藁城8901与豫麦50籽粒淀粉粒的粒度分布特征
2017-06-27闫素辉李文阳王振林
闫素辉,李文阳,王振林
(1.安徽科技学院农学院,安徽凤阳 233100; 2.山东农业大学作物生物学国家重点实验室,山东泰安 271018)
藁城8901与豫麦50籽粒淀粉粒的粒度分布特征
闫素辉1,李文阳1,王振林2
(1.安徽科技学院农学院,安徽凤阳 233100; 2.山东农业大学作物生物学国家重点实验室,山东泰安 271018)
为研究小麦籽粒发育过程中胚乳淀粉粒形成与粒度分布特征,以小麦品种藁城8901与豫麦50为材料,研究了籽粒胚乳淀粉粒形成、生长与分布特征。结果表明,花后4 d,小麦胚乳出现不同范围大小的淀粉粒,最大粒径8 μm。花后7 d,籽粒中淀粉粒增多增大,最大粒径20 μm左右。花后10~14 d,淀粉粒体积继续增大,并产生了一个新的小淀粉粒群体。花后17 d,淀粉粒以体积增大为主。花后21 d,淀粉粒最大粒径较成熟期变化较小。花后24 d,小于0.6 μm的淀粉粒数目急剧增加,大于0.6 μm的淀粉粒数目占比则明显减少,表明这一时期又产生了一个新淀粉粒群体(后期形成的B型淀粉粒)。花后24~28 d,小于0.6 μm的淀粉粒数目仍不断增加,而直径较大的淀粉粒数目增加减少,表明籽粒中新淀粉粒的产生仍在继续,但以小粒径淀粉粒为主。花后28 d至成熟期,最小粒径淀粉粒进一步生长,其他淀粉粒粒径变化相对较小。
小麦;藁城8901;豫麦50;淀粉粒;粒度分布
小麦淀粉主要存在于籽粒胚乳中,淀粉粒是小麦淀粉的存在形式[1],小麦淀粉粒根据粒径大小可分为>10 μm的A型淀粉粒和<10 μm的B型淀粉粒[2-3]。A、B型淀粉粒由于体积、表面积大小的差异,结合蛋白质、脂类和水等的能力存在显著差异[4-5]。B型淀粉粒数目增多,淀粉粒表面积增加,面粉吸水率升高,而A型淀粉粒数目增多,表面积降低,面粉吸水率下降[6-7]。
小麦籽粒A、B型淀粉粒结构的形成、发育是一个受籽粒生长调节的过程,籽粒不同生长阶段所形成的淀粉粒大小及组分组成有显著差异[8]。研究表明,在小麦开花后4 d或之前,最初的、微小的球形淀粉粒开始在质体中形成,并成为淀粉粒核,粒状核在成熟期形成大的、透镜状的A型淀粉粒[9]。B型淀粉粒产生于A型淀粉体[10-11]。通过对不同发育阶段的籽粒淀粉粒的发育进行观察,发现花后10~12 d在淀粉体上会产生一些突起,这些突起进一步分裂成单独的质体,B型淀粉粒由这些分裂出的质体形成[11]。
遗传分析表明,小麦籽粒中B型淀粉粒比例由加性基因效应决定[2]。强筋、弱筋小麦籽粒胚乳中淀粉粒大小分布、性质与结构存在显著差异[12]。本研究选用小麦品种藁城8901(强筋小麦)和豫麦50(弱筋小麦),研究小麦籽粒淀粉粒产生与生长发育特征,以揭示小麦籽粒淀粉形成与生长发育特征,为改善小麦淀粉品质的形成与调控提供理论参考。
1 材料与方法
1.1 试验设计
试验于2007年10月至2008年6月在山东农业大学农学实验站(36°09′N,117°09′E)进行。选用强筋小麦品种藁城8901(湿面筋含量为35.1%)和强筋品种豫麦50(湿面筋含量24.8%)为供试材料。试验地为壤土,前茬作物为玉米。田间种植密度为180×104株·hm-2,小区面积 9 m2(3 m×3 m)。田间管理按小麦一般高产栽培技术规程进行。在小麦开花期,选择生长、开花一致的主茎穗挂牌标记,于开花后4 d、7 d、10 d、12 d、14 d、17 d、21 d、24 d、28 d 和成熟期取样,用于籽粒胚乳淀粉粒分析。
