TaCYP78A5基因过表达小麦的遗传和功能初步分析
2017-06-27陈之忍申玉霞吴林楠刘香利赵惠贤
陈之忍,马 猛,申玉霞,吴林楠,刘香利,2,赵惠贤,2
(1.西北农林科技大学生命科学学院,陕西杨陵 712100;2.西北农林科技大学旱区作物逆境生物学国家重点实验室,陕西杨陵 712100)
TaCYP78A5基因过表达小麦的遗传和功能初步分析
陈之忍1,马 猛1,申玉霞1,吴林楠1,刘香利1,2,赵惠贤1,2
(1.西北农林科技大学生命科学学院,陕西杨陵 712100;2.西北农林科技大学旱区作物逆境生物学国家重点实验室,陕西杨陵 712100)
为了验证小麦籽粒大小相关基因 TaCYP78A5在小麦籽粒发育中的功能,对pINO启动子驱动的 TaCYP78A5基因过表达的转基因小麦后代株系进行了鉴定,检测了T0代植株目标基因拷贝数,定量分析了7个T1代阳性植株的目标基因表达,并对其籽粒大小进行了统计。结果表明,利用Bar试纸条和目标基因特异PCR检测相结合的方法对21株转基因T0代再生苗进行检测,共鉴定出14个阳性植株,除2个植株的目标基因拷贝数为3和1个植株为7外,其余11个T0代转基因植株目标基因插入拷贝数均为1~2个,其中有6个单拷贝植株。与野生型相比,7个T1代阳性植株目标基因表达量均极显著增加,粒厚和粒宽均有不同程度增加,粒重极显著增加。
小麦; TaCYP78A5基因;Bar基因;转基因株系
小麦(TriticumaestivumL.)是主要的粮食作物之一,在谷物种植中居于首位[1]。随着社会经济的发展,全球范围内耕地面积下降,环境逐渐恶化,使得小麦生产面临巨大挑战。因此,提高小麦产量,维持其可持续供应具有重要意义[2]。籽粒大小是决定谷物产量的重要组成因素之一,也是现代作物育种的一个重要选择性状[3-4]。在水稻和拟南芥中,许多与种子大小相关的基因都已被克隆并进行了功能鉴定,但小麦种子大小相关基因的分离和功能研究仅有个别报道[5]。
细胞色素P450(Cytochromes P450)是植物最大的蛋白家族之一[6],此类蛋白含有硫羟基和血红素结构域,在多个生化代谢途径中起着重要的作用,参与多种生物合成和分子解毒[7]。 CYP78A家族是植物特有的细胞色素P450家族成员,在陆生植物中高度保守[8]。许多 CYP78A家族成员作用于植物生殖器官和籽粒的生长发育,被认为是可以用于作物改良[9]。例如,在拟南芥中, KLUH(KLU)/CYP78A5以及 CYP78A7通过非细胞自主信号来促进器官的生长,它们的活性高低与种子大小成正相关[10];在水稻中, CYP78A13通过调节胚与胚乳之间的大小平衡来调节种子的大小,功能缺失突变体的种子胚增大但胚乳减小,而过表达则会导致种子的胚减小和胚乳增大[4,11];在番茄中,KLUH的直系同源基因SIKLUH通过增加果皮和中隔组织细胞数目增大了果实的体积,提高了果实的品质[12]。
本实验室马 猛等[5,9]采用大麦条斑花叶病毒诱导基因沉默(BSMV-VIGS)的技术研究发现,小麦 TaCYP78A3和 TaCYP78A5基因沉默可导致小麦籽粒变小,而在拟南芥中过表达 TaCYP78A3和 TaCYP78A5基因可导致其种子增大。由此初步认为, TaCYP78A3和 TaCYP78A5基因具有调控小麦籽粒大小的功能。为了进一步验证 TaCYP78A5基因在小麦种子发育中的功能,本研究以拟南芥胚珠珠被特异表达启动子pINO(Promoter of INNER NO OUTER)驱动的 TaCYP78A5基因过表达载体pCAMBIA3301-TaCYP78A5 转化春小麦JW1所得T0代和T1代转基因植株为材料,进行分子检测及遗传 分析。
1 材料与方法
1.1 材 料
1.1.1 植物材料及载体
由农杆菌转化并经PPT筛选携带 TaCYP78A5基因的T0代转基因植株和野生型受体小麦JW1由济南邦迪生物科技有限公司提供。转化所用双元植物表达载体pCAMBIA3301- TaCYP78A5由本实验室程绘绘[13]构建,其T-DNA结构如图1所示。
1.1.2 试剂
