福建南日岛南部水域鱼类群落结构及多样性
2017-06-22徐兆礼陈佳杰
管 伟,徐兆礼,陈佳杰
1 中国水产科学研究院东海水产研究所农业部海洋与河口渔业重点开放实验室,上海 200090 2 上海海洋大学海洋科学学院,上海 201306
福建南日岛南部水域鱼类群落结构及多样性
管 伟1,2,徐兆礼1,*,陈佳杰1
1 中国水产科学研究院东海水产研究所农业部海洋与河口渔业重点开放实验室,上海 200090 2 上海海洋大学海洋科学学院,上海 201306
为探究南日岛南部水域鱼类群落结构及多样性特征,在2009年冬季,2010年春季与夏季于南日岛南部海域展开了3个航次的渔业资源综合调查.采用Bray-Curtis聚类方法,对南日岛南部海域鱼类群落进行划分,并在此基础上分析了不同群落鱼类种类数、数量密度、生态类群和多样性特征,讨论了水团变化对鱼类群落格局划分及多样性特征的影响。结果显示:春季调查海域的鱼类可划分为北部沿岸群落和南部近海群落,这两个种群种类数分别是27和32种,密度分别为7.47×103尾/km2和24.34×103尾/km2,多样度H′值分别为2.71和1.97,南部近海群落中暖水种数量密度均高于北部沿岸群落,而暖温种相反。与春季不同,夏季由西至东划分为西部沿岸群落和东部近海群落,种类数分别为55和27种,密度分别为329.24×103尾/km2和106.47×103尾/km2,H′值分别为1.97和1.62,西部沿岸群落中沿岸种和近海种数量密度均高于东部近海群落。冬季调查海域鱼类可划分为北部沿岸群落和南部近海群落,种类数分别为25和32种,密度分别为9.96×103尾/km2和28.85×103尾/km2,H′值分别为2.45和2.33,南部近海群落暖水种与暖温种数量密度均高于北部沿岸群落。冬、春季时,海峡暖水影响的海域鱼类种类数、数量密度、多样度以及暖水种的数量密度均高于浙闽沿岸水控制的海域;夏季,西部沿岸径流影响的海域鱼类种类数、数量密度、多样度以及暖水种的数量密度均高东部海峡暖水影响的海域。可见鱼类群落分布特征与季节性变化的水团有关。
鱼类;群落;多样性;格局;水团;南日岛南部海域
水团的运动是海洋环境产生局部变化的主要因素,这些因素的变化对鱼类的分布、洄游和集群等影响极大[1]。国际上,关于水团对鱼类群落特征的研究非常活跃。如Lazzari等[2]对缅因州墨西哥暖流影响下近海鱼类群落结构的季节和年份变化进行了研究,Ayvazian等[3]研究了澳大利亚西南海岸寒流区的鱼类群落,Aschan等[4]研究了1992—2004年间来自北大西洋的暖水团和北极圈冷水团的对巴伦支海鱼类群落结构时空变化的影响。
南日岛南部海域位于台湾海峡中部,是东海与南海的过渡区,也是不同鱼类区系的过渡带[5]。浙闽沿岸水和海峡暖水的势力在不同季节此消彼长[6],使得该海域鱼类群落结构呈现一定的变化。国内已有学者对南日岛邻近海域鱼类区系、多样性和种类组成进行大尺度研究。如陈永俊[5]等研究了台湾海峡鱼类组成及其生态区系,宗普庆等[7]研究了台湾海峡游泳动物种类组成及其多样性,林龙山等[8]分析了台湾海峡夏、秋季游泳动物资源分布及群落结构。这些研究为了解台湾海峡中部鱼类生态学提供了基础,但大都针对于种类组成、资源分布或群落结构特征与水深关系的描述和简单分析,并未分析水团对群落结构的影响。而且,对台湾海峡大陆沿岸近海海域鱼类群落结构特征的研究多限于河口、海湾,如徐兆礼[9]研究了兴化湾和闽江口春夏季鱼类区系特征,沈长春[10]研究了福建三沙湾鱼类群落组成特征及其多样性。