1.2 测定项目与方法
1.2.1 淀粉粒的提取与分析
淀粉粒提取参考Peng 等[13]和戴忠民等[14]的方法,用美国贝克曼库尔特公司LS 13320 激光衍射粒度分析仪进行淀粉粒粒径分析。
1.2.2 淀粉粒形态、结构观察
参考Baldwin等[15]的方法,用日本JEOL-1200EX型透射电镜观察、拍照。
2 结果与分析
2.1 淀粉粒数目的变化
小麦籽粒发育的不同时期,淀粉粒数目呈单峰或双峰曲线,峰值出现在0.5~1.5 μm之间(图1)。花后4 d,已可检测到不同粒径大小的淀粉粒,淀粉粒数目呈单峰曲线分布,峰值出现在1.3 μm左右;最大粒径为8.5 μm,这些早期形成的淀粉粒将会发育成A型淀粉粒。花后7 d,淀粉粒数目呈单峰曲线,淀粉粒的数目和粒径继续增加,藁城8901的峰值出现在0.9 μm,最大粒径20.0 μm,豫麦50的峰值出现在0.5 μm,最大粒径为23 μm。
花后10 d,藁城8901淀粉粒数目呈双峰曲线,峰值分别出现在0.5 μm和1.3 μm,豫麦50淀粉粒的数目呈单峰曲线,峰值出现在0.8 μm。推测藁城8901在这一时期产生了一个新的小淀粉粒群体,即B型淀粉粒。说明强筋小麦藁城8901产生B型淀粉粒的时间较弱筋小麦豫麦50早。花后12 d,两个品种的淀粉粒数目分布均呈双峰曲线,峰值分别出现在0.45 μm和1.45 μm,此时藁城8901淀粉粒的最大粒径为27 μm,豫麦50淀粉粒的最大粒径为38 μm,<10 μm的淀粉粒数目占总数目的99.6%。花后14 d,两个品种淀粉粒的数目分布呈单峰曲线,峰值为1.1 μm。花后21 d,两个品种淀粉粒数目分布趋势基本一致,均呈单峰曲线,淀粉粒粒径基本达到最大,峰值出现在1.0 μm左右。这一趋势一直保持到花后35 d。
2.2 淀粉粒体积的变化
小麦胚乳淀粉粒的体积分布呈单峰或双峰曲线。花后4 d,两个品种的淀粉粒体积分布呈单峰曲线,峰值出现在3.0 μm左右(图2)。花后7 d,两个品种的淀粉粒的体积仍呈单峰曲线,峰值出现在10.3 μm,藁城8901和豫麦50的淀粉粒最大粒径分别为20.0 μm 和23.0 μm。
花后10 d,淀粉粒的粒径继续增大。藁城8901的淀粉粒体积分布呈双峰曲线,峰值出现在5.4 μm和15 μm。此时,<10 μm的淀粉粒体积占总体积的44.8%。花后12 d,豫麦50和藁城8901的淀粉粒体积分布均呈双峰曲线,峰值出现在5.4 μm和16.4 μm左右。此时,<10 μm的淀粉粒体积占总体积的34.6%。花后14 d,2品种淀粉粒体积分布均呈双峰曲线。藁城8901的峰值出现在3.4 μm和18 μm,豫麦50的峰值出现在2.3 μm和18 μm,2峰的分界点为7.1 μm。花后17 d,两品种淀粉粒的体积分布呈双峰曲线,峰值分别出现在2.8 μm和19.8 μm。花后21 d,淀粉粒体积分布的2个峰值分别出现在5.4 μm和21.7 μm,2个峰的分界点在8.6 μm,淀粉粒的粒径变化幅度较小。花后24 d,淀粉粒的体积分布呈双峰曲线。花后28 d,淀粉粒的体积呈双峰曲线,藁城8901的峰值分别为4.4和21.7 μm,豫麦50的峰值分别为5.4和23.8 μm。
