.AAPB丰度从冻结前期到融化期呈增加趋势,而其所占细菌丰度的比例却先降低后增长,这些结果反映了AAPB这一类群与其他微生物类群不同的季节性模式和作用,以及与其他生理类群的细菌不同的环境响应.这可能是因为AAPB个体和基因组都较普通细菌个体大,这使得它们需要更多的生存能量[25],因此在寡营养的高寒森林溪流中,相对于普通细菌,尽管AAPB能通过光合作用获得能量,却没有竞争优势.
已有研究认为光照、营养物质、温度和叶绿素a可能是调控AAPB丰度的主要因子[10,13].本实验结果表明温度、叶绿素a和DOC是川西高寒森林溪流中AAPB时空分布的主要影响因子,它们强烈影响AAPB的丰度和结构,同时可溶性磷和流速也有一定程度的贡献.与其它生境中的研究结果不同,川西高寒森林溪流中AAPB丰度与pH值、碳酸氢根、铵态氮和硝态氮呈负相关性.同时,AAPB丰度与温度的相关性极强,这也与其它高山湖泊中的较弱相关性结果截然不同[5].自从被发现以来,AAPB的光合代谢方式在寡营养,尤其是低碳的环境中具有重要的竞争优势[8],大部分的研究认为 AAPB在环境较好的条件下生长更好[3,6],但也有研究得到相反结果[12-13].本研究结果显示川西高寒森林溪流中 AAPB丰度随着叶绿素a和DOC的增加而增加,均呈极显著正相关性(P<0.01),这与寡营养高山湖泊和海洋中DOC是调控AAPB丰度的主要因子之一的研究结果一致[26].同时可以推测,在寡营养的高寒森林溪流中,光合作用相关基因的表达并不活跃,但是营养物质浓度参与决定了pufM基因携带者的存在和丰度.比如,本研究中AAPB的丰度比富营养化的太湖中AAPB的丰度低3个数量级[27].
本研究中 AAPB的群落结构多样性也与淡水环境中的相关研究结果有差异[5].稀释曲线显示所构建的克隆文库并不完全饱和,而且各个时期 OTUs的精确数量可能被低估,但是实验结果依然可以表明各个时期pufM基因的高度多样性.同时 Shannon-Weaver指数(2.48-3.17)也表现出较其它环境中高的特点,甚至比乌梁素海Shannon-Weaver指数还高[28].系统发育分析结果显示AAPB丰富的遗传结构组成除了α-变形菌纲的红假单胞菌属、甲基杆菌属、玫瑰杆菌属,β-变形菌纲的红长命菌属和纤毛菌属及 γ-变形菌纲的一些类群外,还有一些与 Arctic spring bacterium sp.和Roseateles sp.同源性较高的类群.甚至还有许多未知的AAPB类群,它们在系统发育分析中组成另一支,并未与参考序列混淆,这表明了川西高寒森林溪流中可能蕴藏着携带pufM基因的新的微生物资源.已有大量研究表明β-变形菌纲是淡水生境中的优势 AAPB类群[7],但是本研究发现,β-变形菌纲并不总是在川西高寒森林溪流环境中占据优势地位,α-变形菌纲的优势地位在冬季冻结前期较β-变形菌纲更突出,只是随着时间的推移以及环境因子的变化,这种优势地位开始下降.另一方面,与海洋环境联系紧密的γ-变形菌纲在川西高寒森林溪流中也被检测到.但是,被认为在高山湖泊和河流中广泛分布的红育菌属(Rhodoferax sp.)和鞘氨醇单胞菌属(Sphingomonas sp.),却没有被检测到.这可能与温度有关,也可能是由于溪流环境冬季紫外辐射较低的原因[7,9].
4 结论
4.1 川西高寒森林冬季溪流中AAPB丰度较低,在冻融循环冻结前期仅(2.80±0.19)×104bp/mL;深冻期迅速增加到(4.65±0.30)×104bp/mL;融化期达到(5.79±0.19)×104bp/mL.同时,AAPB 群落结构多样性丰富且高度动态变化,冻融循环冻结前期α-变形菌纲细菌为 AAPB 优势类群,而深冻期和融化期则由β-变形菌纲细菌占据优势地位.
4.2 环境因子共同调控着AAPB的丰度、多样性及其群落结构.其中,叶绿素 a、可溶性有机碳和温度是主要的调控因子,并正调控α-变形菌纲和β-变形菌纲,负调控γ-变形菌纲.