本文依据2009年冬季和2010年春、夏季在南日岛南部海域中部西侧海域调查资料,研究了调查海域内鱼类群落的划分及不同鱼类群落特征的变化。参考有关文献对这一海域水团及饵料分布的描述,分析其对不同适温性鱼类的影响而最终导致群落特征的影响变化。
1 材料与方法
1.1 调查海域和采样方法
分别于2009 年12 月, 2010 年4 月与8 月在南日岛南部海域(24°55′ —25°13′ N和119°11′—119°32′ E)进行了1个航次的渔业资源调查。采用的方法为单拖网,共设置12个测站(图1)。调查船为闽长渔4412#,网口7.5 m宽×3.5 m高,网目范围为2.5—12 cm,其中囊网网目3—6 cm,上部网衣12 cm,中部网衣8 cm。每网拖曳约0.5 h,平均拖速4.63 km/h。鱼类按照分类阶元逐级鉴定,最后确定其分类地位,鉴定到种,并进行分物种的尾数统计等。

图1 调查站位分布Fig.1 Location of sampling stations
1.2 资源密度的计算
本次鱼类资源密度估算采用扫海面积法[11],在渔获物统计结果基础上,计算各站位鱼类尾数密度,公式如下:
ρi=Ci/(ai×q)
式中,ρi为第i站的鱼类资源尾数密度(103尾/km2);Ci为第i站位单位时间渔获物中鱼类尾数(尾/h);ai为第i站位单位时间的扫海面积(km2/h),其值为网口水平宽度(km,本网具为0.0075 km)×拖曳距离(km),拖曳距离为拖速(km/h) 和实际拖曳时间(h)的乘积;q为网具捕获率(可捕系数= 1-逃逸率),根据本网具参数,尾数逃逸率取0.4—0.6[12]。
1.3 聚类分析
运用 Bray-Curtis[13]的相似性系数计算各季节不同站位之间鱼类群落的相似性,分析调查海域鱼类群落的群聚变化特征:

式中,SB为相似性系数,S为第m站位的种类数,第Xij和Xim分别为第i种类在j和m站位中的渔获尾数。由于不同群落的差异主要为关键种、优势种和常见种的差异,为减少稀有种对相似性分析的干扰,分析前首先剔除出现率低较低的种类,即将仅在1个占位出现的种类剔除,再以尾数密度为指标、站位为样本,进行等级聚类(CLUSTER)。为平衡优势种和非优势种在群落中的作用,计算前对原始数据矩阵进行开四次方根转换。
1.4 生态类群划分
依据聚类结果,参照田明诚等[14]对鱼类适温类型的分析方法,及《江苏鱼类志》[15]、《东海鱼类志》[16]等专著和文献[9],确定调查海域鱼类的生态属性。
1.5 多样性
多样性指数采用Shannon-Wiener指数[17]计算公式为:
式中,S为种类数;ni为第i种尾数;N为总尾数。
2 结果
2.1 鱼类群落组成及结构