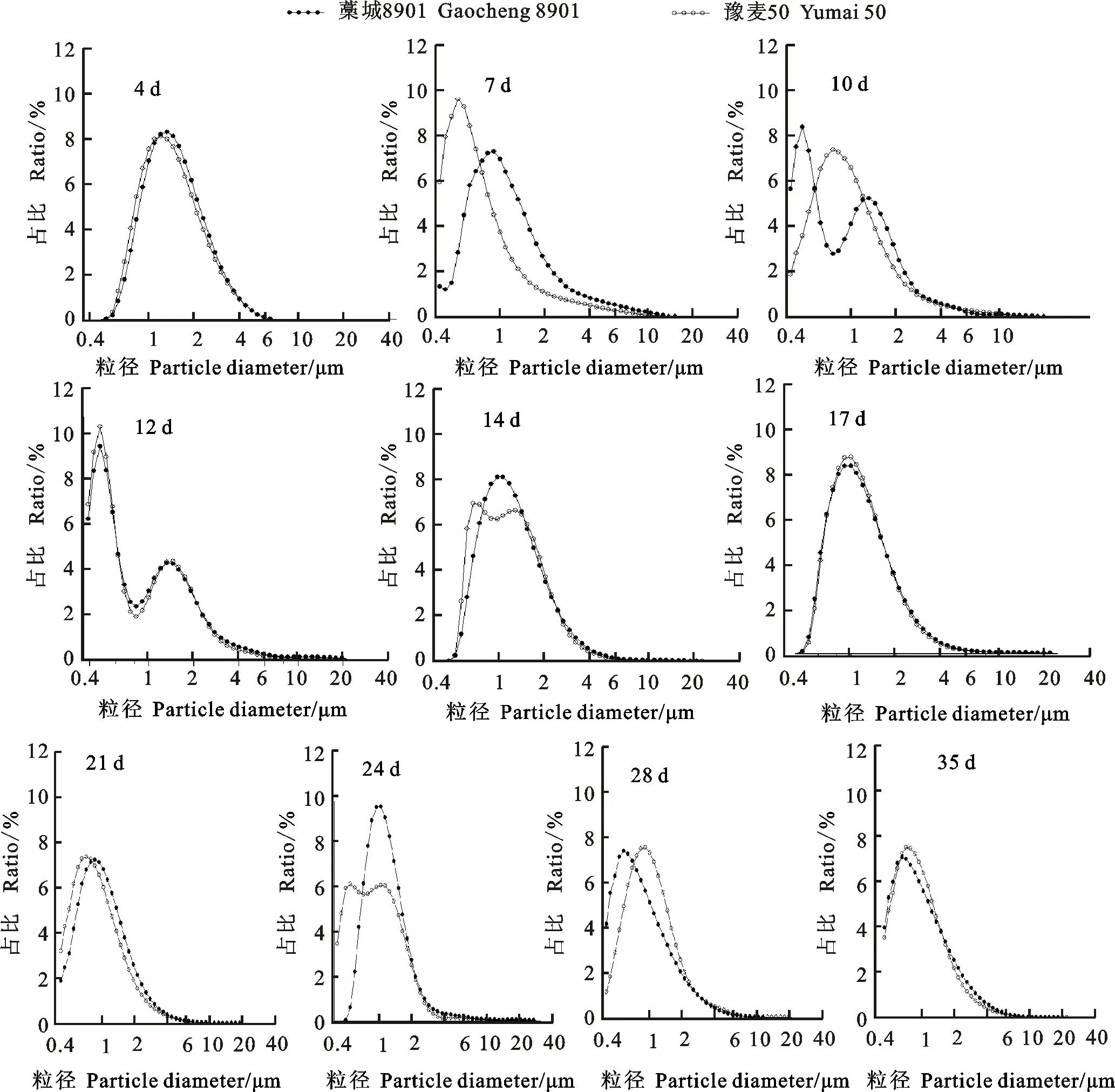
图1 小麦籽粒淀粉粒的数目分布
成熟期(花后35 d),淀粉粒最大粒径为42.5 μm~45 μm,藁城8901淀粉粒体积分布的峰值出现在4.4 μm和21.7 μm,2个峰的分界点在10 μm。自花后21 d至成熟,淀粉粒的最大粒径变化较小,结合数目分布变化趋势,说明在花后21 d,A型淀粉粒的体积和数目基本不再增加,而B型淀粉粒的数目和体积至成熟一直在增加。与弱筋小麦豫麦50相比,强筋小麦藁城8901有较高的B型淀粉粒体积分布比。

图2 小麦籽粒淀粉粒的体积分布
2.3 淀粉粒表面积的变化
花后4 d,两个品种淀粉粒的表面积分布呈单峰曲线,峰值出现在2.3 μm(图3)。花后7 d,两个品种淀粉粒的表面积分布呈双峰曲线,藁城8901的峰值出现在1.5 μm和7.8 μm,豫麦50的峰值分别出现在0.8 μm和7.1 μm,说明早期的淀粉粒发育进程不同。
花后10 d,藁城8901的淀粉粒表面积分布呈多峰分布,说明此时产生的淀粉粒群体大小不一;豫麦50淀粉粒表面积分布仍呈双峰曲线。花后12 d,两个品种的表面积分布均呈多峰分布。花后14 d,两个品种淀粉粒的表面积分布为双峰曲线,藁城8901峰值分别出现在1.9 μm和18 μm,2个峰的分界点为7.8 μm;豫麦50淀粉粒表面积峰值分别出现在1.9 μm和16.4 μm,两个峰的分界点为7.1 μm。花后17 d,两个品种淀粉粒的表面积分布呈双峰曲线,峰值出现在1.7 μm和18 μm,2个峰的分界点在7.8 μm。花后21 d,两个品种淀粉粒表面积分布的2个峰值出现在1.7 μm和19.8 μm,2个峰的分界点在11.3 μm;<10 μm的淀粉粒的表面积占总表面积的81.3%左右。花后24 d,淀粉粒的表面积呈3峰曲线。花后28 d,藁城8901的淀粉粒表面积分布呈双峰曲线,峰值分别出现在2.5 μm和21.7 μm,豫麦50呈3峰曲线分布,峰值分别出现在1.6 μm、5.0 μm和21.7 μm。
花后35 d,两个品种淀粉粒的表面积分布均呈双峰曲线,藁城8901的峰值分别出现在2.5 μm和19.8 μm,豫麦50的峰值分别出现在1.5 μm和19.8 μm,2个峰的分界点在10 μm。结合淀粉粒的数目、体积变化趋势,说明成熟期的淀粉粒存在2种类型,即A型(>10 μm)和B型(<10 μm)。
2.4 淀粉粒的发育
通过透射电镜对淀粉粒的发育进行观察,花后4 d可观察到淀粉粒的存在。花后7 d淀粉粒继续增大。花后10~14 d,在胚乳细胞中观察到新的小淀粉粒的产生。花后28 d,胚乳细胞中存在2种类型的淀粉粒,即A型和B型淀粉粒(图4)。
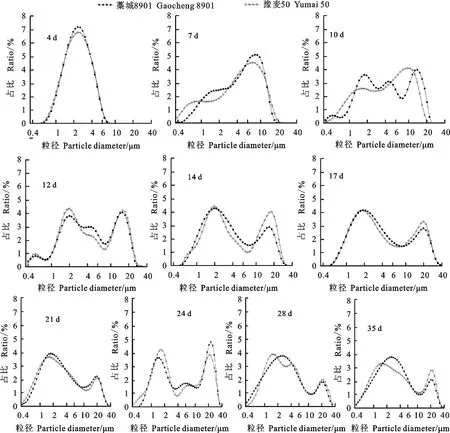
图3 小麦籽粒淀粉粒的表面积分布
3 讨 论