2×EsTaqMasterMix和DNA Marker DL2000为康为世纪产品;2× Premix ExTaq、Fruit-mateTMfor RNA Purification和RNAiso Plus为TaKaRa产品;GoScriptTMReverse Transcriptase System和2× SYBR Green Master Mix为Promega产品;PAT/Bar检测试纸条为EnviroLogix产品;引物和探针由英潍捷基(上海)贸易有限公司合成。
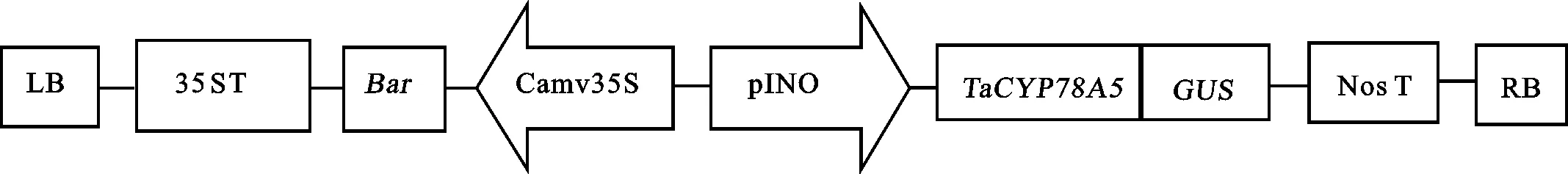
Camv35S:花椰菜病毒35S强启动子;pINO:胚珠珠被特异性启动子。
Camv35S:Cauliflower mosaic virus 35S promoter; pINO:Promoter of INNER NO OUTER.
图1 植物表达载体pCAMBIA3301- TaCYP78A5的T-DNA结构
Fig.1 T-DNA region of plant expression vector pCAMBIA3301- TaCYP78A5
1.2 方 法
1.2.1 T0代转基因植株Bar试纸条检测及目标基因特异PCR检测
野生型JW1和农杆菌转化筛选后再生植株于株高3~5 cm时进行移栽,移栽前剪取部分叶片至1.5 mL离心管中,用研磨棒捣碎,加入500 μL EB2 Extraction Buffer,将试纸条按正确方向插入混合液中,静置10 min,观察条带的变化。
再生植株移栽温室后,三叶期取叶片,利用CTAB法提取其基因组DNA。以ddH2O为空白对照,野生型小麦JW1为阴性对照,植物表达载体pCAMBIA3301- TaCYP78A5为阳性对照,利用特异性引物SPA5-F/SPA5-R(SPA5-F:5′-GC TACGACGATTTCACAAGCCAAG-3′,位于目标基因 TaCYP78A5内;SPA5-R:5′-GCCTG CCCAACCTTTCGGTAT-3′,位于GUS基因内)对转基因小麦进行PCR鉴定。PCR反应体系:2× EsTaqMasterMix 10 μL,10 μmol·L-1的上下游引物各1 μL,DNA模板1 μL,ddH2O 8 μL。反应条件:95 ℃预变性5 min;95 ℃变性30 s,62 ℃退火30 s,72 ℃延伸45 s,35个循环;72 ℃延伸10 min。PCR产物经1%琼脂糖凝胶电泳检测后,照相分析。
1.2.2 T0代转基因植株拷贝数检测
通过qPCR(TaqMan探针法)对Bar试纸条及目标基因特异PCR鉴定均为阳性的转基因植株进行拷贝数检测。内参基因选用小麦中的单拷贝基因Pinb[14]。使用Primer Premier 5.0设计目标基因所用引物和探针,为了防止内源基因对检测的干扰,上游引物位于目标基因 TaCYP78A5上,下游引物和探针位于GUS基因上。引物和探针序列见表1。以5倍梯度稀释的转基因植株T0A5-21的基因组DNA为模板,ddH2O为空白对照,野生型小麦JW1为阴性对照,进行qPCR,3个重复,得到Pinb和 TaCYP78A5基因的标准曲线。PCR反应体系:2× Premix ExTaq10 μL,10 μmol·L-1的上下游引物各0.25 μL,5 μmol·L-1的TaqMan探针1.5 μL,DNA模板1 μL,ddH2O 7 μL。PCR反应条件:95 ℃预变性30 s;95 ℃变性5 s,60 ℃延伸20 s,40个循环。参照刘振华等[14]和Weng等[15]的方法对 TaCYP78A5基因的拷贝数进行测定,各待测样品设置3个重复得到其Ct值,由公式X=2×10[(Ctx-Ix)/Sx-(CtR-IR)/SR] 计算出目标基因的拷贝数。其中,IX和IR是 TaCYP78A5和Pinb的标准曲线的截距,SX和SR是 TaCYP78A5和Pinb的标准曲线的斜率,CtX和CtR是各待测样品 TaCYP78A5与Pinb基因的Ct值。 TaCYP78A5基因的拷贝数估测值是将X值的小数按大于0则入的原则只保留整数得到的。

表1 实时荧光定量PCR的引物及探针序列Table 1 Sequences of primer and probe for real-time PCR
1.2.3 T1代单拷贝转基因植株最适Basta浓度的确定及其目标基因的遗传分析