春季鱼类群落聚类结果(图2)显示,在42.54%的相似性水平上可将调查海域鱼类划分为两个群落,其中Sp1、Sp2、Sp4、Sp5、Sp7为北部沿岸群落,Sp3、Sp6、Sp8、Sp9、Sp10、Sp11、Sp12为南部近海群落。两个群落在特有种上也有明显差别,如北部近海群落特有种为短吻舌鳎(Cynoglossusabbreviatus)凤鲚(Coiliamystus)等,南部近海群特有种有大头白姑鱼(Argyrosomusmacrocephalus)、海鳗(Muraenesoxcinereus)、小黄鱼(Larimichthyspolyactis)等。
夏季,在49.23%的相似性水平上可将调查海域鱼类划分为两个群落(图2),其中Su1、Su2、Su4、Su5、Su7、Su8、Su10、Su11为西部沿岸群落,Su3、Su6、Su9、Su12为东部近海群落。两个群落在特有种上也有明显差别,如西部沿岸群落特有种为多鳞鱚(Sillagosihama)、短吻舌鳎(Cynoglossusabbreviatus)等,东部近海群落群落特有种为鳞鳍叫姑鱼(Johniusdistinctus)、黄姑鱼(Nibeaalbiflora)等。
冬季,在54.21%的相似性水平上可将调查海域鱼类划分为两个群落(图2),其中Wi1、Wi2、Wi3、Wi4为北部沿岸群落,Wi5、Wi6、Wi7、Wi8、Wi9、Wi10、Wi11和Wi12为南部近海群落。两个群落在特有种上也有明显差别,如北部沿岸群落特有种为横纹东方鲀(Takifuguoblongus)、莱氏舌鳎(Cynoglossuslachner)等;南部近海群落特有种为赤魟(Dasyatisakajei)、棘头梅童鱼(Collichthyslucidus)等。

图2 各季节鱼类群落的Bray-Curtis聚类结果Fig.2 Bray-Curtis clustering and nonlinear multidimensional scaling of fish communities
2.2 不同群落生态类群组成的比较
春季,南部近海群落种类数为32种,北部群落有27种。南部近海群落中暖水种与暖温种种类数均高于北部沿岸群落,但暖温种差别不大(表1)。

表1 各群落不同生态类群的种类数(种)
夏季,西部沿岸群落种类数(55种)远高于东部近海群落(27种)(图3)。西部沿岸群落中沿岸种种类数达27种,东部近海群落仅有10种。近海种上西部沿岸群落(24种)也高于东部近海群落(15种)(表1)。
冬季,南部近海群落种类数(32种)高于北部沿岸群落(25种) (图3)。其中南部近海群落中暖水种种类数(24种)高于北部沿岸群落(17种),而两个群落暖温种的种类数相同(表1)。

图3 各季节鱼类种类数平面分布Fig.3 Seasonal distribution of fish species
2.3 不同鱼类群落和生态类群鱼类数量的分析
春季,南部近海群落暖水种尾数密度(23.83×103尾/km2)高于北部沿岸群落(6.12×103尾/km2),北部沿岸群落暖温种尾数密度(1.35×103尾/km2)高于南部近海群落(0.51×103尾/km2) (表2,图4)。
