小兴安岭天然森林沼泽湿地生态系统碳源/汇
2017-06-22牟长城
李 娜,牟长城,王 彪,张 妍,马 莉
东北林业大学生态研究中心, 哈尔滨 150040
小兴安岭天然森林沼泽湿地生态系统碳源/汇
李 娜,牟长城*,王 彪,张 妍,马 莉
东北林业大学生态研究中心, 哈尔滨 150040
采用静态箱-气相色谱法与相对生长方程法,同步测定小兴安岭7种天然沼泽湿地(草丛沼泽-C、灌丛沼泽-G、毛赤杨沼泽-M、白桦沼泽-B、落叶松苔草沼泽-L-T、落叶松藓类沼泽-L-X、落叶松泥炭藓沼泽-L-N)的土壤呼吸(CO2、CH4)净碳排放量、植被年净固碳量,并依据生态系统净碳收支平衡,揭示温带天然沼泽湿地的碳源/汇作用规律。结果表明:①7种天然沼泽CH4年通量(0.006—7.756 mg m-2h-1)呈M(高于其他类型1.0—1291.7倍,P<0.05)>C、G、B(高于针叶林沼泽17.7—649.0倍,P>0.05)>针叶林沼泽变化趋势,其季节动态存在3种类型(C、G单峰型、M、B多峰型及针叶林沼泽排放与吸收交替型);②其CO2年通量(157.40—231.06 mg m-2h-1)呈G(高于森林沼泽28.7%—46.8%,P<0.05)>C(高于森林沼泽7.4%—22.5%,P>0.05)>森林沼泽的变化趋势,其季节动态存在2种类型(C、G、L-X和L-N双峰型和M、B、L-T单峰型);③C、G、M、B、L-N CH4排放仅受0—40 cm不同土壤层温度所控制;7种天然沼泽土壤CO2排放均受气温及0—40 cm不同土壤层温度所控制,但B、L-X、L-N受温度与水位综合控制;④其植被年净固碳量((2.05±0.09)—(6.75±0.27)tC hm-2a-1)呈C(高于其他类型65.4%—229.3%,P<0.05)>G、B、L-T、L-X、L-N(高于M 80.0%—99.0%,P<0.05)>M变化趋势;⑤7种天然沼泽的碳源/汇(-2.32—2.09 tC hm-2a-1)作用不同,C、B和L-N为碳吸收汇(C强汇、B和L-N弱汇),M、G、L-T和L-X则为碳排放源(M、G强源、L-T和L-X弱源)。因此,温带小兴安岭草丛沼泽为碳强汇、灌丛沼泽为碳强源、森林沼泽基本维持碳平衡(除M外)。
小兴安岭;天然沼泽湿地;温室气体排放;年净固碳量;碳源/汇
湿地因其具有巨大土壤碳库、高甲烷排放及其在泥炭形成、沉积物堆积与植物生物量积累等方面的固碳潜力,使其成为全球碳循环的重要组成部分[1-2]。天然湿地能够吸收固定大气中的CO2,被认为是大气温室气体的吸收汇[3- 7],同时天然湿地每年向大气中排放大约1.45×1011kg CH4-C,占全球人为和自然甲烷排放总量的25%[8],故湿地又被认为是大气温室气体的排放源[8- 11]。因此,目前有关天然湿地究竟是碳源还是碳汇仍存有争议。
现有研究结果表明:天然湿地甲烷排放通量随气候区或湿地类型变异性较大,如北美泥炭藓沼泽CH4通量在亚热带、温带、寒温带分布在2.79—25.68 mg CH4-C m-2h-1[12-13],变化幅度达9.2倍;森林湿地CH4通量在热带、亚热带、温带分布在-0.029—92mg CH4-C m-2h-1[14- 18],变化幅度相差几个数量级,总体上呈现随纬度升高而递减趋势。又如北欧同一气候区各类型湿地CH4通量分布在-1.3—1583mg CH4-Cm-2h-1[19],变化幅度也相差几个数量级。这也是导致全球湿地甲烷排放总量估算不确定性的原因之一。且湿地甲烷排放受温度、水位、pH值[8,20-21]、底物类型与质量[22-23]和特定微生物[8,24]等因素影响。特别是水位影响更加强烈,如泥炭地在干旱季节水位低时能够吸收大气甲烷,而被水饱和后则向大气排放甲烷[25-26]。其原因在于水位决定湿地不同深度土壤层有氧和厌氧条件,进而控制着甲烷产生(严格厌氧过程)和氧化过程(需氧过程)[27]。故平均水位是湿地甲烷排放的最佳指示者,水位低于泥炭表面12 cm,甲烷通量较高[28];水位低于泥炭表面25 cm,甲烷氧化过程增强,泥炭地则由甲烷排放源转化为甲烷吸收汇[29]。但也有学者发现湿地CO2排放随着温度的升高和水位的降低而增加[30- 32],且温带森林湿地土壤温室气体排放以CO2排放占绝对优势地位[33- 35]。因此,目前仅根据CH4排放判断湿地碳源是不全面的,尚需同步观测CH4和CO2排放方可得到湿地异养呼吸总碳排放源强。
天然湿地具有丰厚的土壤泥炭层,伴随着泥炭层形成、沉积物堆积和植物生物量累积等过程,能够吸收固定大气中的CO2,如泥炭地仅占全球陆地表面3%,却含有近1/3全球土壤碳库(16%—33%)[1-2],且具有高植被生产力、高水位、低分解速率的湿地能够发挥更强的碳汇作用[36]。目前国际上有关湿地固碳研究集中在北美和欧洲,且多集中在海岸湿地、河口湿地、平原草丛湿地及泥炭地的沉积物、土壤和植物群落的固碳方面[3- 7],国内关于东北山区森林湿地碳循环方面的研究也主要集中在温室气体排放[18,33- 35,37- 47],以及碳储量及固碳量[48- 52]等方面,而真正从生态系统尺度上探讨其碳源/汇作用方面的研究报道甚少。并得到平原湿地固碳能力(0.3 kg C m-2a-1)[6]约为农田和草地(0.16 kg C m-2a-1和0.22kg C m-2a-1)的2倍[53];而泥炭地固碳能力(北方0.015—0.026、温带0.01—0.046、北美0.029kg C m-2a-1)[1,54]似乎仅为农田和草地的1/10—1/5,但远高于北方森林固碳能力(0.003—0.01 kg C m-2a-1)[55]。其固碳机制为湿地土壤一般多处于充分水饱和状态或经常位于水面以下,形成厌氧土壤环境,有机物分解过程缓慢,能够以泥炭形式存储CO2,但当湿地形成未饱和土壤环境或水位显著低于泥炭表面时,CO2排放较水淹状态明显增大[31-32],而CH4排放则减少或转化为吸收汇[56-57]。故有学者认为这两个因素结合决定着湿地系统对温室气体的总贡献[5]。因此,有必要依据生态系统净碳收支平衡探讨湿地碳源/汇作用。
本文以小兴安岭沿湿地过渡带环境梯度依次分布的7种典型天然沼泽湿地(草丛沼泽、灌丛沼泽、5种森林沼泽)为对象,采用静态箱-气象色谱法、碳/氮分析仪测定法与相对生长方程法,野外原位同步测定7种天然沼泽湿地的土壤呼吸(CO2、CH4)年净碳排放量、植被净初级生产力与年净固碳量,并依据生态系统净碳收支平衡,揭示我国东北温带天然沼 泽湿地碳源/汇作用规律,并结合环境因子探讨其形成机制,以期为我国东北林区湿地资源经营管理提供科学依据。
