小麦旗叶大小及穗部相关性状的QTL定位
2017-06-27付金梅党政平赵婉莹杨雯晶刘明宇闵东红
付金梅,党政平,李 波,赵婉莹,赵 婉,杨雯晶,刘明宇,闵东红
(1.西北农林科技大学农学院/旱区作物逆境生物学国家重点实验室,陕西杨凌 712100;2.陕西省富平县种子技术推广中心,陕西富平 711700)
小麦旗叶大小及穗部相关性状的QTL定位
付金梅1,党政平2,李 波1,赵婉莹1,赵 婉1,杨雯晶1,刘明宇1,闵东红1
(1.西北农林科技大学农学院/旱区作物逆境生物学国家重点实验室,陕西杨凌 712100;2.陕西省富平县种子技术推广中心,陕西富平 711700)
为了发掘更多控制小麦旗叶大小及穗部相关性状的QTL,以兰考906和小偃81创制的133个F6~F7重组自交系为试验材料,在6个环境下利用SSR标记对旗叶大小及穗部相关性状进行QTL定位。结果表明,有202对SSR标记被用于构建遗传连锁图谱,图谱覆盖小麦21条染色体,全长1 678.93 cM,标记间平均距离8.30 cM。采用完备区间作图法共检测到30个QTL,分布在1B、2A、3D、4A、4B、4D、5D、6A、6B、6D和7D染色体上。其中,旗叶宽QTL有7个,穗长QTL有9个,小穗数QTL有5个,穗粒数QTL有5个,小穗着生密度QTL有4个,不同环境下单个QTL可解释的表型变异率为4.94%~23.14%,有14个QTL的表型贡献率大于10%,有8个QTL可在2个或2个以上环境中被检测到。其中, Qflw-4A在3个环境中被检测到,贡献率为10.13%~20.77%,是控制旗叶宽的稳定主效QTL; Qsl-4D.2在4个环境中被检测到,贡献率为12.58%~23.14%,是控制穗长的稳定主效QTL; Qker-5D在2个环境中被检测到,贡献率为11.44%~14.32%,是控制穗粒数的稳定主效QTL。这3个稳定主效QTL可作为改良叶宽和增加穗粒数的功能QTL作进一步研究。
小麦;重组自交系;旗叶大小;穗部性状;QTL
小麦旗叶及穗部性状与小麦产量关系密切。旗叶直接影响小麦的光合速率,其光合作用能为成熟籽粒提供20%~30%的干物质[1]。光合作用同时与小麦冠层结构相关,其株叶形态和株型结构的优化可提高旱地小麦产量[2]。研究表明,小麦产量与旗叶宽、旗叶面积呈正相关,与旗叶长呈负相关[3]。因此,对小麦旗叶大小相关QTL的研究有助于小麦理想株型的选育。小穗数、穗粒数是小麦产量构成的重要因子,而穗长与小麦产量构成因素密切相关。因此,对小麦穗部性状进行遗传研究和相关QTL检测能够进一步了解其遗传机理,以便在分子标记辅助育种中得到应用。
吕学莲等[4]定位了22个控制旗叶大小的QTL,主要位于1B、2D、3B、4B、7A、7B和7D染色体上。Wu等[5]将控制旗叶大小的QTL定位在2D、3B、4A、4D、5A、5B、6B、6D和7D染色体上,单个QTL在不同环境下可解释的表型变异率为2.49%~42.41%。Xue等[6]对面包小麦中控制旗叶宽的主效QTL TaFLW1进行研究,结果表明,该QTL位于5A染色体Xwmc492~Xwmc752之间。连俊方等[7]利用90K芯片定位了22个与旗叶相关的QTL,并在2A上定位了一个控制旗叶宽的稳定主效QTL,为理想株型的遗传改良提供了有用信息。Wu等[8]、Xu等[9]和Li等[10]在对小麦QTL的研究中都检测到了与穗部性状相关的QTL,并在3A和2D染色体上检测到了控制穗长的主效QTL。Börner等[11]通过总结前人研究发现,控制穗粒数的QTL主要分布在4AL和7D染色体上。王 瑾等[12]通过对导入系群体穗部性状QTL的精细定位发现,在1A、7B和7D染色体上都分布有控制小穗数和穗粒数的QTL。王佳佳等[13]利用MS-BC3F2:3和MT-F2:3群体将控制穗粒数的QTL定位在4BS染色体上。Patil等[14]将控制小穗数的QTL定位在4BL染色体上,可为进一步QTL精细定位提供帮助。