夏季,西部沿岸群落暖水种、暖温种尾数密度(327.07×103尾/km2和2.16×103尾/km2)均高于东部近海群落(106.37×103尾/km2和0.10×103尾/km2)。沿岸种与近海种尾数密度也为西部沿岸群落(247.20×103尾/km2和80.46×103尾/km2)高于东部近海群落(78.38×103尾/km2和27.90×103尾/km2)。
冬季,南部近海群落暖水种、暖温种尾数密度(24.27×103尾/km2和4.57×103尾/km2)均高于北部沿岸群落(8.13×103尾/km2和1.83×103尾/km2),暖水种与暖温种在各站位均有出现(表2,图4)。
2.4 不同鱼类群落多样性指数分析
春季,北部沿岸群落多样度H′值(2.71)高于南部近海群落(1.97)。夏季,西部沿岸群落(1.97)H′值高于东部近海群落(1.62)。冬季,北部沿岸群落(2.45)H′值高于南部近海群落(2.33)。
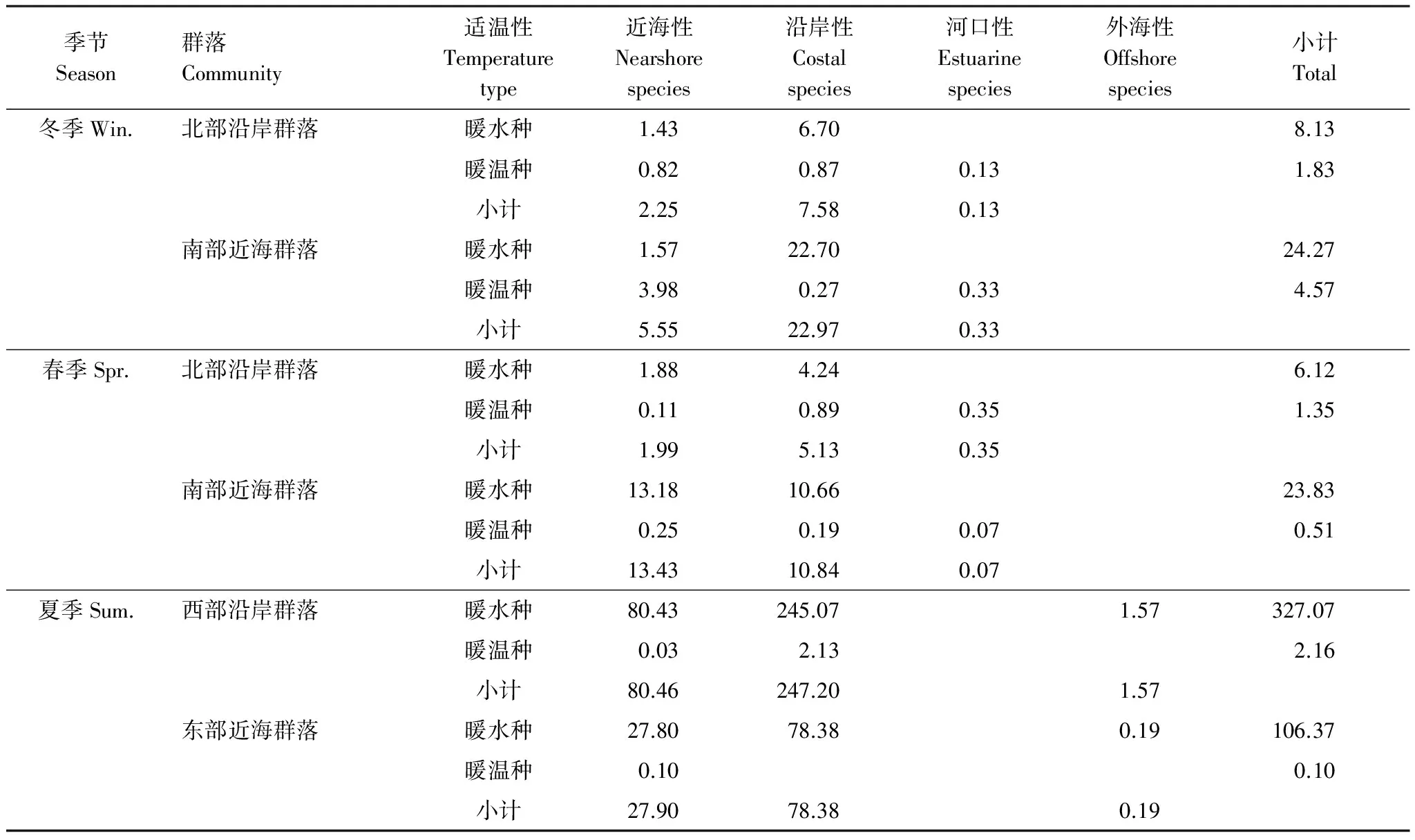
表2 各群落不同生态类群尾数密度(103尾/km2)
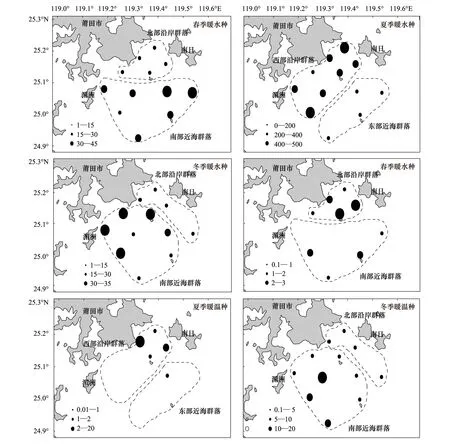
图4 各生态类群尾数密度平面分布Fig.4 Distribution of individual density in each ecological groups

图5 各鱼类多样度平面分布4Fig.5 Distribution of diversity of fish
3 讨论
3.1 水团变化对不同群落种类数的影响
鱼类种类组成是在不同鱼类种群相互联系及其所处环境因子的长期综合影响中逐渐形成的,而在环境因子中,水温、盐度对鱼类群落的影响较大[7]。由于水团运动是影响水体温、盐环境变化的主要原因,而各季节水团环境差异,造成不同鱼类群落间种类数出现较大差异。一般而言,冬、春季北部沿岸群落均低于南部近海群落,夏季西部沿岸群落低于东部近海群落(表1)。这种现象与水团影响及鱼类洄游有关。
冬季调查时间为12月上旬,此时浙闽沿岸水势力较弱,进入台湾海峡中部西侧海域后沿着岸线向南推进[18],因而调查海域离岸侧还受到变性水团的影响。春季调查海域除受浙闽沿岸水控制外,南部海域则受到包含海峡暖水和浙闽沿岸水的海峡混合水的影响[19]。由于冷对冬、春季调查海域北侧造成的影响,大部分洄游性鱼类向南或外海洄游越冬,造成调查海域北侧鱼类种类数下降。而混合水团对南侧的影响,使得调查海域南侧鱼类种类数仍保持一定水平(表1)。
夏季,东部海域受海峡暖水控制,西部近岸海域除了受到海峡暖水影响,还受到径流带来丰富的营养盐的影响[19]。因而调查海域近岸侧在受海峡暖水和地表径流的双重影响下,饵料丰度高于离岸侧[20],适宜鱼类集聚索饵、育肥。因而,西部近岸群落种类数要高于东部外海群落种类数。
综上所述,水团对不同群落种类数特征的形成有重要影响。
3.2 水团变化对不同生态类群数量密度的影响