1 研究区域概况与方法
1.1 研究区域概况
研究地点位于小兴安岭中段,黑龙江省伊春市友好林业局永青林场(48°03′53″—48°17′11″N,128°30′36″—128°45′00″E)。平均海拔260—500 m。受海洋气团和西伯利亚寒流影响,气温波动较大,属温带大陆湿润季风气候。年平均气温约0.4℃,年积温在2000℃到2500℃之间。年平均降水量630 mm,全年有两个降水高峰期,一是冬季的降雪,二是每年7—8月间的降雨,高峰期降水占全年降水量的70%,无霜期约为110 d,最早霜期在9月上旬,最晚霜期在次年的5月中旬结束,霜期长达6个月。地带性植被为温带红松阔叶混交林。地带性土壤是暗棕壤,占所有土壤类型的71%,其他土壤有草甸土、沼泽土和泥炭土,草甸土和沼泽土占27%,泥炭土占2%。主要湿地类型有草丛沼泽、灌丛沼泽、森林沼泽、藓类沼泽和浅水沼泽等多种类型,但以森林沼泽为其主体[38-39,58]。
1.2 样地设置
于2007年在小兴安岭友好林业局永青林场选择了沿湿地过渡带水分环境梯度依次分布的7种天然沼泽湿地(草丛沼泽-C、灌丛沼泽-G、毛赤杨沼泽-M、白桦沼泽-B、落叶松苔草沼泽-L-T、落叶松藓类沼泽-L-X、落叶松泥炭藓沼泽L-N)作为研究对象。2014—2015年野外观测工作是在选定的各天然沼泽湿地类型中各设置3块20 m×30 m样地,共设置21块样地,并在每个类型重复设置3个静态箱,共计设置21个静态箱,进行土壤CO2和CH4同步气体取样观测。
1.3 研究方法
1.3.1 植物净初级生产力与年净固碳量的测定
本研究将全年按照如下时间段划分:春季为3月下旬—6月中旬,夏季为6月下旬—9月中旬,秋季为9月下旬—12月中旬,冬季为12月下旬—次年3月中旬;生长季为5月上旬—9月下旬,非生长季为10月上旬—次年4月下旬。
乔木层净初级生产力测定:于2014年春季生长开始期(5月上旬)和秋季生长停止期(9月下旬),对各类型森林沼泽样地中的林木胸径生长量进行了测定,并利用现有本地区不同树种胸径与生物量的回归方程(表1)[52],计算生长初期和生长结束期的乔木层生物量,取两者的差值估算乔木层的净初级生产力。
灌木层和草本层净初级生产力测定:在每个标准地(20 m×30 m)中心与四角设置5个2 m×2 m灌木样方,随机设置10个1 m×1 m的草本样方,采用收获法,获取灌木层与草本层地上与地下的生物量鲜重,并对其进行取样在70℃下烘干至恒重,计算两者含水率,得到灌木层与草本层的生物量干重;灌木层净初级生产力为其生物量除以平均年龄(5 a),草本层净初级生产力为其地上部分与地下部分当年生物量的加和(地上部分全部视为当年生,地下部分选取靠近茎干、颜色较浅部分作为当年生新根)。
植被有机碳浓度的测定:利用碳/氮分析仪Multi N/C 3100 和 HT 1300 Solids Module (Analytik Jena AG,Germany) 通过1300℃干烧法测定乔木层(干、根、枝、叶)、灌木层(根、茎、叶)、草本层(地上茎叶、地下根系)有机碳浓度。
植被年净固碳量的测定:乔木层、灌木层、草本层的年净固碳量可通过各自的年净初级生产力与其相应的碳浓度的乘积获得,植被层的年净固碳量将三者加和而得到[51]。
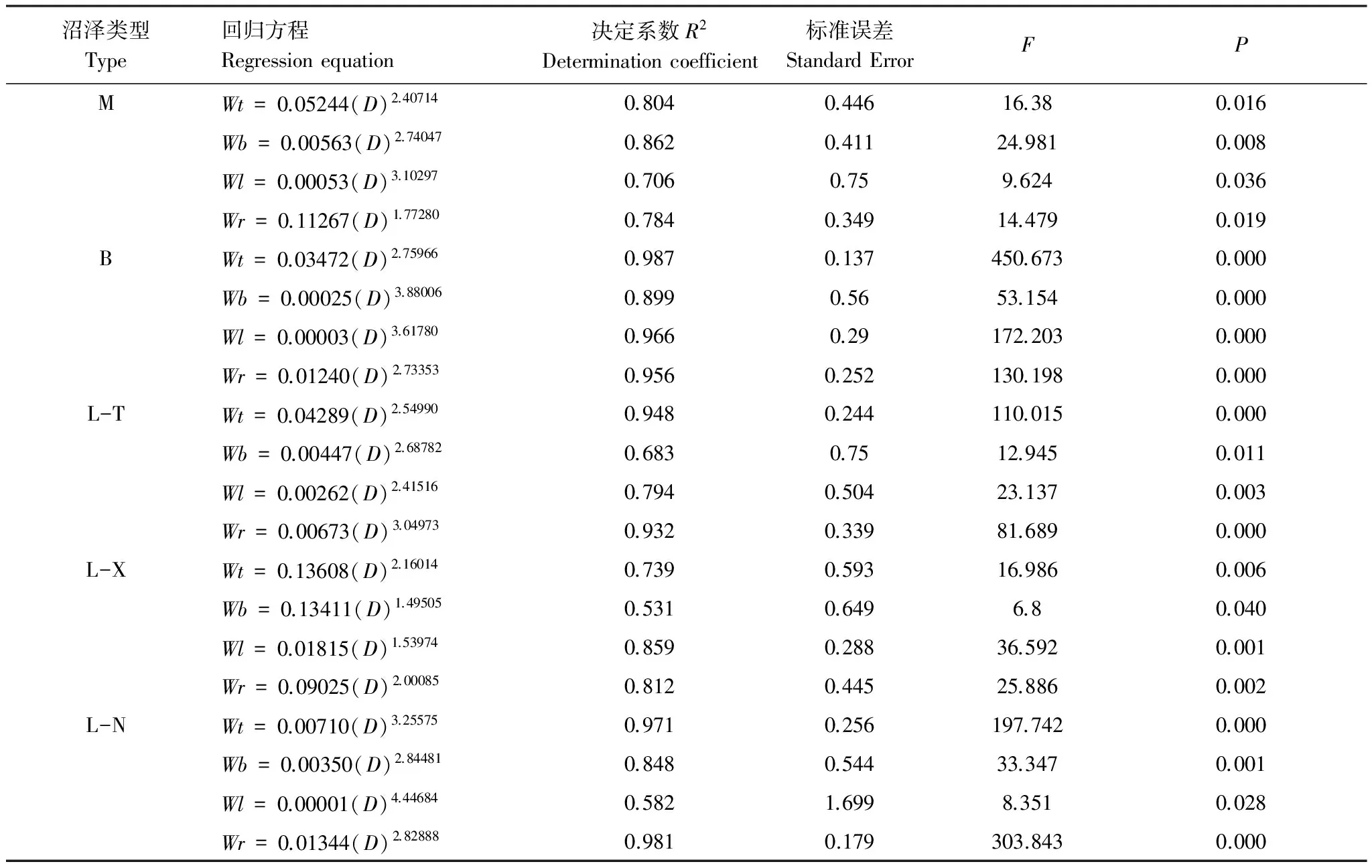
表1 乔木各器官生物量回归方程
Wt、Wb、Wl、Wr分别为乔木干、枝、叶、根的生物量方程;D为乔木胸径; M:毛赤杨沼泽Alnussibirica-C.schmidtiiswamp;B:白桦沼泽B.platyphylla-C.schmidtiiswamp; L-T:落叶松-苔草沼泽Larixgmelinii-C.schmidtiiswamp;L-X:落叶松、藓类沼泽L.gmelinii-moss swamp;L-N:落叶松-泥炭藓沼泽L.gmelinii-Sphagnumspp. swamp
1.3.2 气体采集与分析
气体采集采用静态暗箱-气象色谱法。暗箱由不锈钢顶箱(规格为50 cm×50 cm×50 cm)和不锈钢底座(规格为50 cm×50 cm×10 cm)组成。采样前,把底座插入采样点土中10 cm,在安装完毕后底座放在样地不动直至全年取样结束,以保证对底座内部的土壤干扰最小。采样时,先将底座上部四周的凹槽注水密封,再安放顶箱(每次安放顶箱前需去除观底座内植物及其根系,以排除植物自养呼吸碳排放)。顶箱内部安有直径10 cm的小风扇,取样时保持风扇转动,避免箱内出现气体浓度差,风扇用12V蓄电池供电。在箱顶部开设2个直径1 cm的小孔,其一作为取样口,内置的橡胶塞;其二用于带橡胶塞的数字温度计探头通过。风扇电源线也穿过箱顶部,后用硅胶将小孔密封。顶箱外粘贴泡沫保温材料,以减少箱内温度波动。
取样时间为2014年5月8日—2015年4月28日。每隔10 d观测1次,每次取样在8:00—11:00完成,每个静态箱重复取4次(分别在静态箱封闭后0、10、20、30 min进行)。用60 mL聚氯乙烯医用注射器经三通阀连接铁针头通过箱顶部橡胶塞取样。气体样品用注射器取出后转移进100 mL气体采集袋保存并带回实验室,在一周内利用HP5890Ⅱ气象色谱仪完成浓度分析工作[59-60]。最后计算各沼泽湿地类型的土壤异养呼吸年碳排放量。
温室气体通量计算公式[61]:

式中,F为气体通量(mg m-2h-1),Po、Vo、To分别为标准状态下的标准大气压、气体摩尔体积和绝对温度;dc/dt为采样时气体浓度随时间变化的直线斜率;M为被测气体的摩尔质量;P、T为采样点的实际大气压和温度;H为采样箱高度。F为正值时表示排放,负值表示吸收[39]。根据CH4和CO2中碳元素所占分子式比例分别折算其含碳量,继而推算系统碳排放情况。
1.3.3 环境因子测定
气体采样同时,测定气温、箱温、0、5、10、15、20、30、40 cm土温以及观测点水位。使JM便携式数字温度计和相应长度的探针来进行温度的测定。沼泽水位通过在采集箱附近挖井,用钢卷尺进行测量。
1.3.4 碳源/汇测定
为揭示小兴安岭天然沼泽湿地生态系统的碳源/汇问题,将依据生态系统净碳收支平衡(年净固碳量与土壤异养呼吸碳排放差值)对其加以评价。
1.4 数据处理
采用SPSS 19.0和Microsoft Office Excel 2010软件进行数据分析。采用单因素方差分析(one-way ANOVA),采用最小显著差异法(LSD)分析不同数据组建的差异性,显著性水平设置为α=0.05。采用Pearson相关系数评价不同因子间的相关关系,相关性水平设置为α=0.05。