本研究以兰考906和小偃81创制的133个F6~F7重组自交系为材料,调查2年6个环境下旗叶大小和穗部性状的表型值,对旗叶长、旗叶宽、穗长、小穗数、穗粒数和小穗着生密度进行QTL定位,以期为进一步挖掘旗叶大小及穗部相关性状QTL位点和分子标记辅助育种奠定基础。
1 材料与方法
1.1 材 料
1.1.1 植物材料
兰考906是六倍体小黑麦和普通小麦经过染色体生物工程技术育成的1BL/1RS易位系,具有株型紧凑、穗大粒多、抗倒伏能力强、高抗白粉病和条锈病等特点。小偃81由小偃54和8602杂交而来,具有成穗率高、穗纺锤型、籽粒饱满度好、抗倒伏性较好等特点。两者在旗叶大小和穗部性状上有明显差异(图1),适合用于研究数量性状的QTL定位。因此,本研究以兰考906(父本)和小偃81(母本)杂交后通过单籽粒传法创制的133个F6~F7重组自交系(Recombinant inbred lines,RIL)群体为供试材料。以上所有材料均由西北农林科技大学农学院旱区作物逆境生物学国家重点实验室提供。
1.1.2 引 物
用于小麦旗叶大小及穗部性状QTL定位的1 237对SSR引物来源于Grain Genes 2.0(http://wheat.pw.usda.gov),并由北京奥科生物技术有限责任公司合成。
1.2 方 法
1.2.1 田间种植及表型鉴定
2014-2015年将试验材料分别种植于陕西杨凌(E1)、永寿(E2)、长武(E3),2行区,行长2 m,行距0.25 m。2015-2016年将试验材料分别种植于陕西杨凌(E4)、乾县(E5)、长武(E6),2行区,行长3 m,行距0.25 m。整个生育期管理同当地大田。

图1 兰考906和小偃81的表型
试验材料灌浆20 d后测量旗叶长、旗叶宽。试验材料成熟后单株收获,调查穗长、小穗数、穗粒数和小穗着生密度。调查时每个株系随机选取5株,取平均值作为相应性状的表型值。
1.2.2 DNA提取及PCR扩增
在小麦苗期取亲本及RIL群体叶片,用CTAB法[15]提取DNA。PCR反应体系为15 μL,包括2×Reaction Mix 7.5 μL、上下游引物(10 mmol·L-1)各1 μL、模板DNA(100 ng·μL-1)2 μL、Golden DNA Polymerase 0.2 μL和ddH2O 3.3 μL。PCR反应程序:94 ℃预变性5min;94℃变性45s,56~60℃退火45s,72 ℃延伸45 s,29个循环;72 ℃延伸10 min,4 ℃保存。利用6%非变性聚丙烯酰胺凝胶电泳检测PCR产物。
1.2.3 数据统计
利用SPSS 13.0对亲本的旗叶大小及穗部性状值进行差异显著性分析。利用Microsoft Excel对亲本及RIL群体的旗叶大小及穗部性状值进行描述性统计。
1.2.4 遗传连锁图谱构建和QTL定位
根据SSR引物的检测结果,将与母本兰考906相同的带型记为“A”,与父本小偃81相同的带型记为“B“,杂合或缺失记为“X”。利用QTL IciMapping V3.2(http://www.isbreeding.net)软件构建遗传连锁图谱,采用完备区间作图法进行QTL定位,设置步移为1.0 cM,P值临界值为0.001,LOD值为2.5,当LOD值大于2.5时则认为该处存在一个QTL[16]。以“Q+性状名称缩写+染色体编号+环境”的方式对QTL进行命名[17]。
2 结果与分析
2.1 亲本和RIL群体的表型变异
通过方差分析可知,兰考906和小偃81在旗叶宽、穗长、小穗数、穗粒数和小穗着生密度上存在极显著差异(表1)。同时,这些性状在RIL群体中的变异系数除了小穗数在E6下为9%外,其余均不小于10%,说明群体中表型变异丰富;在峰度值处,这些性状峰度值的绝对值都小于1,符合正态分布,可用于QTL定位(表2);而旗叶长在亲本间没有显著差异,群体间的变异系数较小,峰度值有偏离,不利于QTL定位,因此,本研究中不再对旗叶长进行讨论。