小麦籽粒中淀粉由胚乳中的双层膜结构的淀粉质体合成[10]。小麦胚乳淀粉的增长即是淀粉粒数目的增加和淀粉粒体积的增大[16]。小麦籽粒发育不同阶段所形成的淀粉粒的形态、体积和组成不同[7,17]。本研究对小麦籽粒不同灌浆阶段胚乳淀粉粒分布特征进行了分析,结果表明,花后4 d,小麦胚乳已出现不同大小的淀粉粒,最大粒径8 μm。花后7 d,淀粉粒增多增大,最大粒径20 μm左右。花后10~14 d,淀粉粒体积继续增大,并产生了一个新的小淀粉粒群体。花后17 d,淀粉粒以体积增大为主。花后21 d,淀粉粒最大粒径较成熟期变化较小。花后24 d,小于0.6 μm的淀粉粒数目急剧增加,大于0.6 μm的淀粉粒数目占比则明显减少,表明这一时期又产生了一个新淀粉粒群体(后期形成B型淀粉粒)。花后24~28 d,小于0.6 μm的淀粉粒数目仍不断增加,而粒径较大的淀粉粒占比不断减少,表明籽粒中新淀粉粒的产生仍在继续,但以小粒径淀粉粒为主。花后28 d至成熟,主要是小粒径淀粉粒的生长,其他粒径淀粉粒粒径的变化相对较小。说明小麦籽粒A型淀粉粒的形成和发育时期基本在花后21 d之前,而B型淀粉粒在成熟前,其体积和数目一直在增加。

图4 小麦胚乳第二层细胞淀粉粒的超微结构
Briarty等[18]认为,A型淀粉粒到成熟时最大直径为35 μm,而本研究所选品种籽粒A型淀粉粒直径可生长到45 μm。Bechtel等[16]研究表明,成熟期小麦淀粉粒的体积百分比呈三峰趋势。本研究则发现,成熟期小麦淀粉粒的体积分布呈双峰曲线。以上结果差异的原因可能是由于所选供试品种(如强筋、弱筋小麦品种差异)或淀粉粒分析测定方法不同。强筋小麦有较多的B型淀粉粒,淀粉粒和蛋白骨架结合致密,在磨面过程中淀粉粒破碎率较高,面团形成过程中吸水也较多,适合制作面包,而弱筋小麦与之相反,胚乳中有较多的A型淀粉粒,淀粉粒和蛋白骨架结合疏松,在磨面过程中在淀粉粒破碎率较低,面团形成过程中吸水也较少,适合制作糕点[19-20]。本研究发现,与弱筋小麦豫麦50相比,强筋小麦藁城8901产生B型淀粉粒时间较早,在成熟期有较高的B型淀粉粒体积分布比和较低的A型淀粉粒体积分布比。原因可能是弱筋小麦籽粒淀粉合成能力较强,其淀粉积累速率和合成相关酶活性均高于强筋小麦[19,21-22]。
[1] PAUL C.The structure of starch [J].Nature,1997,389:338.
[2] STODDARD F L.Survey of starch particle-size distribution in wheat and related species [J].CerealChemistry,1999,76:146.
[3] XIONG F,YU X,ZHOU L,etal.Effect of nitrogen fertilizer on distribution of starch granules in different regions of wheat endosperm [J].TheCropJournal,2014,2:47.
[4] ZHOU G,WILLETT J L,CARRIERE C J,etal.Effect of starch granule size on viscosity of starch-filled poly(hydroxy ester ether) composites [J].JournalofPolymersandtheEnvironment,2015,8:148.