[1]Boeuf D, Humily F, Jeanthon C. Diversity of Arctic Pelagic Prokaryotes with an emphasis on photoheterotrophic bacteria: a review [J]. Biogeosciences Discussions, 2014,11(2):2419—455.
[2]Béjà O, Suzuki M T, Heldelberg J F, et al. Unsuspected diversity among marine aerobic anoxygenic phototrophs [J]. Nature, 2002, 415:630—633.
[3]Kolber Z S, Plumley F G, Lang A S, et al. Contribution of aerobic photoheterotrophic bacteria to the carbon cycle in the ocean [J]. Science, 2001,292:2492—2495.
[4]Kalashnikov A M, Gaisin V A, Sukhacheva M V. Anoxygenic phototrophic bacteria from microbial communities of Goryachinsk thermal spring (Baikal Area, Russia) [J]. Microbiology, 2014,83(4):407—421.
[5]Mašín M, Nedoma J, Pechar L, et al. Distribution of aerobic anoxygenic phototrophs in temperate freshwater systems [J]. Environmental Microbiology, 2008,10:1988—1996.
[6]Joan C, Emilio O C. Environmental controls and composition ofanoxygenic photoheterotrophs in ultraoligotrophic high-altitude lakes (Central Pyrenees) [J]. Environmental Microbiology Reports, 2004,6(2):145—151.
[7]Salka I, Cuperova Z, Mašín M, et al. Rhodoferax-related pufM gene cluster dominates the aerobic anoxygenic phototrophic communities in German freshwater lakes [J]. Environmental Microbiology, 2011,13:2865—2875.
[8]Kolber Z S, Van Dover C L, Niederman R A, et al. Bacterial photosynthesis in surface waters of the open ocean [J]. Nature, 2000,407:177—179.
[9]Waidner L A, Kirchman D L. Aerobic anoxygenic photosynthesis genes and operons in uncultured bacteria in the Delaware River [J]. Environmental Microbiology, 2005,7:1896—1908.
[10]Monica R S, Matthew T C, David L K. Leucine incorporation by aerobic anoxygenic phototrophic bacteria in the Delaware estuary [J]. International Society for Microbial Ecology, 2014,8:2339—2348.
[11]Jeanthon C, Boeuf D, Dahan O, et al. Diversity of cultivated and metabolically active aerobic anoxygenic phototrophic bacteria along an oligotrophic gradient in the Mediterranean Sea [J]. Biogeoscience, 2011,8:1955—1970.
[12]Lehours A C, Cottrell M T, Dahan O, et al. Summer distributionand diversity of aerobic anoxygenic phototrophic bacteria in the Mediterranean Sea in relation to environmental variables [J]. FEMS Microbiology Letters, 2010,74:397—409.
[13]Ferrera I, Borrego C M, Salazar G, et al. Marked seasonality of aerobic anoxygenic phototrophic bacteria in the coastal NW Mediterranean Sea as revealed by cell abundance, pigment concentration and pyrosequencing of pufM gene [J]. Environmental Microbiology, 2014,16(9):2953—2965.
[14]岳 楷,杨万勤,彭 艳,等.高寒森林溪流对凋落叶分解过程中木质素降解的影响 [J]. 植物生态学报, 2016,40(9):893—901.
[15]何奕忻,吴 宁,朱求安,等.青藏高原东北部5000年来气候变化与若尔盖湿地历史生态学研究进展 [J]. 生态学报, 2014,34(7):1615—1625.
[16]李 俊,吴福忠,杨万勤,等.长江上游高山森林小溪流磷输出及其汇流特征 [J]. 环境科学学报, 2015,35(4):1136—1142.
[17]He W, Wu F Z, Zhang D J, et al. The effects of forest gaps on cellulose degradation in the foliar litter of two shrub species in an alpine fir forest [J]. Plant and Soil, 2015,393(1):109—122.
[18]Chon K, Sarp S, Lee S, et al. Evaluation of a membrane bioreactor and nanofiltration for municipal waste water reclamation: Trace contaminant control and fouling mitigation [J]. Desalination, 2011,272(1):128—134.
[19]白 洁,刘小沙,侯 瑞,等.南海南部海域浮游细菌群落特征及影响因素研究 [J]. 中国环境科学, 2014,34(11):2950—2957.