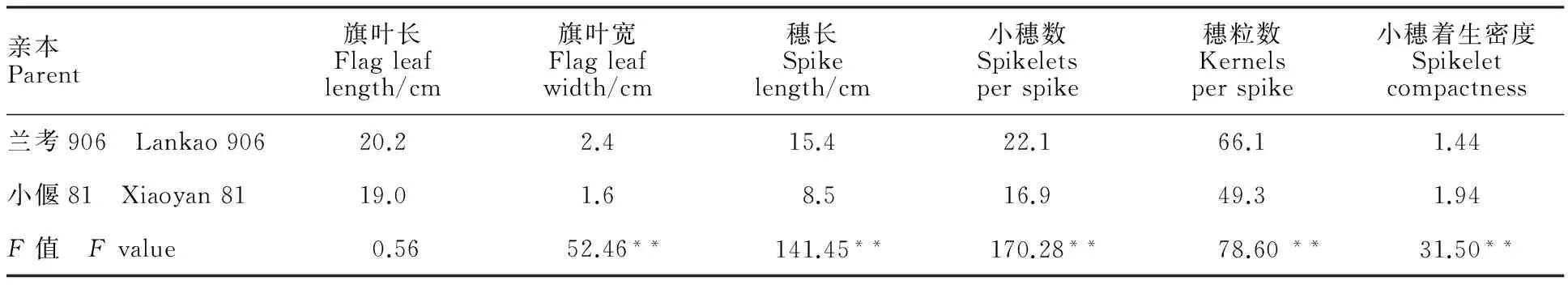
表1 兰考906和小偃81的表型变异Table 1 Phenotypic variation of Lankao 906 and Xiaoyan 81
**:P<0.01.
2.2 遗传连锁图谱的构建
利用1 237对SSR引物在亲本间进行多态性检测,有298对引物表现出多态性,多态率为24.1%。将298对多态性引物继续在RIL群体中扩增,有202对引物在群体中被检测出,且带型清晰,占多态性标记的67.8%。利用QTL IciMapping V3.2构建遗传连锁图谱,图谱全长1 678.93 cM,覆盖了小麦21条染色体,标记间平均距离8.30 cM。含有标记最多的是2A染色体,有30个;最少的是1D染色体,有4个。
2.3 QTL定位
采用完备区间作图法在6个环境下共检测到30个控制旗叶宽、穗长、小穗数、穗粒数和小穗着生密度QTL,分布在1B、2A、3D、4A、4B、4D、5D、6A、6B和7D染色体上(表3和图2)。