2 结果与分析
2.1 天然森林沼泽湿地CH4与CO2排放通量及其季节动态趋势
2.1.1 天然森林沼泽湿地CH4排放通量及其季节动态
由图1可以得到,小兴安岭7种天然沼泽CH4通量季节动态趋势有所不同。其中,C、G、M和B的CH4通量自早春开始缓慢升高,进入夏季迅速波动升高,并形成1—4个排放峰值(依次为13.6、26.0、30.25—39.80、11.06—14.75 mg m-2h-1),夏末秋初开始波动下降,直至整个冬季维持在较低排放水平;而3种针叶林沼泽(L-T、L-X和L-N)CH4通量在全年观测期间均呈现出排放与吸收交替出现的季节变化趋势。因此,小兴安岭7种天然沼泽类型中的草丛沼泽与灌丛沼泽CH4通量呈单峰型、2种阔叶林沼泽呈多峰型及3种针叶林沼泽呈排放与吸收交替型的季节变化趋势。
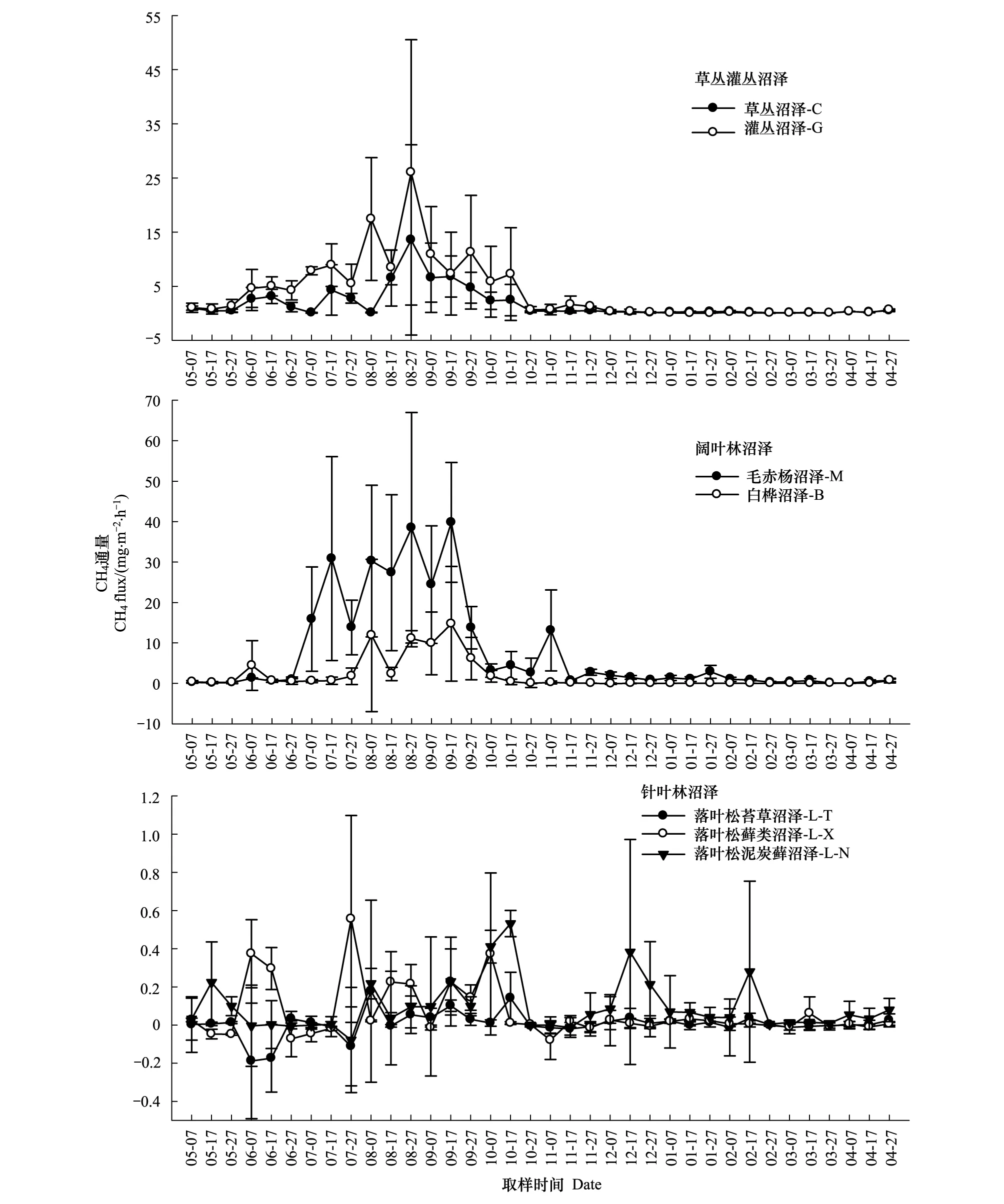
图1 小兴安岭7种天然沼泽类型CH4排放通量的季节动态Fig.1 Seasonal dynamics of methane emission fluxes from seven kinds of typical natural forested wetlands in Xiaoxing′anling of China
由表2可以得到,7种天然沼泽CH4年通量及季节通量也有所不同。其年通量分布在0.006—7.756 mg m-2h-1之间,以M最高(显著高于其他6种沼泽1.0—1291.7倍,P<0.05),C、G和B居中(三者高于3种针叶林沼泽17.7—649.0倍,P>0.05),3种针叶林沼泽最低且相近。因此,7种天然沼泽CH4年通量呈现出毛赤杨沼泽>草丛沼泽、灌丛沼泽和白桦沼泽>3种针叶林沼泽的变化规律性。
其季节通量分布在-0.035—24.647 mg m-2h-1之间,春季CH4排放通量以G最高(高于5种森林沼泽0.3—43.1倍,P<0.05),C居中(高于3种针叶林沼泽13.7—26.7倍,P<0.05);夏、冬季M高于其他6种沼泽类型(1.3—769.2倍,P<0.05);秋季M高于G以外的其他5种沼泽类型(2.8—232.3倍,P<0.05),G高于3种针叶林沼泽(17.9—154.0倍,P<0.05)。由此可见,毛赤杨沼泽在夏、秋、冬季CH4排放通量均最高,灌丛沼泽在春季、秋季及草丛沼泽在春季相对较高。此外,在生长季、非生长季7种天然沼泽的CH4排放通量与其年通量变化趋势相同(即M>C、G、B>L-T、L-X、L-N)。
2.1.2 天然沼泽湿地土壤CO2排放通量及其季节动态趋势
由图2可以得到,小兴安岭7种天然沼泽土壤CO2排放季节动态趋势基本一致,自早春开始波动升高,进入夏季达到排放峰值,夏末秋初开始波动下降,直至冬季维持较低排放水平。但CO2排放峰值个数及其出现时间不同,其中,C、G、L-X和L-N在6月上旬至7月下旬(682.9、678.7、598.0、534.7 mg m-2h-1)及8月下旬(807.1、865.6、527.7、532.6 mg m-2h-1)出现2个排放峰值;而M、B、L-T仅在8月下旬或中旬(569.2 、531.5、504.0 mg m-2h-1)出现1个排放峰值。因此,小兴安岭7种天然沼泽类型的土壤CO2排放季节动态趋势存在双峰型与单峰型2种类型。
由表3可以得到,7种天然沼泽土壤CO2排放年通量及季节通量存在显著差异性。其年通量分布在157.40—231.06 mg m-2h-1之间,以G最高(显著高于5种森林沼泽28.7%—46.8%,P<0.05),C居中等(高于5种森林沼泽7.4%—22.5%,P>0.05),而5种森林沼泽最低且相近(4.2%—14.1%,P>0.05)。因此,小兴安岭7种天然沼泽湿地的土壤CO2排放年通量呈现出灌丛沼泽>草丛沼泽>森林沼泽的变化规律性。
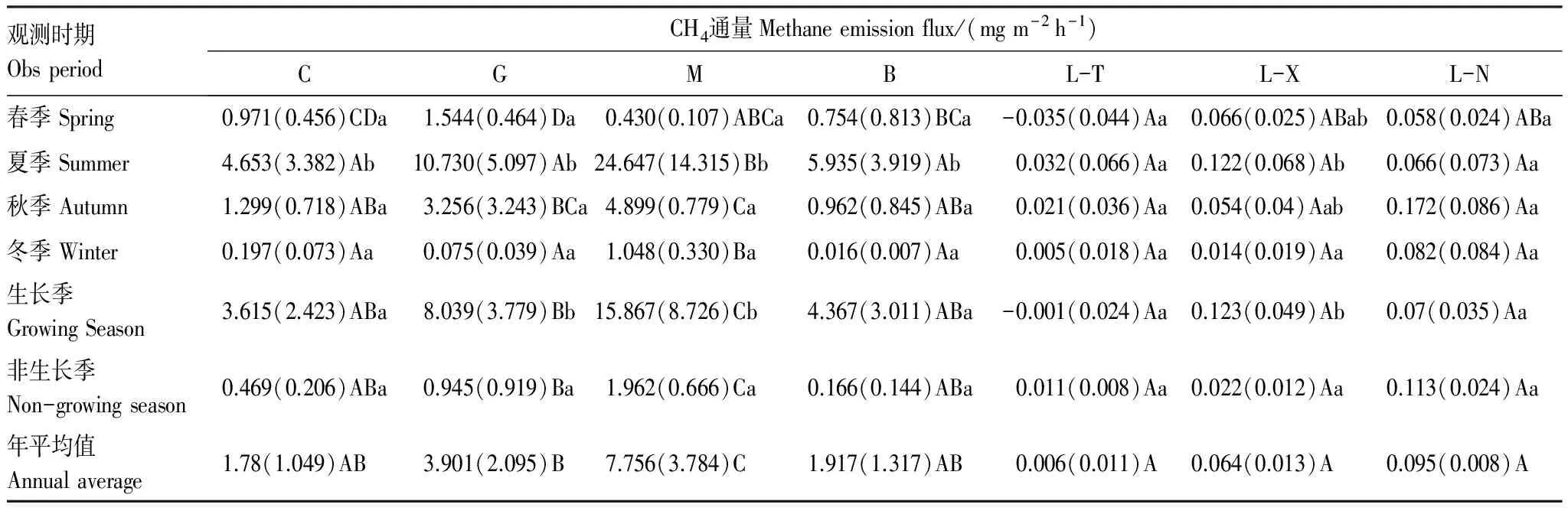
表2 小兴安岭7种天然沼泽湿地土壤CH4排放年通量及季节通量 (mg m-2 h-1)
大写字母为同一时间不同样地间比较,小写字母为同一样地不同季节间比较

图2 小兴安岭7种天然沼泽类型土壤CO2排放季节动态Fig.2 Seasonal dynamics of carbon dioxide emission fluxes from seven kinds of natural forested wetlands in Xiaoxing′anling of China
其季节通量分布在10.67—533.33 mg m-2h-1之间,春季G显著高于其他6种沼泽(29.6%—81.2%,P<0.05);夏季G显著高于L-T (51.5%,P<0.05);秋季C显著高于其他6种沼泽(43.6%—80.8%,P<0.05);冬季C显著高于除L-T以外的其他4种森林沼泽(46.1%—134.2%,P<0.05),G显著高于L-X(89.8%,P<0.05) 。在生长季G高于除L-X外的其他4种森林沼泽(39.0%—52.5%,P<0.05);在非生长季C高于M、L-T和L-X(47.7%—56.3%,P<0.05)。故灌丛沼泽在春季、夏季及生长季相对较高,草丛沼泽在秋季、冬季及非生长季相对较高,而森林沼泽在各季节均相对较低。

表3 小兴安岭7种天然沼泽湿地土壤CO2排放年通量及季节通量
大写字母为同一时间不同样地间比较,小写字母为同一样地不同季节间比较
2.1.3 天然森林沼泽湿地土壤CO2和CH4排放主控因子
由表4可以得到,小兴安岭7种天然沼泽CH4排放的主控因子不同。C、G、M、B 的CH4排放通量依次与30—40、5—40、5—40、10—40 cm土壤温度显著正相关;L-N的CH4排放通量与0—10 cm土壤温度显著负相关;而L-T和L-X的CH4排放通量与气温、土温及水位相关性均不显著。因此,小兴安岭草丛沼泽、灌丛沼泽和阔叶林沼泽及针叶林沼泽中的落叶松泥炭藓沼泽的CH4排放仅受土壤温度所控制。
7种天然沼泽土壤CO2排放的主控因子也不同,其中,C、G、M、L-T的CO2排放通量依次与0—30、0—40、0—30、0—10 cm土壤层温度及气温显著正相关;而B、L-X、L-N的CO2排放通量依次与0—30、0—20、0—30 cm土壤层温度及气温显著正相关,且与水位显著负相关。因此,小兴安岭草丛沼泽、灌丛沼泽、毛赤杨沼泽、落叶松苔草沼泽土壤CO2排放受气温及土壤温度所控制,白桦沼泽、落叶松藓类沼泽和落叶松泥炭藓沼泽则受气温、土温及水位三者综合控制。
2.2 天然森林沼泽湿地植被净初级生产力与年净固碳量