南日岛南部海域属于典型的亚热带海域,鱼类群落整体上表现为暖水性,因而各群落均表现为暖水种数量密度高于暖温种(表2,图4)。而在不同季节的环境背景下,也造成不同群落间暖水种、暖温种的数量密度的差异。
冬季,相对各季节,浙闽沿岸水势力最强,而海峡暖水最弱[21],因而南部群落所在的海域环境相比其他季节具有更加明显暖温带性。有利于暖温种群落的发展,形成冬季南部近海群落中暖温种数量密度高于北部沿岸群落暖温种数量密度(表2,图4)的特征,与其他季节明显不同。尽管如此,冬季,调查水域无论南北群落均显示出暖水种数量密度高于暖温种数量,这是南日岛南部海域具有典型的亚热带海域鱼类的群落特征决定的。
春季,影响调查海域的浙闽沿岸水势力减弱,浙闽沿岸水势力对南部海域环境影响力降低,海峡暖水在逐步增强的过程中,对南部海域的影响强于浙闽沿岸水[6]。暖温种种群部分退出南部海域,而在北部海域还有一定的数量,形成南部海域近海群落中暖温种数量密度低于北部沿岸群落暖温种数量密度(表2,图4),这样与冬季不同的特征。暖水种尾数密度依然保持为南部近海群落高于北部沿岸群落,且春季南、北部两个群落中暖水种尾数密度的差值高于冬季(表2,图4),这进一步证实春季海峡暖水对南部海域的影响较冬季有所增强。
夏季海峡暖水基本上控制了调查海域,因而夏季该域海鱼类尾数密度均较高,东、西部两个鱼类种群几乎均由暖水种组成(表2,图4)。西部沿岸群落在受海峡暖水的同时,还因地表径流量增大,使得水体盐度低于东部海区,以及径流带来丰富的饵料[22],因而西部沿岸群落中沿岸种数量密度极高(表2,图4)。而东部海域受海峡暖水及上升流带来的高盐水的共同影响[[23],东部海域中近海种的比例高于西部海域(表2)。但夏季东、西部两个群落的鱼类数量密度均高于冬、春季其他群落(表2,图4),这种随着夏季适宜范围内温升,鱼类数量有增加的趋势与国内外学者的观点较为一致[24- 26]。
以上分析可见,冷、暖水团的交替变化,影响了各季节不同群落中不同生态类群数量密度的高低变化。
3.3 水团变化对不同群落鱼类多样性影响
影响多样度(H′)数值高低,一是种类数大小,二是同一站位不同种类尾数之间的均匀性[27]。2.4显示多样度(H′)为冬、春季北部群落高于南部,夏季西部群落高于东部。由于水团变化直接影响和限制了鱼类的行为和栖息环境的变化[28],从而对鱼类生物多样性造成影响。可以认为调查海域鱼类多样度的这种分布趋势与交替影响该海域的水团有关。
冬季,影响调查海域北侧的浙闽沿岸水逐渐增强,而南部还受逐渐减弱的海峡暖水与浙闽沿岸水的混合水团影响[21]。这种南、北海域分受不同性质水团影响的情况,使得鱼类多趋向于温暖的南部海域或外海越冬或产卵。因而在种类数、数量密度上均表现为受海峡暖水影响的南部高于受浙闽沿岸水的北部群落(表1,表2)。这使得南、北侧不同鱼类群落的组成结构发生了变化。且暖水团控制的海域已形成个别种类的优势种群,导致鱼类群落多样度降低。故冬、春季北部沿岸群落多样度均高于南部近海群落(图5)。