分布最多的是2A染色体,有7个,最少的是3D、6A和6D染色体,各有1个。旗叶宽QTL有7个,穗长QTL有9个,小穗数QTL有5个,穗粒数QTL有5个,小穗着生密度QTL有4个。其中,8个QTL在2个或2个以上环境中被检测到,14个QTL的贡献率大于10%。
2.3.1 旗叶宽QTL
在5个环境中检测到了7个旗叶宽QTL,分别位于1B、2A、4A、4B、6B、6D和7D染色体上。 Qflw-2A.1在4个环境中被检出,贡献率为9.72%~17.86%。 Qflw-4A在3个环境中被检出,贡献率为10.13%~20.77%。 Qflw-6B和 Qflw-7D均在2个环境中检测到,贡献率分别为4.94%~7.67%和8.71%~9.93%。其他3个QTL均在单一环境中被检出。除了 Qflw-1B的加性效应来自小偃81外,其他QTL的加性效应全部来自兰考906。
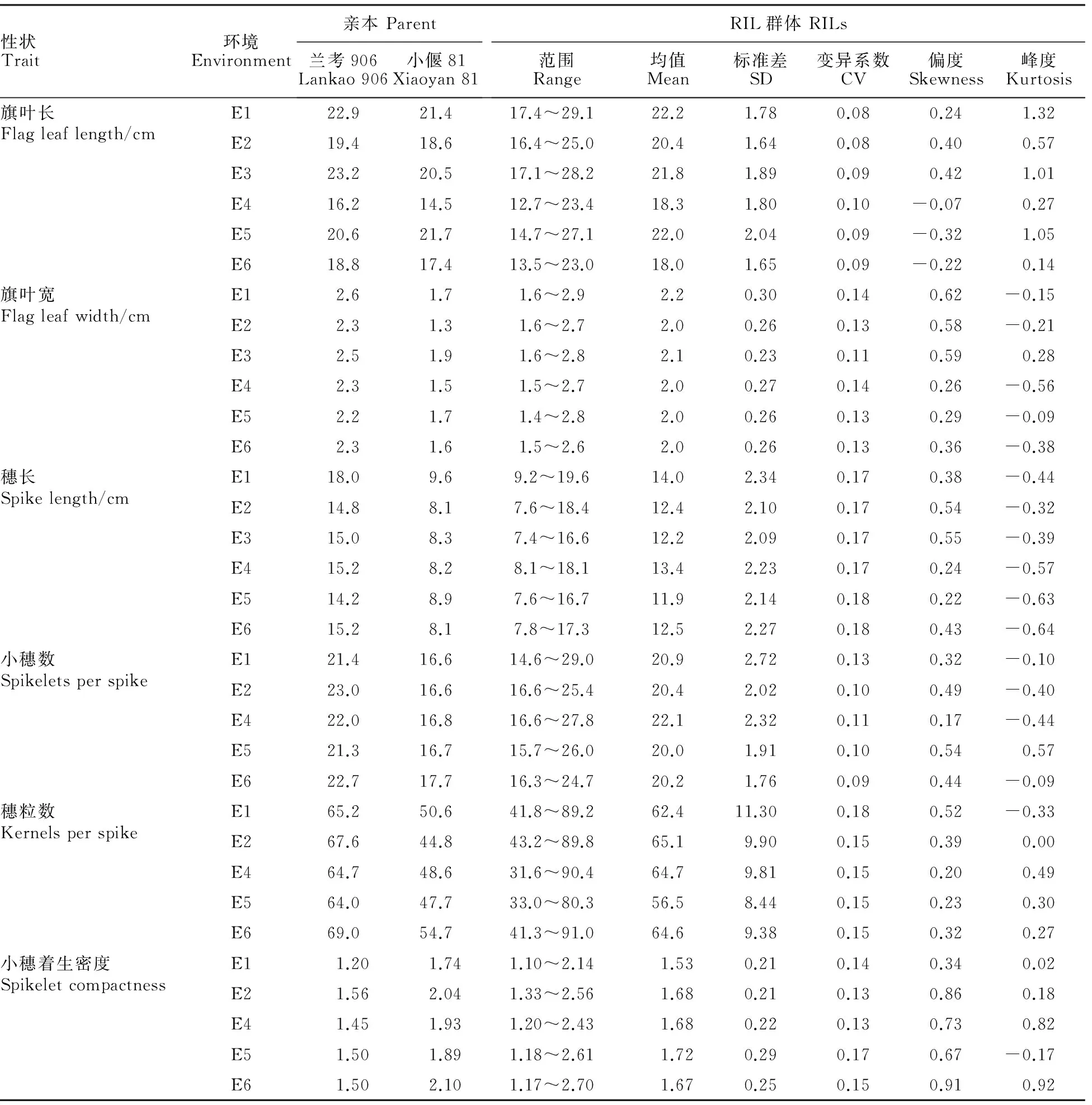