由表5可以得到,小兴安岭7种天然沼泽的植被净初级生产力((4.42±0.19)—(15.44±0.76) t hm-2a-1)和年净固碳量((2.05±0.09)—(6.75±0.27) tC hm-2a-1)存在较大差异性。其中,C的植被净初级生产力和年净固碳量显著高于G和5种森林沼泽(67.8%—249.3%和65.4%—229.3%,P<0.05),G、B和3种落叶松沼泽显著高于M(77.1%—108.1%和80.0%—99.0%,P<0.05)。因此,小兴安岭7种天然沼泽植被净初级生产力和年净固碳量呈现出草丛沼泽>灌丛沼泽、白桦沼泽和3种落叶松沼泽>毛赤杨沼泽的变化规律性。
表4 小兴安岭天然森林沼泽湿地温室气体排放与温度、水位的相关性
Table 4 Correlation between greenhouse gas flux with temperature and water table of seven kinds of forested wetlands in Xiaoxing′anling in China
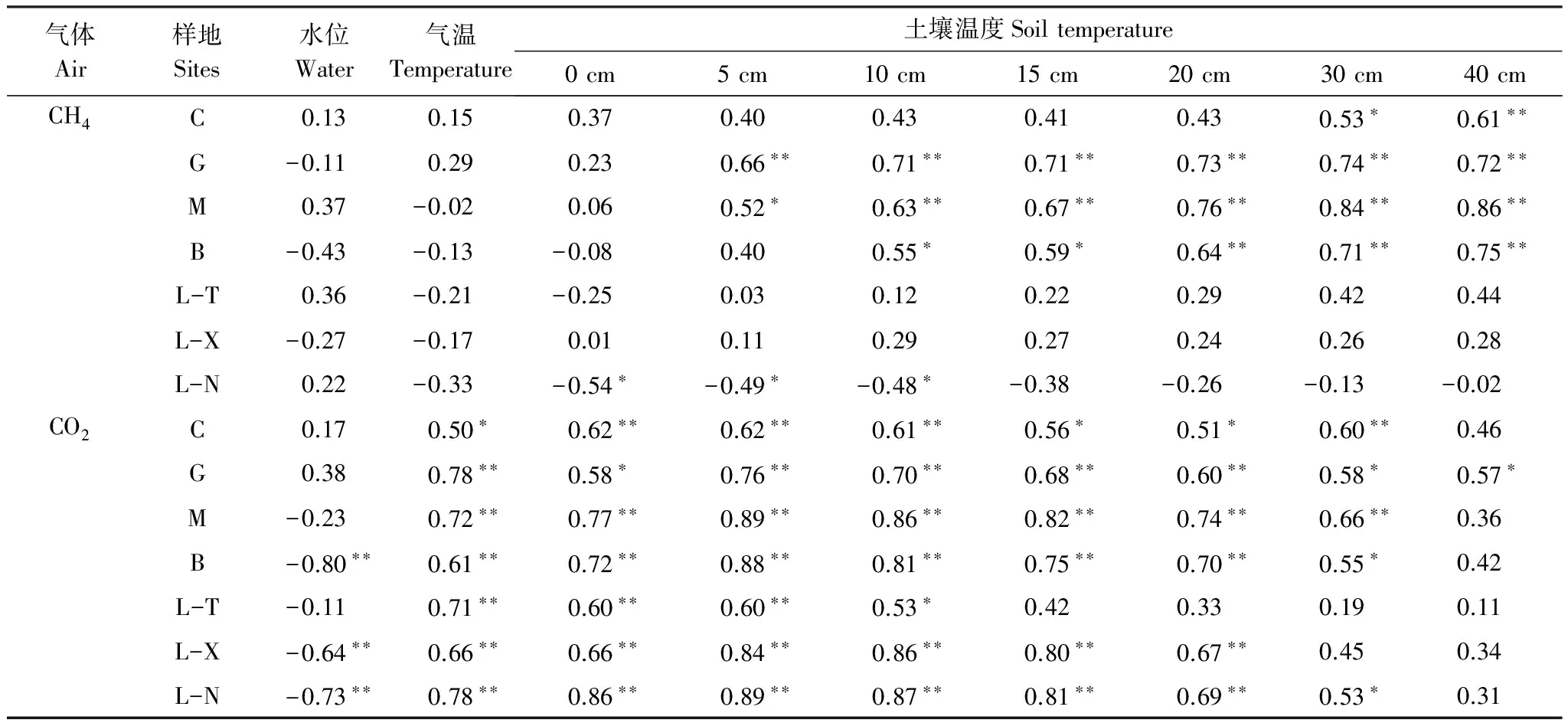
气体Air样地Sites水位Water气温Temperature土壤温度Soiltemperature0cm5cm10cm15cm20cm30cm40cmCH4C0.130.150.370.400.430.410.430.53∗0.61∗∗G-0.110.290.230.66∗∗0.71∗∗0.71∗∗0.73∗∗0.74∗∗0.72∗∗M0.37-0.020.060.52∗0.63∗∗0.67∗∗0.76∗∗0.84∗∗0.86∗∗B-0.43-0.13-0.080.400.55∗0.59∗0.64∗∗0.71∗∗0.75∗∗L-T0.36-0.21-0.250.030.120.220.290.420.44L-X-0.27-0.170.010.110.290.270.240.260.28L-N0.22-0.33-0.54∗-0.49∗-0.48∗-0.38-0.26-0.13-0.02CO2C0.170.50∗0.62∗∗0.62∗∗0.61∗∗0.56∗0.51∗0.60∗∗0.46G0.380.78∗∗0.58∗0.76∗∗0.70∗∗0.68∗∗0.60∗∗0.58∗0.57∗M-0.230.72∗∗0.77∗∗0.89∗∗0.86∗∗0.82∗∗0.74∗∗0.66∗∗0.36B-0.80∗∗0.61∗∗0.72∗∗0.88∗∗0.81∗∗0.75∗∗0.70∗∗0.55∗0.42L-T-0.110.71∗∗0.60∗∗0.60∗∗0.53∗0.420.330.190.11L-X-0.64∗∗0.66∗∗0.66∗∗0.84∗∗0.86∗∗0.80∗∗0.67∗∗0.450.34L-N-0.73∗∗0.78∗∗0.86∗∗0.89∗∗0.87∗∗0.81∗∗0.69∗∗0.53∗0.31
**在0.01 水平(双侧)上显著相关,*在0.05 水平(双侧)上显著相关.
7种天然沼泽类型各植被层的净初级生产力和年净固碳量也有所不同。草本层净初级生产力和年净固碳量以C为最高(显著高于其他6种沼泽1.3—54.1倍和1.3—55.3倍,P<0.05),G居中(显著高于5种森林沼泽3.8—23.2倍和4.0—23.6倍,P<0.05),而5种森林沼泽最低且相近;灌木层以G为最高(显著高于M、B和L-T 10.5—19.2倍和10.3—21.6倍,P<0.05),L-X和L-N居中(高于M、B和L-T 4.0—8.9倍和3.8—9.4倍,P>0.05),而后3种森林沼泽最低;乔木层以B和3种针叶林沼泽相对较高(显著高于M 94.1%—117.2%和88.7%—111.9%,P<0.05),M相对较低。因此,小兴安岭7种天然沼泽类型各植被层的净初级生产力和年净固碳量排序为草本层:草丛沼泽>灌丛沼泽>5种森林沼泽;灌木层:灌丛沼泽>落叶松藓类沼泽、落叶松泥炭藓沼泽>毛赤杨沼泽、白桦沼泽和落叶松苔草沼泽;乔木层:白桦沼泽和3种落叶松沼泽林>毛赤杨沼泽。
表5 小兴安岭7种天然森林沼泽湿地植被净初级生产力与年净固碳量
Table 5 Net primary productivity and net carbon sequestration of seven kinds of typical natural forested wetlands in Xiaoxing′anling in China

指标Item层次Layer处理TreatmentCGMBL-TL-XL-N净初级生产力乔木层3.38(0.24)A7.02(0.65)B7.08(1.64)B6.56(0.46)B7.34(2.27)BNetprimaryproductivity灌木层2.42(1.78)B0.12(0.01)A0.22(0.11)A0.21(0.14)A1.10(0.37)AB1.19(0.35)AB(NPP)/(thm-2a-1)草本层15.44(0.74)C6.78(1.32)B0.92(0.08)A1.40(0.39)A0.54(0.22)A0.77(0.28)A0.28(0.12)A植被15.44(0.74)C9.20(1.04)B4.42(0.19)A8.64(0.24)B7.83(1.71)B8.43(1.00)B8.81(2.18)B年净固碳量乔木层1.59(0.11)A3.28(0.31)B3.27(0.75)B3.00(0.22)B3.37(1.04)BAnnualnetcarbon灌木层1.13(0.84)B0.05(0.01)A0.10(0.05)A0.09(0.06)A0.48(0.16)AB0.52(0.16)ABsequestration(ANCS)/草本层6.75(0.27)C2.95(0.61)B0.41(0.04)A0.59(0.17)A0.22(0.07)A0.34(0.13)A0.12(0.05)A(thm-2a-1)植被6.75(0.27)C4.08(0.49)B2.05(0.09)A3.97(0.12)B3.59(0.76)B3.83(0.46)B4.02(0.99)B
*大写字母不同表示不同处理类型之间差异显著(P<0.05)
2.3 天然森林沼泽湿地的碳源/汇作用