将6个T0代单拷贝植株及野生型JW1植株种子在温室中加代繁殖,于三叶期,利用CTAB法提取T1代基因组DNA,对其进行PCR鉴定,PCR反应体系及条件同1.2.1。于拔节期,设置5个Basta溶液浓度梯度(10、20、50、100和200 mg·L-1),用棉签将Basta溶液均匀涂抹于转基因小麦和野生型JW1叶片,5个重复,每隔2 d观察表型,一周后统计结果。利用确定的最适Basta浓度涂抹T1代各转基因植株,比较其和PCR鉴定结果,统计阳性植株和阴性植株的数量。
1.2.4 T1代转基因植株目标基因的表达量检测
选取7个T1代阳性植株(T1A5-18株系所对应的T1代植株由于错过取样时间而没有取样)和野生型JW1,取其开花前胚珠,采用试剂盒提取总RNA并反转录合成cDNA第一链。反转录体系:5×Reaction Buffer 4 μL,25 mmol·L-1MgCl24 μL,Random Primer 0.5 μL,qA5-R 0.5 μL,PCR Nucleotide Mix 1 μL,Ribonuclease Inhibitor 0.4 μL,Reverse Transcriptase 1 μL,RNA 8 μL,用ddH2O补至20 μL。反转录条件:25 ℃退火5 min,42 ℃延伸30 min,70 ℃反转录酶失活15 min。得到cDNA后通过qRT-PCR来检测目标基因的表达情况,内参基因选择GADPH来进行归一化处理。定量PCR以ddH2O为空白对照,野生型小麦JW1为阴性对照,每个样品设置3个重复。PCR反应体系:2× SYBR Green Master Mix 10 μL,10 μmol·L-1的上下游引物各0.25 μL,cDNA模板1 μL,ddH2O补至20 μL。PCR反应条件:95 ℃预变性3 min;95 ℃变性15 s,60 ℃延伸1 min,45个循环。目标基因检测所用引物为qA5-F/qA5-R(表1),内参基因引物为GADPH-F/GADPH-R(GADPH-F:5′-CCTTCCGTGTTCCCACTGTTG-3′;GADPH-R:5′-ATGCCCTTGAGGTTTCCCTC-3′)。qRT-PCR结束后,分析各植株的Ct值,计算出相对表达量。
1.2.5 T1代转基因植株籽粒大小的统计
待小麦种子成熟并干燥后,选取qRT-PCR分析所用7个阳性植株的主穗穗中部的10粒种子,用游标卡尺测定粒宽、粒长和粒厚,同时使用电子天平测量单粒重。
2 结果与分析
2.1 T0代转基因植株的鉴定结果
对T0代转基因植株进行了Bar试纸条检测及多次重复PCR检测,图2A中Bar试纸条检测结果显示,阴性对照(野生型JW1)和阴性转基因植株只显示一条控制线,而阳性转基因植株则可同时显出控制线和检测线;图2B中PCR检测检测结果显示,空白对照(ddH2O)和阴性对照(野生型JW1)基因组均没有扩增出预期目的条带,而阳性对照(pCAMBIA3301-TaCYP78A5质粒)与阳性转基因植株可扩增出与理论值一致的1 353 bp目的条带。从图2中可以看出,Bar试纸条检测结果与PCR检测结果一致,21株转基因再生苗中共鉴定出14株阳性植株。
2.2 T0代转基因植株的转入基因拷贝数
以T0A5-21的基因组DNA(5倍的稀释,设置5个梯度)作为模板进行实时定量PCR,得到Pinb和 TaCYP78A5基因的扩增曲线,并以模板浓度(C)的Log值和对应的Ct值为X轴和Y轴,得到Pinb和 TaCYP78A5基因的标准曲线(图3)。2个基因的扩增标准曲线的相关系数都为0.998,接近于1,所以得到的标准曲线可以用于测定T0代转基因植株的目标基因拷贝数。
分别以各转基因阳性植株的基因组DNA为模板,用内参基因Pinb和 TaCYP78A5基因特异引物和探针进行TaqMan实时定量PCR扩增,每个待测样品设定3个重复,得到其Ct值(表2),参照刘振华等[14]和Weng等[15]的方法进一步计算各转基因阳性植株中目标基因的拷贝数(表2)。农杆菌转化后代目标基因拷贝数均较低,14个阳性植株中除了2个植株的外源基因拷贝数为3和1个植株为7外,其余的均为1~2个拷贝,其中有6个单拷贝植株,约占总检测数目的一半。

T0A5-1至T0A5-24:转基因植株;M:DL2000;H2O:ddH2O;Y:pCAMBIA3301- TaCYP78A5。
T0A5-1-T0A5-24:Transgenic plants; M:DL2000; H2O:ddH2O; Y:pCAMBIA3301- TaCYP78A5.