表2 RIL群体在不同环境下旗叶大小及穗部相关性状的变异Table 2 Phenotypic variation of flag leaf size and spike related traits in RILs in different environments
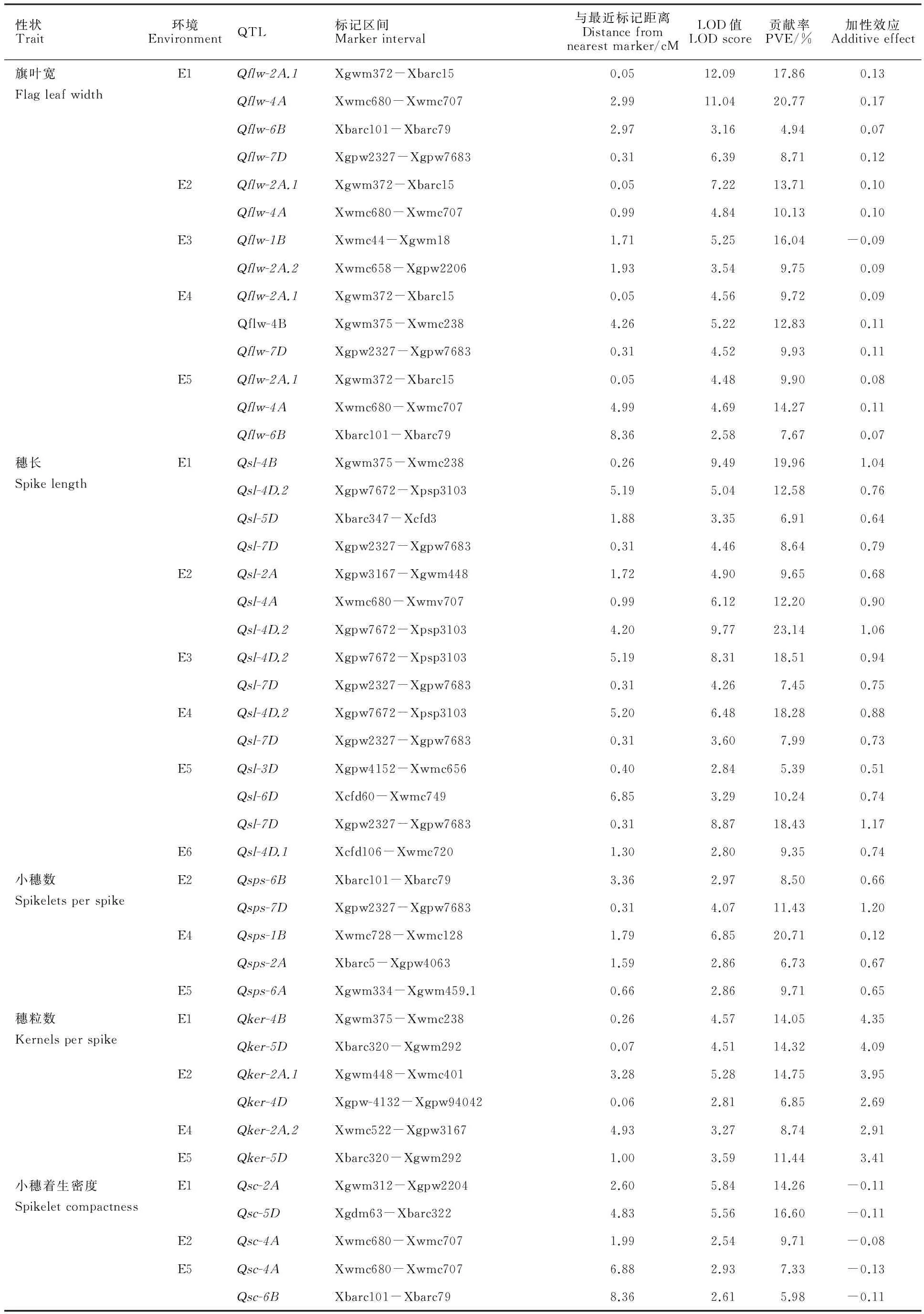
表3 不同环境下检测到的控制旗叶宽及穗部相关性状的QTLTable 3 QTLs for flag leaf width and spike related traits detected in different environments
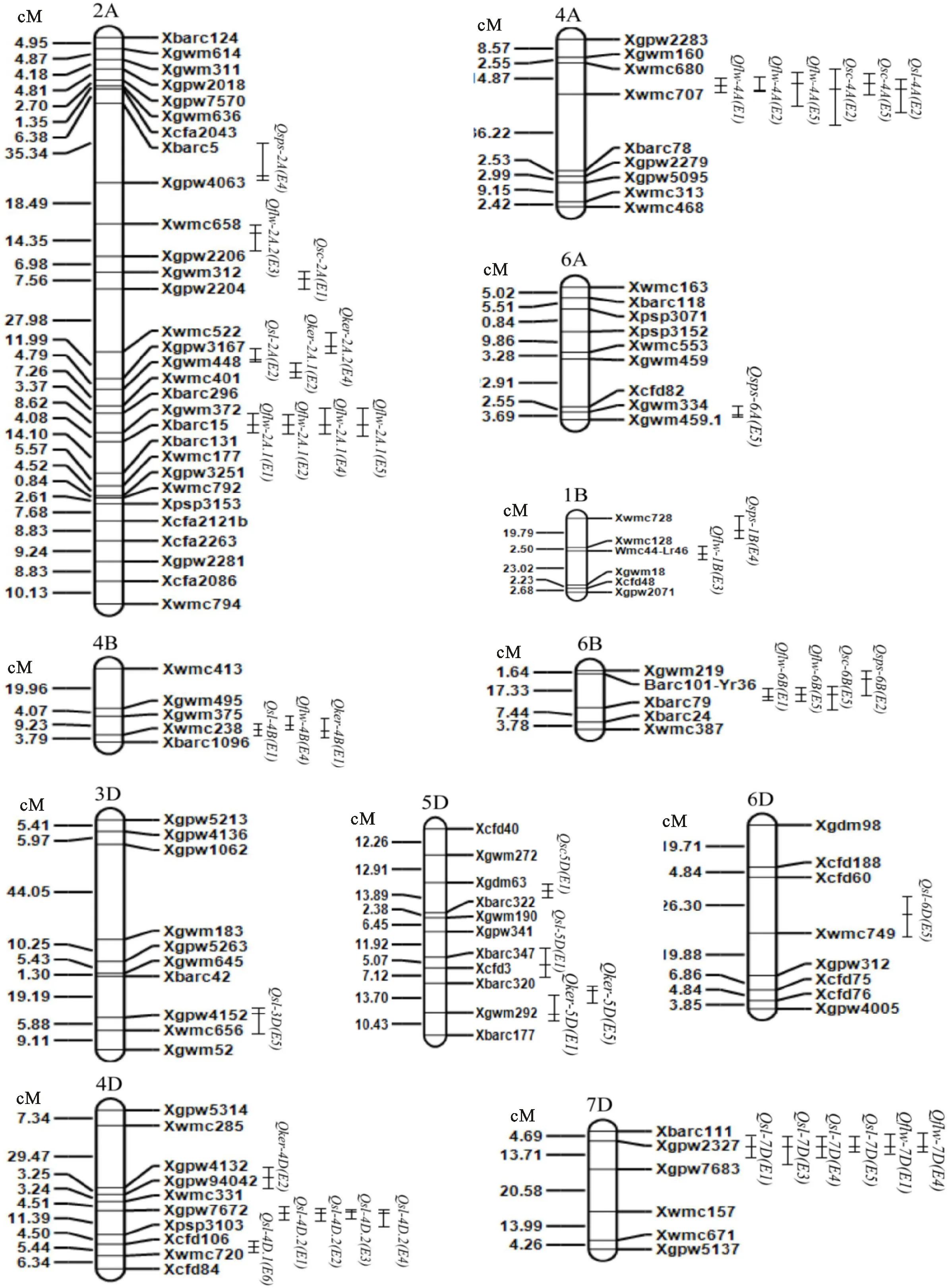
QTL左侧的竖线表示QTL的置信区间;竖线中间的横线表示LOD值最大处。
The vertical bar on the left side of the QTL represents the QTL confidence interval. The LOD maximum is shown by horizontal line in the middle of the vertical line.