[5] 李文阳,尹燕枰,时侠清,等.小麦籽粒A、B 型淀粉粒淀粉构成与糊化特性的比较[J].华北农学报,2011,26(1):138.LI W Y,YIN Y P,SHI X Q,etal.Comparison of starch composition and pasting properties between A and B type starch granule in wheat grain [J].ActaAgriculturaeBoreali-Sinica,2011,26(1):138.
[6] GAINES C S,RAEKER M O,TILLEY M.Associations of starch gel hardness,granule size,waxy allelic expression,thermal pasting,milling quality,and kernel texture of 12 soft wheat cultivars [J].CerealChemistry,2000,77(2):166.
[7] DAI Z,YIN Y,WANG Z.Starch granule size distribution from wheat cultivars under different water rengimes [J].CerealChemistry,2009,86:85.
[8] PARKER M L.The relationship between A-type and B-type starch granules in the developing endosperm of wheat [J].JournalofCerealScience,1985,3:271.
[9] EVERS A D.Scanning electron microscopy of wheat starch.Ⅲ.Granule development in the endosperm [J].Starch/Stärke,1971,23:159.
[10] 盛 婧,郭文善,朱新开,等.不同专用类型小麦籽粒淀粉粒形成过程[J].作物学报,2004,30(9):954.SHENG J,GUO W S,ZHU X K,etal.Starch granules development in grain endosperm of wheat for different end uses [J].ActaAgronomicaSinica,2004,30(9):954.
[11] EVERS A D,LINDLEY J.The particle-size distribution in wheat endosperm starch [J].JournaloftheScienceofFoodandAgriculture,1977,28:100.
[12] LI W Y,YAN S H,YIN Y P,etal.Comparison of starch granule size distribution between hard and soft wheat cultivars in eastern China [J].AgriculturalSciencesinChina,2008,7(8):913.