图2 转基因植株Bar试纸条检测(A)和PCR检测(B)结果
Fig.2 Pictures of transgenic plants by Basta strips (A) and PCR (B)

图3 内参基因Pinb(A)和目标基因 TaCYP78A5(B)的扩增标准曲线
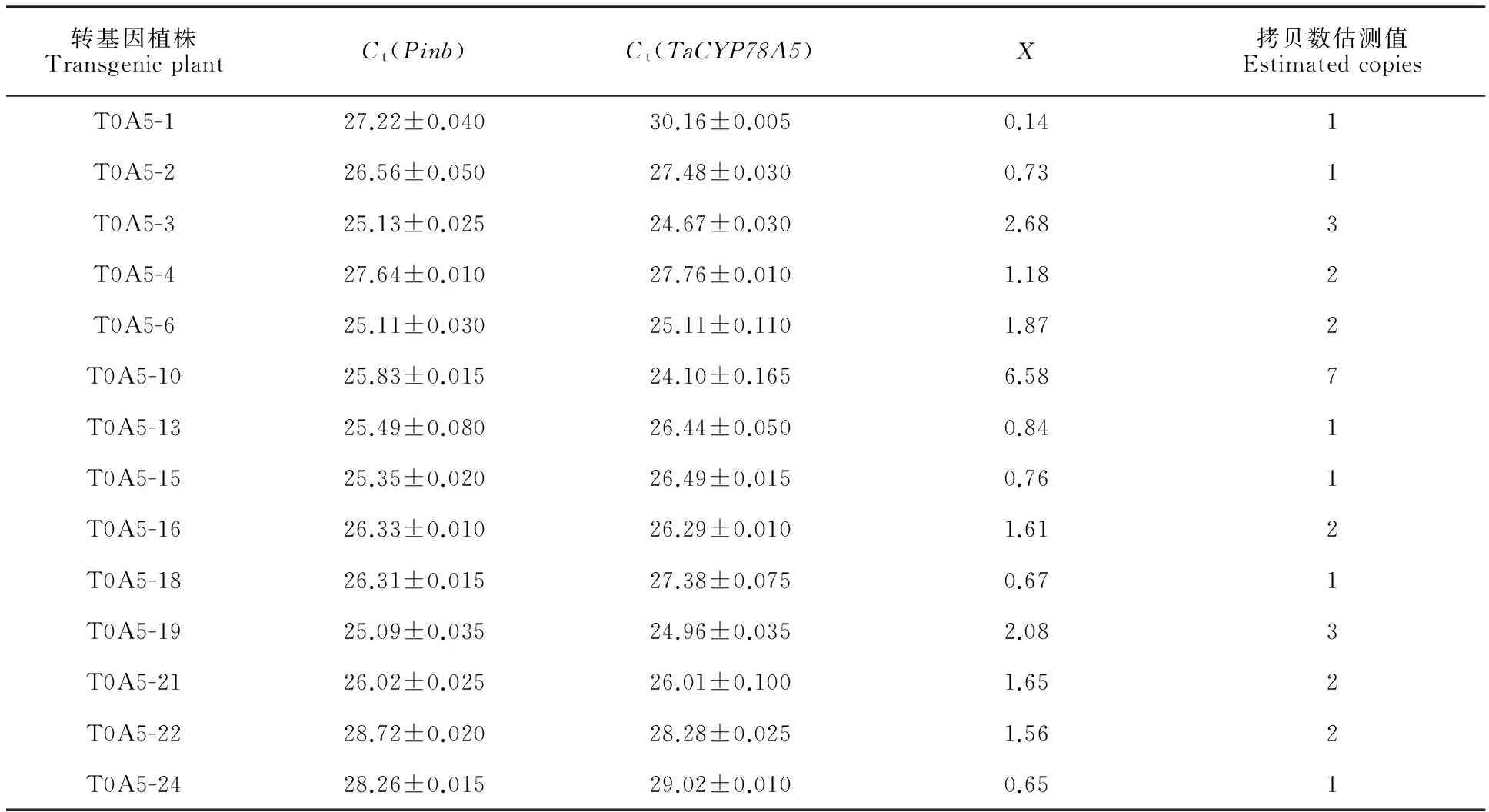
转基因植株TransgenicplantCt(Pinb)Ct(TaCYP78A5)X拷贝数估测值EstimatedcopiesT0A5-127.22±0.04030.16±0.0050.141T0A5-226.56±0.05027.48±0.0300.731T0A5-325.13±0.02524.67±0.0302.683T0A5-427.64±0.01027.76±0.0101.182T0A5-625.11±0.03025.11±0.1101.872T0A5-1025.83±0.01524.10±0.1656.587T0A5-1325.49±0.08026.44±0.0500.841T0A5-1525.35±0.02026.49±0.0150.761T0A5-1626.33±0.01026.29±0.0101.612T0A5-1826.31±0.01527.38±0.0750.671T0A5-1925.09±0.03524.96±0.0352.083T0A5-2126.02±0.02526.01±0.1001.652T0A5-2228.72±0.02028.28±0.0251.562T0A5-2428.26±0.01529.02±0.0100.651
X:拷贝数计算值。X:Caculated copies.