夏季,与冬、春季优势种群所在海域多样度较低不同,夏季优势种群分布在西部海域,而西部沿岸群落多样度却依然高于东部近海群落,尤以1、2、4和5号站位最高(图5)。这是因为夏及调查海域除受海峡暖水影响之外,地表径流的增大成为影响沿岸海域的重要因素之一。多样度较高的4个站位位于西部海域的北部,这里是兴化湾湾口所在,受海湾内外水体交换影响较为显著,水文环境多变[29- 30]。受这些因素共同影响,夏季西部海域鱼类种类数与数量密度均较高(图3,图4),虽然出现了优势种群,但常见种种类数较多,种间尾数密度均匀,使得西部海域多样度大于东部海域。
因此,水环境的多样性被认为是鱼类生物多样性差异的主要外部因素。而环境适应的多样性作为鱼类群落形成的内在因素,其作用机理也与水团相联系。
3.4 鱼类适温性多样性与群落形成的关系
由于受冷暖水团交替变化影响,该海域不同适温性鱼类的分布具有随水团变化的季节性和区域性。不同适温性鱼类随水团及饵料变化而呈现的分布差异,是不同鱼类群落生物多样性形成的内在因素。
冬季,低温、低盐的浙闽沿岸水深入到受海峡混合水影响的南部,这里平均温度较高,因而适宜暖水种的生存[14,31]。由于平均水温也在暖温种的适宜范围内,暖温种也有一定的数量。不过,暖温种的分布对群落形成的影响不及暖水种的作用显著,因而,冬季群落的划分,与暖水种的分布趋势较为一致(图4)。
春季,浙闽沿岸水暖温种高值区集中分布在调查海域北部(图3,图5)。而由于海峡暖水的势力增强,使得南部海域具有更加明显的亚热带的性质,暖水种在这一海域大量出现,其尾数密度分布高值区亦位于此(4,5)。暖水种与暖温种分高值区的南北对应,与春季群落划分的南北格局相对应(图2)。说明春季群落的划分,受鱼类环境适应性的影响明显。
夏季,受海峡暖水控制,暖水种在12个调查站位均大量出现,尤以调查海域的西部最为显著(图4)。高温、高盐的海峡暖水与沿岸径流种丰富的营养盐使得暖水种及暖温种均大量出现于西部近岸,这也与夏季近岸海域鱼类饵料丰度较高有关[20,22]。由于东部海域水温较高,平均水温超过了暖温种的适温上限[26],暖温种的出现受到限制。仅6号站有少量暖温种出现。因此可将暖水种与暖温种大量出现的西部鱼群和基本由暖水种构成的东部鱼群划分为东、西两个群落,即为西部沿岸群落和东部近海群落。
综上,冬、春季与夏季两种群落格局划分方式主要体现出南日岛南部海域鱼类群落特征在种类数、数量密度、多样度及适温性方面的区域性差异,而造成这种区域性差异的外在原因主要为南日岛南部海域季节性交替变化的水团及其带来的水文环境及饵料分布上的差异。
致谢:本文撰写过程得到沈晓民先生的大力指导和帮助,特此致谢。
[1] 陈新军. 渔业资源与渔场学(第二版). 北京: 海洋出版社, 2014: 472- 472.
[2] Lazzari M A, Sherman S, Brown C S, King J, Joule B J, Chenoweth S B, Langton R W. Seasonal and annual variations in abundance and species composition of two nearshore fish communities in Maine. Estuaries, 1999, 22(3): 636- 647.