图2 旗叶宽及穗部相关性状QTL在染色体上的分布
Fig.2 Distribution of QTLs for flag leaf width and spike related traits on chromosomes
2.3.2 穗长QTL
在6个环境中共检测到了9个穗长的QTL,分布在2A、3D、4A、4B、4D、5D、6D和7D染色体上。 Qsl-4D.2和 Qsl-7D均在4个环境中被检出,其中, Qsl-4D.2贡献率为12.58%~23.14%。另外7个QTL均只在一个环境中被检出,表型贡献率为5.39%~19.96%。
2.3.3 小穗数QTL
在3个环境中检测到了5个小穗数QTL,1B、2A、6A、6B和7D染色体上各有1个,且都只在1个环境中被检测到。 Qsps-7D和 Qsps-1B可解释的表型变异率较大,分别为11.43%和20.71%。 Qsps-2A的表型变异率最小,为6.73%。
2.3.4 穗粒数QTL
在4个环境中检测到了5个控制穗粒数的QTL,位于2A、4B、4D和5D染色体上。其中, Qker-5D在2个环境中被检测到,另外4个QTL在单一环境中被检出,贡献率为6.85%~14.75%。除 Qker-2A.2和 Qker-4D外,其他3个穗粒数QTL的表型贡献率均大于10%。
2.3.5 小穗着生密度QTL
在3个环境中共检测到了4个小穗着生密度QTL,分别位于2A、4A、5D和6B染色体上。 Qsc-4A在2个环境中被检测到,贡献率为7.33%~9.71%。另外3个都只在一个环境中被检测到,贡献率为5.98%~16.60%,其中, Qsc-2A和 Qcd-5D的贡献率在10%以上。小穗着生密度QTL的加性效应全部来自小偃81。
3 讨 论
本研究在6个环境中共定位了30 个与旗叶宽和穗部性状相关的QTL,有14个QTL的表型贡献率大于10%,是主效QTL;8个QTL在多个环境中被检测到,另外22个QTL都只在单一环境中检测到,说明这些数量性状的表达在一定程度上受到了环境的影响。与前人的研究相比较发现,本研究定位的QTL位点有些与之前的相同,有些是新发现的QTL位点,这可能是因为各个研究所涉及的标记和作图群体不同,同时,旗叶宽、穗长、小穗数、穗粒数和小穗着生密度是数量性状,很大程度上受到环境、基因型与环境互作的影响[18]。
与吕学莲等[4]和Fan等[19]在4B和6B染色体上定位的旗叶宽QTL相比较,本研究也在多个环境中检测到了位于4B和6B染色体上控制旗叶宽的QTL,但所处区间不同,是新发现的QTL位点,说明在4B和6B染色体上可能存在控制旗叶宽的重要QTL位点。
Wu等[8]和Xu等[9]将控制穗长的主效QTL定位在了3A和2D染色体上,这与本研究的结果不一致。王 瑾等[12]和Wang等[20]分别在4A染色体Xwmc680-Xwmc491和Xbarc170-Xwmc707上检测到穗长QTL Qsl.caas-4A和 Qsl.wa-4A,本研究也在4A染色体上检测到了穗长主效QTL Qsl-4A,所处区间为Xwmc680-Xwmc707,说明控制穗长的QTL可能与Xwmc680和Xwmc707连锁。
Patil等[14]和王佳佳等[13]分别在2A和4B染色体上定位到的小穗数QTL Qsps.macs-2A和穗粒数QTL Qker-4B与本研究在2A和4B染色体上定位到的小穗数和穗粒数QTL所属区段不同,是不同的QTL位点。张坤普等[21]在5D染色体上检测到的小穗着生密度QTL Qsc-5D与本研究在5D染色体上检测到的小穗着生密度QTL Qsc-5D都与标记Xgdm63连锁,具有较高的应用价值。
Patil等[14]、Heidari等[22]和刘 凯等[23]都在相应的研究中检测到了一因多效QTL,即QTL富集区。本研究在4A、4B、6B和7D染色体上各发现了一个一因多效QTL。4A上的QTL富集区与旗叶宽、穗长和小穗着生密度相关;4B上的QTL富集区控制着旗叶宽、穗长和穗粒数;6B上的QTL富集区控制旗叶宽、小穗数和小穗着生密度;7D上的QTL富集区控制旗叶宽、穗长和小穗数。王佳佳等[13]也在4B上检测到一个QTL富集区,控制的是穗长、穗粒数和小穗数,但与本研究所处区间不同。Heidari等[22]利用DH群体定位了一个同时与穗长和小穗着生密度紧密连锁的标记Xgwm261,而本研究并没有得出类似结果。出现这种结果可能是由基因引起的,也可能是受到了环境的影响,还需要做进一步验证。
[1] 曹树青,赵永强,温家立,等.高产小麦旗叶光合作用及与籽粒灌浆进程的关系的研究[J].中国农业科学,2000,33(6):19.CAO S Q,ZHAO Y Q,WEN J L,etal.Studies on photosynthesis in flag leaves and it's relation to grain filling course of high-yield wheat [J].ScientiaAgriculturaSinica,2000,33(6):19.