由表6可以得到,小兴安岭7种天然沼泽类型的碳源/汇作用不同(其中,年碳排放量是根据所排放CH4和CO2中碳元素所占分子式比例进行折算加和所得)。C、B和L-N的生态系统净碳收支为正值(2.09、0.13、0.03 tC hm-2a-1),故三者为碳的吸收汇,但C的汇强显著高于B和L-N 15.1—68.7倍(P<0.05),B高于L-N但差异性不显著(3.3倍,P>0.05);而G、M、L-T、L-X的生态系统净碳收支为负值(-1.61、-2.32、-0.31、-0.41 tC hm-2a-1),故四者为碳的排放源,且M的源强显著高于L-T、L-X 4.7—6.5倍(P<0.05),G的源强也高于后两者但差异性不显著(2.9—4.2倍,P>0.05)。因此,小兴安岭7种天然沼泽类型中草丛沼泽为碳的强吸收汇,白桦沼泽和落叶松泥炭藓沼泽为弱吸收汇;毛赤杨沼泽和灌丛沼泽为碳强排放源,落叶松苔草沼泽和落叶松藓类沼泽为碳弱排放源。
此外,7种天然沼泽的植被固碳能力和土壤呼吸碳排能力也存在差异性。C的植被年净固碳量最高,G、B和3种针叶林沼泽居中,M最低(见2.2)。G的土壤年净碳排放量显著高于5种森林沼泽(30.4%—48.8%,P<0.05),C的年净碳排放量高于5种森林沼泽但差异性也不显著(6.6%—21.7%,P>0.05),5种森林沼泽的年净碳排放量相对较低且相近(1.8%—14.1%,P>0.05)。故草丛沼泽固碳最强而排放居中、灌丛沼泽排放最强而固碳居中、4种森林沼泽固碳居中排放最低(M除外),毛赤杨沼泽固碳最低排放也最低。
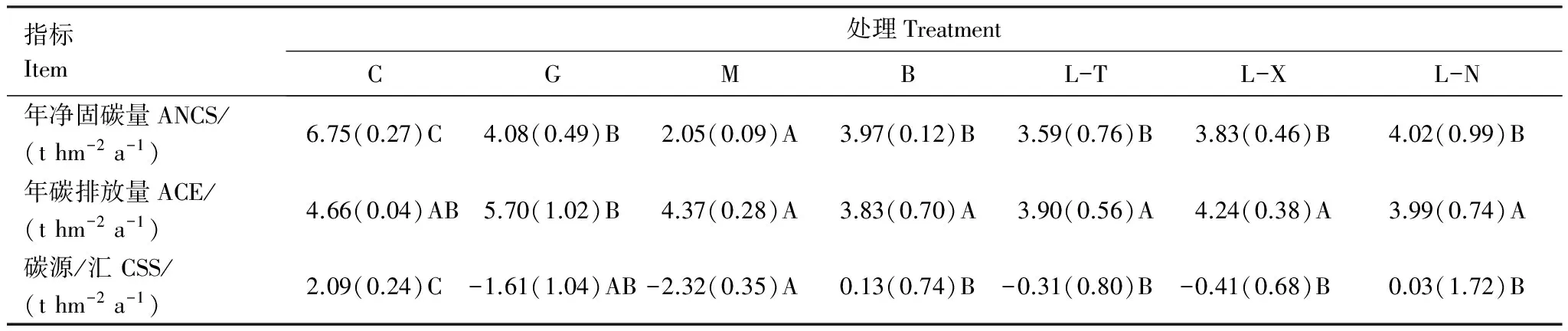
表6 小兴安岭7种典型天然森林沼泽湿地的碳源/汇
大写字母不同表示差异显著(P<0.05). ANCS:年净固碳量Annual net carbon sequestration(t hm-2a-1);ACE:年碳排放量Annualt carbon emission(t hm-2a-1); CSS:碳源/汇Carbon source/sink(t hm-2a-1)
3 结论与讨论
3.1 天然沼泽湿地CH4排放通量及其季节动态
小兴安岭7种天然沼泽CH4排放季节动态存在单峰型、多峰型和排放与吸收交替型与现有结论沼泽湿地CH4排放呈夏、秋季高于春季[18,39, 62- 64]及水位变化引起湿地CH4源/汇功能发生转化[25- 29]基本一致。其CH4年通量(0.006—7.76 mg m-2h-1)上限已超过温带森林沼泽的上限(4.0 mg m-2h-1)[16],但其变化幅度(1292倍)与北欧同一气候区不同湿地类型CH4通量相差几个数量级相一致[19]。
其原因则在于湿地甲烷排放受温度、水位、底物和特定微生物[8,20- 24]等因素影响,夏季随降水增多土壤水位升高,易形成甲烷产生菌所需的厌氧环境,加之夏季温度高,故CH4排放量增大。至于各类型CH4排放呈单峰型、多峰型及排放与吸收交替型,则在于各类型沿过渡带水分环境梯度分布不同,C、G处于过渡带下部地段,生长季水位高(平均水位3.7—10.0 cm),泥炭层始终处于水淹厌氧状态(水位已不是其限制因子),故CH4排放通量随温度的季节变化而变化,夏季温度高导致CH4排放呈单峰型;M、B处于过渡带的中下部地段,生长季水位有所降低(平均水位-10.4—-11.8 cm),且经常发生上下浮动,水位升高时厌氧反应空间增大,CH4排放量增大,水位下降时厌氧反应空间缩小,CH4排放量减少,故两者CH4通量呈多峰型;而L-T、L-X、L-N处于过渡带中上部地段,生长季水位进一步降低(平均水位-13.4—-16.5 cm),水位浮动加剧,使得有氧环境与厌氧环境经常发生转换,进而引起CH4排放与吸收交替发生。
3.2 天然沼泽湿地CO2排放通量及其季节动态
小兴安岭7种天然沼泽土壤CO2排放年通量(13.8—20.2 t hm-2a-1)与早期研究结果生长季土壤CO2排放量(13.8—22.0 t hm-2a-1)[33- 35,37]相近(此次去除植物根系呼吸部分);略低于我国三江平原草丛沼泽(19.7—25.7 t hm-2a-1)[65];但高于北美温带草丛泥炭地(4.7—13.2 t hm-2a-1)[66-67]及欧洲北方泥炭地(3.4—11.9 t hm-2a-1)[68- 70].故小兴安岭天然沼泽土壤应属于CO2强排放源。
小兴安岭7种天然沼泽土壤CO2排放季节动态存在双峰型和单峰型及季节分布存在夏>春≈秋>冬、夏>春>秋≈冬和夏>春>秋>冬型与现有结论沼泽CO2排放呈现夏季高于春、秋季[33- 35,37]基本一致。其原因在于CO2排放受土壤温度控制[65,71],7种天然沼泽夏季0—40 cm土壤温度较春、秋季提高1.1—8.7℃和0.8—10.4℃,有利于微生物分解活动,进而提高其CO2排放量。至于夏季形成双峰型或单峰型,则是温度与水位综合作用结果,如5种森林沼泽在8月下旬土壤温度高时,水位却较低(较平均水位降低6.1—21.5 cm),有氧空间增大,进而提高CO2排放量并形成排放峰值,这与水位显著低于泥炭表面时,CO2排放较水淹状态明显增大[31-32]相一致;相反,C、G、L-X、L-N处于过渡带下部谷地及L-X、L-N处于过渡带上部平缓地段,排水不畅,遇有强降水易形成水淹环境(如7月上旬出现50 mm强降雨),抑制土壤微生物有氧呼吸活动,降低CO2排放量,随着后期水位下降,CO2排放量又随温度升高而再一次增大,故导致CO2排放通量呈双峰型。
3.3 天然沼泽湿地CH4与CO2排放影响因子
小兴安岭C、G、M、B、L-N的CH4排放仅受土壤温度控制,而与水位相关性不显著,前者与温度是影响湿地甲烷排放的重要因子[8,18,20,72-73]一致;后者与水位是影响湿地CH4通量的重要因子[21,25-26]不同,这可能与本区各沼泽类型生长季平均水位高(-16.5—10.0 cm),全部或大部分泥炭层处于水淹状态,甲烷产生菌所需要的厌氧环境得以满足有关。
小兴安岭C、G、M、L-T土壤CO2排放受气温及不同土壤层温度控制,而B、L-X、L-N土壤CO2排放则受气温、土温及水位三者综合控制。前者与现有结论土壤温度是影响土壤呼吸速率和土壤微生物活性的一个重要因素[33- 35,74- 76]一致,后者与湿地CO2排放随温度升高和水位降低而增大[30- 32]一致。原因在于森林沼泽为季节性水淹环境,使得厌氧环境与有氧环境交替出现,故水淹时会抑制微生物的有氧分解活动,进而导致土壤CO2排放量减少,即两者存在显著负相关。
3.4 天然沼泽湿地植被净初级生产力与年净固碳量