[20]Mantoura R F C, Llewellyn C A. The rapid determination of algal chlorophyll and carotenoid pigments and their breakdown products in natural waters by reverse-phase high performance liquid chromatography [J]. Analytica Chimica Acta, 1983,151:297—314.
[21]Ruivo M, Cartaxana P, Cardoso M I, et al. Extraction and quantification of pigments in aerobic anoxygenic phototrophic bacteria [J]. Limnology and Oceanography: Methods, 2014,12:338—350.
[22]Wang A, Wu F Z, Yang W Q, et al. Abundance and composition dynamics of soil ammonia-oxidizing archaea in an alpine fir forest on the eastern Tibetan Plateau of China [J]. Canadian Journal of Microbiology, 2012,58:572—580.
[23]Bakkea I, Schryverb P De, Boonb N, et al. PCR-based community structure studies of Bacteria associated with eukaryotic organisms: A simple PCR strategy to avoid co-amplification of eukaryotic DNA [J]. Journal of Microbiological Methods, 2011,84(2):349—351.
[24]Zeng Y H, Chen X H, Jiao N Z. Genetic diversity assessment of anoxygenic photosynthetic bacteria by distance based grouping analysis of pufM sequences [J]. Letters in Applied Microbiology, 2007,45(6):639−645.
[25]Sieracki M E, Gilg I C, Thier E C, et al. Distribution of planktonic aerobic anoxygenic photoheterotrophic bacteria in the northwest Atlantic [J]. Limnology and Oceanography, 2006,51(1):38—46.
[26]张 瑶.海洋典型功能细菌群的生态过程研究 [D]. 厦门:厦门大学, 2006.
[27]Shi L M, Cai Y F, Chen Z T, et al. Diversity and abundance of aerobic anoxygenic phototrophic bacteria in two cyanobacterial bloom-forming lakes in China [J]. Annales De Limnologie-International Journal of Limnology, 2010,46:233—239.
[28]何一平.基于 pufM 基因的乌梁素海好氧不产氧光合细菌系统发育多样性分析 [D]. 呼和浩特:内蒙古大学, 2010.
The diversity of aerobic anoxygenic phototrophic bacteria in a high-frigid forest stream of West Sichuan
HUANGChun-ping1,2, WU Fu-zhong1, YANG Wan-qin1, YUE Kai1, ZHANG Chuan1, ZHANG Jian1*(1.Key laboratory of Ecological Forestry Engineering of Sichuan Province, Institute of Ecology & Forest, Sichuan Agricultural University, Chengdu 611130, China;2.College of life science, Sichuan Normal University, Chengdu 610101, China). China Environmental Science, 2017,37(6):2332~2339
In order to understand AAPB community structure and distribution in a high-frigid forest stream with cold environment, here, a field investigation was carried out in West Sichuan, China. Water was sampled in the stream at onset of the freezing period (FP), the deep freezing period (FPD) and the thawing period (TP). The methods of real-time fluorescent quantitative PCR and clone library were used to check the richness and diversity of AAPB. Temperature and other water characters were also observed to analysis the relationships between environment and AAPB. The results showed that low abundance (2.80 ± 0.19) ×104bp / mL of AAPB was observed, at FP, but which rapidly increased to (4.65 ± 0.30) ×104bp / mL at FPD, then reached to (5.79 ± 0.19) ×104bp / mL at TP. Phylogenetic relationships from clone libraries displayed that Alphaproteobacteria classes dominated AAPB at FP, but Betaproteobacteria classes dominated AAPB at both FPD and TP. In addition, correlationships revealed that the concentration of chlorophyll a and dissolved organic carbon, as well as average temperature had closely relationships with AAPB abundance and diversity during the seasonal freeze-thaw period. All the results suggest that AAPB have low abundance but exhibit highly dynamical patters in the high-frigid forest streams and their communities share low homology with those in other freshwater.
aerobic anoxygenic phototrophic bacteria (AAPB);abundance and diversity;high-frigid forest stream;seasonal freeze-thaw cycle
X172
A
1000-6923(2017)06-2332-08
黄春萍(1980-),女,四川乐山人,副教授,博士,主要从事微生物生态学和环境微生物学方向研究.发表论文20余篇.
2016-11-10
国家自然科学基金资助项目(31500358,31570445, 31570601)
* 责任作者, 教授, sicauzhangjian@163.com