[3] Ayvazian S G, Hyndes G A. Surf-zone fish assemblages in south-west Australia: do adjacent nearshore habitats and the warm Leeuwin Current influence the characteristics of the fish fauna?. Marine Biology, 1995, 122(4): 527- 536.
[4] Aschan M, Fossheim M, Greenacre M, Primicerio. Change in fish community structure in the Barents Sea. PLoS One, 2013, 8(4): e62748.
[5] 陈永俊, 张静, 宋普庆, 张然, 李渊, 钟指挥, 林龙山. 台湾海峡鱼类组成及其生态区系. 生物多样性, 2014, 22(4): 525- 531.
[6] 翁学传, 张启龙, 颜廷壮, 王从敏, 梁红星. 台湾海峡中、北部海域春、夏季水团分析. 海洋与湖沼, 1992, 23(3): 235-244.
[7] 宋普庆, 张静, 林龙山, 许章程, 朱小明. 台湾海峡游泳动物种类组成及其多样性. 生物多样性, 2012, 20(1): 32- 40.
[8] 林龙山, 郑元甲, 马春艳. 台湾海峡夏秋季游泳动物资源分布及群落结构. 应用生态学报, 2005, 16(10): 1948-1951.
[9] 徐兆礼. 兴化湾和闽江口海域春夏季鱼类区系特征. 生物多样性, 2011, 19(1): 79- 84.
[10] 沈长春. 福建三沙湾鱼类群落组成特征及其多样性. 海洋渔业, 2011, 33(3): 258- 264.
[11] 詹秉义. 渔业资源评估. 北京: 中国农业出版社, 1995: 18- 46, 124- 130.
[12] 杨吝, 张旭丰, 谭永光, 张鹏. 南海区拖网网囊最小网目尺寸选择性研究. 中国水产科学, 2003, 10(4): 325- 332.
[13] Bray J R, Curtis J T. An ordination of the upland forest communities of Southern Wisconsin. Ecological Monographs, 1957, 27(4): 325- 349.
[14] 田明诚, 孙宝龄, 杨纪明. 渤海鱼类区系分析. 海洋科学集刊, 1993, (34): 157-167.
[15] 福建鱼类志编写组. 福建鱼类志. 福州: 福建科学技术出版, 1984.
[16] 朱元鼎, 张春霖, 成庆泰. 东海鱼类志. 北京: 科学出版社, 1963.
[17] Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana: University of Illinois Press, 1975.
[18] 潘爱军, 万小芳, 郭小钢, 靖春生. 平潭岛外海浙闽沿岸流对2006年冬季风松弛的响应特征与机理. 中国科学: 地球科学, 2012, 42(9): 1317- 1328.
[19] 肖晖, 郭小钢, 吴日升. 台湾海峡水文特征研究概述. 台湾海峡, 2002, 21(1): 126-138.
[20] 田丰歌, 徐兆礼. 福建中部近海浮游动物数量分布与水团变化的关系. 生态学报, 2012, 32(4): 1097- 1104.
[21] 张彩云, 商少凌, 陈德文, 商少平. 冬季浙闽沿岸水分布的短期变动与风的关系初探. 遥感学报, 2005, 9(4): 452- 458.
[22] 阙江龙, 徐兆礼, 陈佳杰. 台湾海峡中部近海虾类数量和优势种分布特征. 中国水产科学, 2014, 21(6): 1211-1219.
[23] 黄荣祥. 台湾海峡中、北部海域的上升流现象. 海洋湖沼通报, 1989, (4): 8-12.
[24] Senta T, Kinoshita L. Larval and juvenile fishes occurring in surf zones of western Japan. Transactions of the American Fisheries Society, 1985, 114(4): 609- 618.
[25] Modde T, Ross S T. Seasonality of fishes occupying a surf zone habitat in the Northern Gulf of Mexico. Fishery Bulletin, 1981, 78(4): 911- 922.
[26] 吴美琴, 钟俊生, 葛珂珂, 练青平, 蒋日进. 长江口沿岸碎波带仔稚鱼分布的季节性变动. 渔业科学进展, 2010, 31(1): 1- 7.