小兴安岭7种天然沼泽湿地的植被净初级生产力(0.44—1.54 kg m-2a-1)与现有中国东北植被净初级生产力(0.6—1.4 kg m-2a-1)[77- 79]相近(除C略高于其上限及M略低于其下限外,G及其他4种森林沼泽(0.78—0.92 kg m-2a-1)高于其下限值(30.0%—53.3%),处于中等水平),但G及其他4种森林沼泽仅接近温带森林沼泽植被净初级生产力(1.0—1.5 kg m-2a-1)[80-81]的下限值(略低8.0%—22.0%),说明其是属于植被净初级生产力相对较低的温带森林沼泽类型。
其植被年净固碳量(0.21—0.68 kgC m-2a-1)与现有中国陆地植被平均固碳能力(0.49 kgC m-2a-1)[82]和全球平均植被固碳能力(0.41 kgC m-2a-1)[83]相比得到:C高于两者(38.8%—65.9%),G和4种森林沼泽(M除外)(0.36—0.41 kgC m-2a-1)略低于前者(16.3%—26.5%),但与后者相近(0.0%—12.2%),说明小兴安岭草丛沼泽固碳能力很强,但灌丛沼泽和森林沼泽的固碳能力多处于中等水平。
3.5 天然沼泽湿地碳源/汇作用
小兴安岭7种天然沼泽类型的碳源/汇作用并不同,即草丛沼泽为碳的强吸收汇,白桦沼泽和落叶松泥炭藓沼泽为弱吸收汇;毛赤杨沼泽和灌丛沼泽为碳的强排放源,落叶松苔草沼泽和落叶松藓类沼泽为碳的弱排放源。前者与现有研究结论天然湿地能够吸收固定大气中的CO2,可能成为大气温室气体的吸收汇[3- 7]相一致;而后者与天然湿地因排放大量甲烷,可能成为大气温室气体的排放源[8- 11]并不一致(因CH4碳排放量仅占其碳排放总量的0.1%—11.5%,而土壤CO2碳排放量却占其碳排放总量的88.5%—99.9%(表2和表3),故其源主要是来自土壤CO2排放而不是来自CH4排放)。
至于7种天然沼泽湿地的碳源/汇作用并不同的原因,可能主要是由于草丛沼泽的植被固碳能力最强,而其土壤呼吸碳排放量却处于中等排放水平,故使其成为碳的强吸收汇;而毛赤杨沼泽的植被固碳能力最低(因其寿命较短30a左右,调查样地处于衰退阶段),土壤呼吸碳排放量却与其他森林沼泽相近,灌丛沼泽的植被固碳能力虽属于中等水平,但其土壤呼吸碳排放量却最高,故使两者成为碳的强排放源;其他4种森林沼泽的固碳能力属于中等水平,土壤呼吸碳排放属于低排放水平(表6),故使其成为碳的弱吸收汇或碳的弱排放源。但这4种森林沼泽湿地的生态系统净碳收支并无显著的差异性(表6),说明四者目前基本维持碳的收支平衡状态。
[1] Gorham E. Northern peatlands: role in the carbon cycle and probable responses to climatic warming. Ecological Applications, 1991, 1(2): 182- 195.
[2] Maltby E, Immirzi P. Carbon dynamics in peatlands and other wetland soils regional and global perspectives. Chemosphere, 1993, 27(6): 999- 1023.
[3] Alongi D M, Trott L A, Pfitzner J. Deposition, mineralization, and storage of carbon and nitrogen in sediments of the far northern and northern Great Barrier Reef shelf. Continental Shelf Research, 2007, 27(20): 2595- 2622.
[4] Bedard-Haughn A, Jongbloed F, Akkerman J, Uijl A, De Jong E, Yates T, Pennock D. The effects of erosional and management history on soil organic carbon stores in ephemeral wetlands of hummocky agricultural landscapes. Geoderma, 2006, 135: 296- 306.
[5] Brevik E C, Homburg J A. A 5000 year record of carbon sequestration from a coastal lagoon and wetland complex, Southern California, USA. Catena, 2004, 57(3): 221- 232.
[6] Euliss Jr N H, Gleason R A, Olness A, McDougal R L, Murkin H R, Robarts R D, Bourbonniere R A, Warner B G. North American prairie wetlands are important nonforested land-based carbon storage sites. Science of the Total Environment, 2006, 361(1/3): 179- 188.
[7] Krogh L, Noergaard A, Hermansen M, Greve M H, Balstroem T, Breuning-Madsen H. Preliminary estimates of contemporary soil organic carbon stocks in Denmark using multiple datasets and four scaling-up methods. Agriculture, Ecosystems & Environment, 2003, 96(1/3): 19- 28.
[8] Whalen S C. Biogeochemistry of methane exchange between natural wetlands and the atmosphere. Environmental Engineering Science, 2005, 22(1): 73- 94.
[9] Carroll P, Crill P. Carbon balance of a temperate poor fen. Global Biogeochemical Cycles, 1997, 11(3): 349- 356.
[10] Whiting G J, Chanton J P. Greenhouse carbon balance of wetlands: methane emission versus carbon sequestration. Tellus B, 2001, 53(5): 521- 528.