2.3 转基因植株最适Basta浓度
分别用10、20、50、100和200 mg·L-1五个浓度梯度的Basta溶液涂抹拔节期的阳性转基因小麦T1A5-2、阴性转基因小麦T1A5-5和野生型JW1,结果见图4。从图中可以看出,野生型JW1和转基因阴性对照T1A5-5的叶片在Basta浓度为10 mg·L-1时,几乎没有变化,随着Basta溶液浓度的增大,叶片的枯萎面积逐步增大,当浓度达到100 mg·L-1时,叶片涂抹部分已全部枯萎,而转基因阳性小麦株系叶片在Basta溶液浓度200 mg·L-1时仍保持正常生长。为了防止假阳性结果出现,选用200 mg·L-1的Basta来筛选T1代转基因植株,鉴定部分统计结果见表3。从表中可以看出,200 mg·L-1的Basta涂抹结果与前期PCR鉴定结果相一致,所以200 mg·L-1的Basta溶液叶片涂抹可用于大规模筛选含Bar基因的转基因植株。
2.4 转基因单拷贝插入的T0∶1株系目标基因的遗传分析
结合PCR检测与Basta叶片涂抹法的鉴定结果,统计出各T0∶1代株系所对应的T1代阳性植株与阴性植株的数量,其结果见表4。从表4中可以看出,目标基因是可遗传。
2.5 T1代转基因阳性植株目标基因的表达量
T1代阳性植株和野生型JW1开花前胚珠总RNA的RT-PCR分析结果如图5所示,从图中可以看出,与野生型JW1相比,各转基阳植株目标基因 TaCYP78A5表达量都极显著增加,且各植株间表达水平存在一定的差异,T1A5-2-8、T1A5-13-2、T1A5-13-9和T1A5-15-7表达量相对较高,而T1A5-1-4和T1A5-15-13表达量相对较低。
+:阳性; -:阴性。
+:Positive; -:Negative.