[27] 徐兆礼, 沈盎绿. 兴化湾海域鱼类多样性的时空变化. 中国水产科学, 2011, 18(2): 416- 426.
[28] 龙华. 温度对鱼类生存的影响. 中山大学学报: 自然科学版, 2005, 44(S1): 254- 257.
[29] 王珊珊, 曹志敏, 蓝东兆, 许江, 李桂海, 翟滨. 福建省兴化湾湾口海域营养盐的分布状况及其影响因素. 环境化学, 2006, 25(2): 234- 235.
[30] 陈水土. 福建省兴化湾海域营养盐分布变化特征及其影响因素. 海洋通报, 2000, 19(5): 26- 31.
[31] 陈永俊. 台湾海峡鱼类物种多样性及生态区系的研究. 厦门: 国家海洋局第三海洋研究所, 2014.
Structure and diversity of fish communities in the waters south of Nanri Island
GUAN Wei1,2, XU Zhaoli1,*, CHEN Jiajie1
1EastChinaSeaFisheriesResearchInstitute,ChineseAcademyofFisherySciences,KeyandOpenLaboratoryofMarineandEstuarineFisheries,MinistryofAgriculture,Shanghai200090,China2CollegeofMarineSciences,ShanghaiOceanUniversity,Shanghai201306,China
To explore the characteristics and diversity of fish communities in the waters south of the Nanri Island, three comprehensive surveys of fishery resources were conducted in the winter of 2009 and the spring and summer of 2010. Bray-Curtis cluster was used to classify the fish communities in the surveyed area, and the number of species, density, ecological groups, and diversity of fish species were analyzed on the basis of the division of fish communities. The effects of the changes in water masses on fish communities′ classification and diversity were discussed in detail. The results indicate that in spring, the fish communities comprise two different groups: the northern coastal community and the southern nearshore community. The number of species, density, and the individual diversity index (H′) of the northern coastal community were 27 species, 7.47 × 103individuals/km2, and 2.71, respectively, and the corresponding values of the southern nearshore community were 32 species, 24.34 × 103individuals/km2,and 1.97, respectively. The number of species and density of warm water species of the southern offshore community were higher than the corresponding values of the northern offshore community. In contrast, the warm temperate species exhibited the opposite scenario-0. In summer,the number of species, density, and H' of the western coastal community were 55 species, 329.24 × 103individuals/km2, and 1.97, respectively, while the respective values of the eastern nearshore community were 27 species, 106.47 × 103individuals/km2, and 1.62, respectively. The individual density of both coastal species and nearshore species of the western coastal communities were higher than those of the eastern nearshore community. In winter, the fish communities were divided into northern coastal community and southern nearshore community. The number of species, density, andH′ in the northern coastal community were 25, 9.96 × 103individuals/km2, and 2.71, whereas the corresponding values in the southern nearshore community were 32 species, 28.85 × 103individuals/km2, and 1.97, respectively. The individual density of warm water species and warm temperate species in the southern coastal community were higher than those in the northern nearshore community. In winter and spring, the number of fish species, individual density, and diversity, of the warm water species in the area controlled by the Taiwan Strait warm waters were higher than those of the species dominated in the Fujian-Zhejiang coastal waters. In summer, with the influence of the Taiwan Strait warm water and overland runoff, the corresponding values of the fish community in the western region of the study area were higher than those in the eastern region. We conclude that the demarcation and characteristics of the fish communities were closely related to the seasonal changes of water masses.
fish; communities; diversity; pattern; water mass; waters south of Nanri Island
2016- 01- 24; 网络出版日期:2016- 12- 19
10.5846/stxb201601240167
*通讯作者Corresponding author.E-mail: xiaomin1957@126.com
管伟,徐兆礼,陈佳杰.福建南日岛南部水域鱼类群落结构及多样性.生态学报,2017,37(9):3172- 3181.
Guan W, Xu Z L, Chen J J.Structure and diversity of fish communities in the waters south of Nanri Island.Acta Ecologica Sinica,2017,37(9):3172- 3181.