[2] 李朴芳,程正国,赵 鸿,等.旱地小麦理想株型研究进展[J].生态学报,2011,31(9):2631.LI P F,CHENG Z G,ZHAO H,etal.Current progress in plant ideo type research of dryland wheat(TriticumaestivumL.) [J].ActaEcologicaSinica,2011,31(9):2631.
[3] 何贤芳,赵 莉,朱昭进,等.小麦株型性状与产量性状及生物学产量关系的初步研究[J].园艺与种苗,2011(4):6.HE X F,ZHAO L,ZHU Z J,etal.Preliminary study on the relationship between plant-type traits and yield components & biomass in wheat [J].Horticulture&Seed,2011(4):6.
[4] 吕学莲,白海波,董建力,等.春小麦旗叶大小相关性状的QTL定位分析[J].麦类作物学报,2016,36(12):1587.LÜ X L,BAI H B,DONG J L,etal.QTL mapping for size traits of flag leaf in spring wheat [J].JournalofTriticeaeCrops,2016,36(12):1587.
[5]WU Q H,CHEN Y X,FU L,etal.QTL mapping of flag leaf traits in common wheat using an integrated high-density SSR and SNP genetic linkage map [J].Euphytica,2016,208(2):337.
[6]XUE S L,XU F,LI G Q,etal.Fine mapping TaFLW1,a major QTL controlling flag leaf width in bread wheat(TriticumaestivumL.) [J].TheoreticalandAppliedGenetics,2013,126(8):1941.
[7] 连俊方,张德强,武炳瑾,等.利用90K基因芯片进行小麦旗叶相关性状的QTL定位[J].麦类作物学报,2016,36(6):689.LIAN J F,ZHANG D Q,WU B J,etal.QTL mapping for flag traits using an integrated high-density 90K genotyping chip [J].JournalofTriticeaeCrops,2016,36(6):689.
[8]WU X Y,CHENG R R,XUE S L,etal.Precise mapping of a quantitative trait locus interval for spike length and grain weight in bread wheat(TriticumaestivumL.) [J].MolecularBreeding,2014,33(1):129.
[9]XU Y F,WANG R F,TONG Y P.Mapping QTLs for yield and nitrogen-related traits in wheat:Influence of nitrogen and phosphorus fertilization on QTL expression [J].TheoreticalandAppliedGenetics,2014,127(1):59.
[10]LI C L,BAI G H,CARVER B F,etal.Single nucleotide polymorphism markers linked to QTL for wheat yield traits [J].Euphytica,2015,206(1):89.
[11]BÖRNER A,SCHUMANN E,FURSTE A,etal.Mapping of quantitative trait loci determining agronomic important characters in hexaploid wheat(TriticumaestivumL.) [J].TheoreticalandAppliedGenetics,2002,105(6-7):921.
[12] 王 瑾,廖祥政,杨学举,等.人工合成小麦Am3大穗多粒QTL的发掘与利用[J].植物遗传资源学报,2008,9(3):277.WANG J,LIAO X Z,YANG X J,etal.Mapping of large-spike and much-kernel QTL by using a synthetic wheat Am3 as donor [J].JournalofPlantGeneticResources,2008,9(3):277.
[13] 王佳佳,王盈盈,张照贵,等.小麦穗部性状和株高的QTL定位[J].分子植物育种,2015,13(1):77.WANG J J,WANG Y Y,ZHANG Z G,etal.Mapping QTLs for the traits related to spike and plant height in wheat [J].MolecularPlantBreeding,2015,13(1):77.