2.6 T1代转基因阳性植株的籽粒大小
各个阳性植株主穗中部籽粒粒宽、粒长、粒厚和粒重的统计结果如图6所示。从图中可以看出,与野生型相比,各转基因阳性植株的粒宽、粒重和粒厚均有所增加。在粒宽方面,除T1A5-13-2外,其他植株均极显著增加;在粒长方面,除T1A5-1-4极显著增加外,其他植株变化不明显;在粒厚方面,T1A5-1-4、T1A5-13-2和T1A5-15-7极显著增加,T1A5-13-9和T1A5-29-9显著增加,其余植株变化不明显;在粒重方面,所有植株都极显著增加。说明 TaCYP78A5基因在小麦中过表达能够增加小麦的籽粒大小,从而增加粒重。
3 讨 论
外源基因整合在转基因植株中的拷贝数是影响外源基因稳定性遗传和表达水平的一个重要因素。转基因植株中以单拷贝形式整合到基因组中的外源基因具有稳定遗传和较好的表达特性[16-17],而多拷贝则更容易导致外源基因的沉默[18]。因此,通过外源基因拷贝数测定,获得低拷贝数稳定遗传的转基因株系对于植物遗传改良有非常重要作用。检测外源基因拷贝数的方法主要有Southern blot和荧光定量PCR,Southern blot所需样品DNA量大、耗时、费力以及使用放射性同位素等缺点[19];荧光定量PCR快速灵敏,检测结果也与Southern blot相一致,所以应用起来较为方便[15]。本研究对小麦 TaCYP78A5过表达T0代阳性转基因植株利用TaqMan探针法进行了拷贝数检测,14个植株中共有6个单拷贝植株,5个双拷贝植株,其余3个植株在3拷贝以上,这与Zhang等[20]报道的农杆菌介导的遗传转化中外源基因主要以单拷贝形式整合到基因组中相一致。

表4 T0∶1代转基因株系目标基因的遗传分析Table 4 Genetic analysis of target genes in T0∶1 transgenic lines

**:P<0.01.
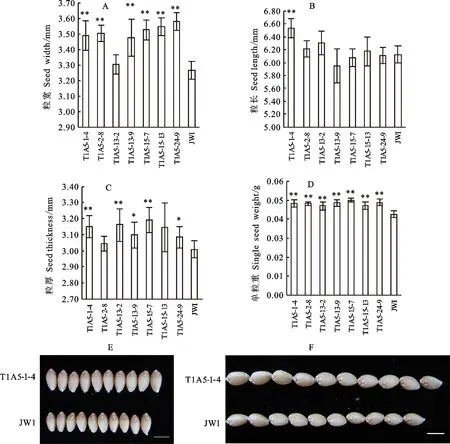
E和F中标尺为5 mm。 *:P<0.05; **:P<0.01; Scale bars are 5 mm in E and F.
在大规模转基因后代植株检测时,传统PCR步骤繁杂、结果不稳定,而Bar试纸条成本较高,即两种方法均存在一定的局限性。Sato等[21]对大豆转基因植株叶片涂抹Liberty除草剂所得到的结果与后期PCR和Southern blot结果相一致。李 欣等[22]使用叶片涂抹和植株喷洒对含Bar基因的转基因小麦进行鉴定,结果表明,200 mg·L-1的Liberty涂抹叶片法比PCR结果更精确,而植株喷洒Basta最适浓度为100 mg·L-1,Liberty最适浓度为150 mg·L-1。Basta溶液涂抹叶片成本低且效率高,不同转基因受体品种对Basta溶液的耐受性不同,因此本研究对小麦JW1转基因阳性植株的最适Basta浓度进行了摸索,结果表明,在拔节期涂抹200 mg·L-1的Basta溶液能够有效地筛选出阳性植株。
细胞色素P450在植物生长发育进程中发挥着相当重要的作用,其中, CYP78A是植物细胞色素P450家族中特有的一类[23]。近年来, CYP78A家族中一些成员都已被证实与植物生殖器官的发育有着密切的关系,如,拟南芥中的 CYP78A5、 CYP78A6、 CYP78A8和 CYP78A9[24],水稻中的 CYP78A13[11],大豆中的 CYP78A10和 CYP78A72[8,25],麻风树中的 CYP78A98[26],以及小麦中的 TaCYP78A3和 TaCYP78A5[5,9]。在拟南芥中,Adamski等[10]通过采用pKLU::vYFPer报告株系发现, KLUH/CYP78A5在胚珠发育时的内珠被中表达,并且KLU能促进胚珠中珠被细胞的增殖,进而来控制种子的大小。Xu等[4]通过水稻 CYP78A13突变体证实了该基因过表达导致籽粒增大。Zhao等[8]将 CYP78A72基因在拟南芥及大豆中过表达导致拟南芥和大豆种子均增大。Tian等[26]从麻风树中克隆出 CYP78A98基因,构建pINO:: JcCYP78A98过表达载体转化烟草, CYP78A98基因过表达使烟草籽粒大小和粒重增加,并且还提高了种子蛋白和脂肪酸的含量。Ma[5]等利用3种启动子(Camv35S、pINO和pKLU)启动 TaCYP78A5基因在拟南芥中过表达,结果显示, TaCYP78A5基因通过促进胚表皮细胞增殖来影响胚的发育,其表达量的高低影响着器官和种子的发育,过表达可导致拟南芥种子增大,初步证实小麦 TaCYP78A5基因正调控种子生长。本研究对 TaCYP78A5基因过表达转基因植株进行了鉴定分析,并统计了7个T1代阳性植株的籽粒大小和粒重,在粒宽、粒厚和粒重方面,阳性植株与野生型相比均增大,而在粒长方面,T1A5-13-9和T1A5-24-92与野生型相比变小,其余植株均增大。总体来说, TaCYP78A5基因在小麦中过表达能够增加小麦的籽粒大小并显著提高粒重,这为后期的小麦高产育种提供了科学依据。本研究材料为杂合株系,后期仍需要对转基因后代进行追踪检测以期获得转基因纯合株系,并能够对该基因的功能进行深入研究。
[1] 陈俊男,高润红,邓志英,等.小麦成熟胚组织培养体系优化及优良转化受体基因型的筛选[J].山东农业科学,2010(11):1.CHEN J N,GAO R H,DENG Z Y,etal.Optimization of regeneration system of tissue culture from mature embryos and selection of suitable transformation acceptor genotype in wheat [J].ShandongAgriculturalSciences,2010(11):1.