[11] Wolf D C, Wagner G H. Carbon transformations and soil organic matter formation//Sylvia D M, Fuhrman J, Hartel P G, Zuberer D A, eds. Principles and applications of soil microbiology. 2nd ed. Upper Saddle River, NJ, USA: Prentice Hall, 2005: 285- 332.
[12] Armentano T V, Menges E S. Patterns of change in the carbon balance of organic soil-wetlands of the temperate zone. The Journal of Ecology, 1986, 74(3): 755- 774.
[13] Turetsky M, Wieder K, Halsey L, Vitt D. Current disturbance and the diminishing peatland carbon sink. Geophysical Research Letters, 2002, 29(11): 21- 1- 21- 4.
[14] Alford D P, Delaune R D, Lindau C W. Methane flux from Missippi River deltaic plain wetlands. Biogeochemistry, 1997, 37(3): 227- 236.
[15] Bartlett K B, Crill P M, Sebacher D I, Harriss R C, Wilson J O, Melack J M. Methane flux from the central Amazonian floodplain. Journal of Geophysical Research: Atmospheres (1984- 2012), 1988, 93(D2): 1571- 1582.
[16] Bartlett K B, Harriss R C. Review and assessment of methane emissions from wetlands. Chemosphere, 1993, 26(1/4): 261- 320.
[17] Kang H, Freeman C. The influence of hydrochemistry on methane emissions from two contrasting northern wetlands. Water, Air, and Soil Pollution, 2002, 141(1/4): 263- 272.
[18] Sun X X, Mu C C, Song C C. Seasonal and spatial variations of methane emissions from montane wetlands in Northeast China. Atmospheric Environment, 2011, 45(10): 1809- 1816.
[19] Søvik A K, Augustin J, Heikkinen K, Huttunen J T, Necki J M, Karjalainen S M, Kløve B, Liikanen A, Mander Ü, Puustinen M, Teiterg S, Wachniew P. Emission of the greenhouse gases nitrous oxide and methane from constructed wetlands in Europe. Journal of Environmental Quality, 2006, 35(6): 2360- 2373.
[20] Knoblauch C, Zimmermann U, Blumenberg M, Michaelis W, Pfeiffer E M. Methane turnover and temperature response of methane-oxidizing bacteria in permafrost-affected soils of northeast Siberia. Soil Biology and Biochemistry, 2008, 40(12): 3004- 3013.
[21] Waddington J M, Roulet N T, Swanson R V. Water table control of CH4emission enhancement by vascular plants in boreal peatlands. Journal of Geophysical Research: Atmospheres (1984- 2012), 1996, 101(D17): 22775- 22785.
[22] Bellisario L M, Bubier J L, Moore T R, Chanton J P. Controls on CH4emissions from a northern peatland. Global Biogeochemical Cycles, 1999, 13(1): 81- 91.
[23] Ström L, Ekberg A, Mastepanov M, Christensen T R. The effect of vascular plants on carbon turnover and methane emissions from a tundra wetland. Global Change Biology, 2003, 9(8): 1185- 1192.
[24] Tietz A, Langergraber G, Watzinger A, Haberl R, Kirschner A K T. Bacterial carbon utilization in vertical subsurface flow constructed wetlands. Water research, 2008, 42(6- 7): 1622- 1634.