[14]PATIL R M,TAMHANKAR S A,OAK M D,etal.Mapping of QTL for agronomic traits and kernel characters in durum wheat(TriticumdurumDesf.) [J].Euphytica,2013,190(1):117.
[15]MURRAEY M G,THOMPSON W F.Rapid isolation of high molecular weight plant DNA [J].NucleicAcidsResearch,1980,8(19):4321.
[16] 王建康.数量性状基因的完备区间作图方法[J].作物学报,2009,35(2):239.WANG J K.Inclusive composite interval mapping of quantitative trait genes [J].ActaAgronomicaSinica,2009,35(2):239.
[17]MCCOUCH S R,CHO Y G,YANO M,etal.Report on QTL nomenclature [J].RiceGeneticsNewsletter,1997,14(11):11.
[18]SIMGH G,BHULLAR G S,GILL K S.Genetic control of grain yield and its related traits in bread wheat [J].TheoreticalandAppliedGenetics,1986,72(4):536.
[19]FAN X L,CUI F,ZHAO C H,etal.QTLs for flag leaf size and their influence on yield-related traits in wheat(TriticumaestivumL.) [J].MolecularBreeding,2015,35(1):24.
[20]WANG J S,LIU W H,WANG H,etal.QTL mapping of yield-related traits in the wheat germplasm 3228 [J].Euphytica,2011,177(2):277.
[21] 张坤普,徐宪斌,田纪春.小麦籽粒产量及穗部相关性状的QTL定位[J].作物学报,2009,35(2):270.ZHANG K P,XU X B,TIAN J C.QTL mapping for grain yield and spike related traits in common wheat [J].ActaAgronomicaSinica,2009,35(2):270.
[22]HEIDARI B,SAYED-TABATABARI B E,SAEIDI G,etal.Mapping QTL for grain yield,yield components,and spike features in a double haploid population of bread wheat [J].Genome,2011,54(6):517.
[23] 刘 凯,邓志英,李青芳,等.利用高密度SNP遗传图谱定位小麦穗部性状基因[J].作物学报,2016,42(6):820.LIU K,DENG Z Y,LI Q F,etal.Mapping QTLs for wheat panicle traits with high density SNP genetic map [J].ActaAgronomicaSinica,2016,42(6):820.
QTL Mapping for Flag Leaf Size and Spike Related Traits in Wheat(TriticumaestivumL.)
FU Jinmei1,DANG Zhengping2,LI Bo1,ZHAO Wanying1,ZHAO Wan1,YANG Wenjing1,LIU Mingyu1,MIN Donghong1
(1.College of Agronomy,Northwest A&F University/State Key Laboratory of Crop Biology for Arid Areas,Yangling,Shaanxi 712100,China; 2.Fuping Seed Technology Promotion Center in Shaanxi Province,Fuping,Shaanxi 711700,China)
In order to explore more QTLs that control flag leaf size and spike related traits in wheat,a population of 133 F6:7recombinant inbred lines(RILs) derived from a cross between Lankao 906 and Xiaoyan 81 was used to detect the QTLs for flag leaf size and spike related traits in six environments. The results showed that 202 SSR markers were used to construct genetic linkage maps,covering 21 wheat chromosomes,with the full mapping length of 1 678.93 cM and an average genetic distance of 8.30 cM between the markers. A total of 30 QTLs were detected on 1B,2A,3D,4A,4B,4D,5D,6A,6B,6D and 7D chromosomes by Inclusive Composite Interval Mapping(ICIM,LOD>2.5). There were seven QTLs for flag leaf width(FLW),nine QTLs for spike length(SL),five QTLs for spikelets per spike(SPS),five QTLs for kernels per spike(KER),and four QTLs for spike compactness(SC),with the phenotypic variations ranging from 4.94% to 23.14% in different environments. Among these QTLs, Qflw-4A was detected in three environments with phenotypic variations ranging from 10.13% to 20.77%,which was a stable and major QTL associated with FLW. Qsl-4D.2 was detected in four environments with phenotypic variations ranging from 12.58% to 23.14%,which was a stable and major QTL associated with SL. Qker-5D was detected in two environments with phenotypic variations ranging from 11.44% to 14.32%,which was a stable and major QTL associated with KER. The three stable and major QTLs on chromosomes 4A,4D and 5D identified in this study might be useful for marker-assisted selection of flag leaf size and spike traits in wheat breeding.
Wheat; RILs; Flag leaf size; Spike traits; QTL
10.7606/j.issn.1009-1041.2017.06.01
时间:2017-06-07
2017-01-20
2017-02-20
国家重点研发计划项目(2016YFD0101802)
E-mail:fjm620@foxmail.com
闵东红(E-mail:mdh2493@126.com)
S512.1;S330
A
1009-1041(2017)06-0713-08
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20170607.1004.002.html