[2] 金松灿,王春平,孔欣欣,等.黄淮麦区小麦产量和生理性状的遗传增益研究[J].种子,2014,33(9):1.JIN S C,WANG C P,KONG X X,etal.Genetic gain for grain yield and its relational traits in Yellow-Huai River valley winter wheat region [J].Seed,2014,33(9):1.
[3]SHOMURA A,IZAWA T,EBANA K,etal.Deletion in a gene associated with grain size increased yields during rice domestication [J].NatureGenetics,2008,40(40):1023.
[4]XU F,FANG J,OU S,etal.Variations in CYP78A13 coding region influence grain size and yield in rice [J].Plant,Cell&Environment,2015,38(4):800.
[5]MA M,ZHAO H X,LI Z J,etal. TaCYP78A5 regulates seed size in wheat(Triticumaestivum) [J].JournalofExperimentalBotany,2016,67(5):1397.
[6]NELSON D R,SCHULER M A,PAQUETTE S M,etal.Comparative genomics of rice andArabidopsis.Analysis of 727 cytochrome P450 genes and pseudogenes from a monocot and a dicot [J].PlantPhysiology,2004,135(2):756.
[7]SCHULER M A,WERCK-REICHHART D.Fuctional genomics of P450s [J].AnualReviewofPlantBiology,2003,54(1):629.
[8]ZHAO B T,DAI A H,WEI H C,etal.ArabidopsisKLUhomologue GmCYP78A72 regulates seed size in soybean [J].PlantMolecularBiology,2016,90(1):33.
[9]MA M,WANG Q,LI Z J,etal.Expression of TaCYP78A3,a gene encoding cytochrome P450 CYP78A3 protein in wheat (TriticumaestivumL.),affects seed size [J].ThePlantJournal,2015,83(2):312.
[10]ADAMSKI N M,ANASTASIOU E,ERIKSSON S,etal.Local maternal control of seed size by KLUH/CYP78A5-dependent growth signaling [J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2009,106(47):20115.
[11]NAGASAWA N,HIBARA K I,HEPPARD E P,etal.GIANTEMBRYOencodes CYP78A13,required for proper size balance between embryo and endosperm in rice [J].ThePlantJournal,2013,75(4):592.
[12]CHAKRABARTI M,ZHANG N,SAUVAGE C,etal.A cytochrome P450 regulates a domestication trait in cultivated tomato [J].ProceedingsoftheNationalAcademyofSciences,2013,110(42):17125.
[13] 程绘绘. TaCYP78A3和 TaCYP78A5基因表达载体的构建及小麦的遗传转化[D].杨凌:西北农林科技大学,2015:21-23.CHENG H H.Expression vector construction of TaCYP78A3/TaCYP78A5 and wheat transformation [D].Yangling:Northwest A&F University,2015:21-23.
[14] 刘振华,刘香利,李冰冰,等.小麦转基因株系中外源脂氧合酶抑制基因(Loxi)拷贝数分析[J].农业生物技术学报,2014,22(8):949.LIU Z H,LIU X L,LI B B,etal.Copy number analysis of lipoxygenase RNAi gene (Loxi) in transgenic lines of wheat (Triticumaestivum) [J].JournalofAgriculturalBiotechnology,2014,22(8):949.
[15]WENG H B,PAN A H,YANG L T,etal.Estimating number of transgene copies in transgenic rapeseed by real-time PCR assay withHMGI/Yas an endogenous reference gene [J].PlantMolecularBiologyReporter,2004,22(3):289.