[13] PENG M,GAO M,ABDEL-AAL S M,etal.Separation and characterization of A- and B-type starch granules in wheat endosperm [J].CerealChemistry,1999,76:376.
[14] 戴忠民,王振林,张 敏,等.不同品质类型小麦籽粒淀粉粒度的分布特征[J].作物学报,2008,34(3):466.DAI Z M,WANG Z L,ZHANG M,etal.Starch granule size distribution in grains of strong and weak gluten wheat cultivars [J].ActaAgronomicaSinica,2008,34(3):466.
[15] BALDWIN P M,JEREMY A,DAVIES M C,etal.Holes in starch granules:Confocal,SEM and light microscopy studies of starch granule structure [J].Starch/Stärke,1994,46(9):343.
[16] BECHTEL D B,ZAYAS L,DEMPSTER R,etal.Size-distribution of starch granules isolated from hard red winter and soft red winter wheats [J].CerealChemistry,1990,70:239.
[17] 戴忠民,尹燕枰,王振林.鲁麦21和济南17胚乳发育过程中淀粉粒的动态变化[J].中国农业科学,2009,42(3):818.DAI Z M,YIN Y P,WANG Z L,etal.Starch particle size distribution in developing endosperm of wheat cultivars Lumai 21 and Jinan 17 [J].ScientiaAgriculturaSinica,2009,42(3):818.
[18] BRIARTY L G,HUGHES C E,EVERS A D.The developing endosperm of wheat-a stereological analysis [J].AnnalsofBotany,1979,44:645.
[19] CHIOTELLI E,MESTE M L.Effect of small and large wheat starch granules on thermo mechanical behavior of starch [J].CerealChemistry,2002,79:289.
[20] TURNBULL K M,RAHMAN S.Endosperm texture in wheat.[J].JournalofCerealScience,2002,36:333.
[21] LI W,YAN S,WANG Z.Effect of spikelet position on starch proportion,granule distribution and related enzymes activity in wheat grain [J].PlantSoilandEnvironment,2013,59:573.
[22] 闫素辉,王振林,戴忠民,等.两个直链淀粉含量不同的小麦品种籽粒淀粉合成酶活性与淀粉积累特征的比较[J].作物学报,2007,33(1):88.YAN S H,WANG Z L,DAI Z M,etal.Activities of enzymes involved in starch synthesis and accumulation in grains of two wheat cultivars with a different amylose content [J].ActaAgronomicaSinica,2007,33(1):88.
Size Distribution of Starch Granule in Grain of Wheat Cultivars Gaocheng 8901 and Yumai 50
YAN Suhui1,LI Wenyang1,WANG Zhenlin2
(1.College of Agronomy,Anhui Science and Technology University,Fengyang,Anhui 233100,China; 2.National Key Laboratory of Crop Biology,Shandong Agricultural University,Tai’an,Shandong 271018,China)
Formation and size distribution of starch granule of wheat endosperm in wheat cultivars,Gaocheng 8901 and Yumai 50,were studied during grain development in this study. The results showed that the starch granules initiated at 4 DAA(day after anthesis),and increased in size to a broad band of variously sized granules up to about 20 μm in diameter at 7 DAA. The diameter band of starch granules continued to grow up and a burst of synthesis created a new population of small starch granules during 10 to 14 DAA,which reduced the number proportion of big starch granules. At 17 DAA,starch granules in grain mainly increased in volume. At 21 DAA,the biggest diameter of starch granule ceased basically.At 24 DAA,the number of starch granules less than 0.6 μm increased sharply and the ratio of more than 0.6 μm reduced evidently,indicating that a new distinct class of small granules was synthesized at this stage,but the increase rate was reduced.From 28 DAA to maturity(35 DAA),the granules with the smallest diameter continued to develop,but not the other granules.
Winter wheat; Gaocheng 8901; Yumai 50; Starch granule; Size distribution
10.7606/j.issn.1009-1041.2017.06.10
时间:2017-06-07
2016-09-17
2016-10-27
安徽省自然科学基金项目(1408085QC54);国家星火计划项目(2015GA710018)
E-mail:yansh@ahstu.edu.cn
王振林(E-mail:zlwang@sdau.edu.cn)
S512.1;S311
A
1009-1041(2017)06-0794-07
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20170607.1004.018.html