[16]CHENG M,FRY J E,PANG S Z,etal.Genetic transformation of wheat mediated byAgrobacteriumtumefaciens[J].PlantPhysiology,1997,115(3):971.
[17]VAUCHERET H,BÉCLIN C,ELMAYAN T,etal.Transgene-induced gene silencing in plants [J].ThePlantJournal,1998,16(6):651.
[18]SONG P,CAI C Q,SKOKUT M,etal.Quantitative real-time PCR as a screening tool for estimating transgene copy number in WHISKERSTM-derived transgenic maize [J].PlantCellReports,2002,20(10):948.
[19] 杨立桃,赵志辉,丁嘉羽,等.利用实时荧光定量PCR方法分析转基因水稻外源基因拷贝数[J].中国食品卫生杂志,2005,17(2):140.YANG L T,ZHAO Z H,DING J Y,etal.Estimating copy number of transgenes in transformed rice by real-time quantitative PCR [J].ChineseJournalofFoodHygiene,2005,17(2):140.
[20]ZHANG J,CAI L,CHENG J Q,etal.Transgene integration and organization in cotton (GossypiumhirsutumL.) genome [J].TransgenicResearch,2008,17(2):293.
[21]SATO S,XING A Q,YE X G,etal.Production of γ-linolenic acid and stearidonic acid in seeds of marker-free transgenic soybean [J].CropScience,2004,44(2):646.
[22] 李 欣,杜丽璞,殷桂香,等.转bar基因小麦和非转基因小麦抗除草剂鉴定方法比较[J].植物遗传资源学报,2012,13(4):596.LI X,DU L P,YIN G X,etal.Comparison of different methods forbargene detection in trangenic or non-transgenic wheat using herbcide as tool [J].JournalofPlantGeneticResources,2012,13(4):596.
[23]NELSON D R.Plant cytochrome P450s from moss to poplar [J].PhytochemistryReviews,2006,5(2-3):193.
[24]BAK S,BEISSON F,BISHOP G,etal.Cytochromes P450 [J].TheArabidopsisBook,2011,9(6):e0144.
[25]WANG X B,LI Y H,ZHANG H W,etal.Evolution and association analysis of GmCYP78A10 gene with seed size/weight and pod number in soybean [J].MolecularBiologyReports,2015,42(2):489.
[26]TIAN Y S,ZHANG M,HU X L,etal.Over-expression of CYP78A98 ,a cytochrome P450 gene fromJatrophacurcasL.,increases seed size of transgenic tobacco [J].ElectronicJournalofBiotechnology,2016,19:15.
Genetic and Functional Analysis of Transgenic Wheat Overexpressing TaCYP78A5 Gene
CHEN Zhiren1,MA Meng1,SHEN Yuxia1,WU Linnan1,LIU Xiangli1,2,ZHAO Huixian1,2
(1.College of Life Science,Northwest A&F University,Yangling,Shaanxi 712100,China;2.State Key Labaratory of Crop Stress Biology for Arid Areas,Northwest A&F University,Yangling,Shaanxi 712100,China)
In order to verify the function of TaCYP78A5 gene,which is associated with wheat (TriticumaestivumL.) grain size in the process of wheat grain development,the transgenic wheat lines with the TaCYP78A5 gene overexpression under the controlling of pINO promoter were used as materials. The copy number of target gene in T0generation transgenic plants was detected.The expression of the target gene of seven T1positive plants was tested and the grain size were measured. The results showed that out of 21 regenerated seedlings,14 positive plants were obtained by combining Bar strip test and PCR detection. The results of copy number detection showed that two and one transgenic plants had three and seven copies of target gene,respectively,and the rest eleven transgenic plants each had a copy number of 1 or 2,among which there were six single copy plants.Compared with the wild type JW1,the expression levels of the target gene in seven T1positive plants were significantly increased,the grain width and grain thickness were increased by different rates,and the grain weight were significantly increased.
Wheat; TaCYP78A5 gene;Bargene; Transgenic lines
10.7606/j.issn.1009-1041.2017.06.02
时间:2017-06-07
2017-01-16
2017-04-18
国家自然科学基金项目(31471482,31101205);西北农林科技大学基本科研业务费专项(Z109021565,Z109021568)
E-mail:chenzhiren0405@163.com
刘香利(E-mail:liuxianglii@163.com);赵惠贤(E-mail:hxzhao212@nwafu.edu.cn)
S512.1;S330
A
1009-1041(2017)06-0721-09
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20170607.1004.004.html