[25] Augustin J, Merbach W, Rogasik J. Factors influencing nitrous oxide and methane emissions from minerotrophic fens in northeast Germany. Biology and Fertility of Soils, 1998, 28(1): 1- 4.
[26] Harriss R C, Sebacher D I, Day Jr F P. Methane flux in the great dismal swamp. Nature, 1982, 297(5868): 673- 674.
[27] Kelley C A, Martens C S, Ussler W. Methane dynamics across a tidally flooded riverbank margin. Limnology and Oceanography, 1995, 40(6): 1112- 1129.
[28] Moore T R, Roulet N T. Methane flux: water table relations in northern wetlands. Geophysical Research Letters, 1993, 20(7): 587- 590.
[29] Roulet N T, Ash R, Quinton W, Moore T. Methane flux from drained northern peatlands: effect of a persistent water table lowering on flux. Global Biogeochemical Cycles, 1993, 7(4): 749- 769.
[30] Bridgham S D, Richardson C J. Mechanisms controlling soil respiration (CO2and CH4) in southern peatlands. Soil Biology and Biochemistry, 1992, 24(11): 1089- 1099.
[31] Moore T, Dalva M. Methane and carbon dioxide exchange potentials of peat soils in aerobic and anaerobic laboratory incubations. Soil Biology and Biochemistry, 1997, 29(8): 1157- 1164.
[32] Waddington J M, Rotenberg P A, Warren F J. Peat CO2production in a natural and cutover peatland: implications for restoration. Biogeochemistry, 2001, 54(2): 115- 130.
[33] 牟长城, 程伟, 孙晓新, 吴云霞. 小兴安岭落叶松沼泽林土壤CO2, N2O和CH4的排放规律. 林业科学, 2010, 46(7): 7- 15.
[34] 牟长城, 刘霞, 孙晓新, 石兰英, 冯登军, 程伟, 吴云霞. 小兴安岭阔叶林沼泽土壤CO2、CH4和N2O排放规律及其影响因子. 生态学报, 2010, 30(17): 4598- 4608.
[35] 牟长城, 石兰英, 孙晓新. 小兴安岭典型草丛沼泽湿地CO2、CH4和N2O的排放动态及其影响因素. 植物生态学报, 2009, 33(3): 617- 623.
[36] Tolonen K, Turunen J. Carbon accumulation in mires in Finland//Proceedings of the International Workshop on Northern Peatlands in Global Climate Change. Helsinki, Finland: Publications of the Academy of Finland, 1996.
[37] 石兰英, 牟长城, 田新民. 小兴安岭典型沼泽湿地生态系统呼吸及其影响因子. 生态学杂志, 2009, 28(12): 2477- 2482.
[38] 石兰英, 牟长城, 田新民, 金建丽, 金志民. 小兴安岭不同沼泽甲烷排放及其影响因子. 生态学报, 2010, 30(19): 5289- 5295.
[39] 孙晓新, 牟长城, 石兰英, 程伟, 刘霞, 吴云霞, 冯登军. 小兴安岭森林沼泽甲烷排放及其影响因子. 植物生态学报, 2009, 33(3): 535- 545.
[40] Theroux S, Hartman W, He S M, Tringe S. Microbial diversity and carbon cycling in San Francisco Bay wetlands//Proceedings of the JGI 9th Annual User Meeting. Walnut Creek, CA (United States): JGIUM, 2014.
[41] Weston N B, Neubauer S C, Velinsky D J, Vile M A. Net ecosystem carbon exchange and the greenhouse gas balance of tidal marshes along an estuarine salinity gradient. Biogeochemistry, 2014, 120(1/3): 163- 189.
[42] 牟长城, 吴云霞, 李婉姝, 徐蕊. 采伐对小兴安岭落叶松-泥炭藓沼泽温室气体排放的影响. 应用生态学报, 2010, 21(2): 287- 293.
[43] 牟长城, 张博文, 韩丽冬, 于丽丽, 顾韩. 火干扰对小兴安岭白桦沼泽温室气体排放的短期影响. 应用生态学报, 2011, 22(4): 857- 865.
[44] 孙晓新, 牟长城, 宋长春, 吴云霞. 采伐对小兴安岭森林沼泽甲烷通量的影响. 土壤通报, 2011, 42(1): 190- 194.
[45] 于丽丽, 牟长城, 顾韩, 张博文. 火干扰对小兴安岭落叶松-苔草沼泽温室气体排放的影响. 生态学报, 2011, 31(18): 5180- 5191.
[46] 顾韩, 牟长城, 张博文. 火干扰对小兴安岭毛赤杨沼泽温室气体排放动态影响及其影响因素. 生态学报, 2012, 32(24): 7808- 7817.
[47] 顾韩, 牟长城, 张博文, 于丽丽. 火干扰对小兴安岭草丛、灌丛沼泽温室气体短期排放的影响. 生态学报, 2012, 32(19): 6044- 6055.
[48] 李婉姝, 牟长城, 吴云霞, 徐蕊. 小兴安岭地区天然沼泽林生产力与植被碳密度. 东北林业大学学报, 2010, 38(8): 39- 44.
[49] 牟长城, 韩士杰, 罗菊春, 王襄平. 长白山森林/沼泽生态交错带群落和环境梯度分析. 应用生态学报, 2001, 12(1): 1- 7.
[50] 牟长城, 万书成, 苏平, 宋宏文, 孙志虎. 长白山毛赤杨和白桦-沼泽生态交错带群落生物量分布格局. 应用生态学报, 2004, 15(12): 2211- 2216.
[51] 牟长城, 王彪, 卢慧翠, 包旭, 崔巍. 大兴安岭天然沼泽湿地生态系统碳储量. 生态学报, 2013, 33(16): 4956- 4965.
[52] 周文昌, 牟长城, 刘夏, 顾韩. 小兴安岭天然森林沼泽生态系统碳汇功能. 东北林业大学学报, 2012, 40(7): 71- 75, 127- 127.
[53] McCarty G W, Ritchie J C. Impact of soil movement on carbon sequestration in agricultural ecosystems. Environmental Pollution, 2002, 116(3): 423- 430.
[54] Trumbore S E, Harden J W. Accumulation and turnover of carbon in organic and mineral soils of the BOREAS northern study area. Journal of Geophysical Research: Atmospheres, 1997, 102(D24): 28817- 28830.
[55] Turunen J, Tomppo E, Tolonen K, Reinikainen A. Estimating carbon accumulation rates of undrained mires in Finland-application to boreal and subarctic regions. The Holocene, 2002, 12(1): 69- 80.
[56] Freeman C, Ostle N J, Fenner N, Kang H. A regulatory role for phenol oxidase during decomposition in peatlands. Soil Biology and Biochemistry, 2004, 36(10): 1663- 1667.
[57] Hanson R S, Hanson T E. Methanotrophic bacteria. Microbiological Reviews, 1996, 60(2): 439- 471.
[58] 郎惠卿. 中国湿地植被. 北京: 科学出版社, 1999.
[59] Wang Y S, Wang Y H. Quick measurement of CH4, CO2and N2O emissions from a short-plant ecosystem. Advances in Atmospheric Sciences, 2003, 20(5): 842- 844.
[60] 李兆富, 吕宪国, 杨青. 湿地土壤CO2通量研究进展. 生态学杂志, 2002, 21(6): 47- 50, 23- 23.
[61] 王跃思. 碳交换的箱法测定//陈泮勤. 地球系统碳循环. 北京: 科学出版社, 2004.
[62] Song C C, Wang Y S, Wang Y Y, Zhao Z C. Emission of CO2, CH4and N2O from freshwater marsh during freeze-thaw period in Northeast of China. Atmospheric Environment, 2006, 40(35): 6879- 6885.
[63] Wang Z P, Han X G. Diurnal variation in methane emissions in relation to plants and environmental variables in the Inner Mongolia marshes. Atmospheric Environment, 2005, 39(34): 6295- 6305.
[64] Huttunen J T, Nykänen H, Turunen J, Martikainen P J. Methane emissions from natural peatlands in the northern boreal zone in Finland, Fennoscandia. Atmospheric Environment, 2003, 37(1): 147- 151.
[65] Song C C, Yan B X, Wang Y S, Wang Y Y, Lou Y J, Zhao Z C. Fluxes of carbon dioxide and methane from swamp and impact factors in Sanjiang Plain, China. Chinese Science Bulletin, 2003, 48(24): 2749- 2753.
[66] Roehm C L, Roulet N T. Seasonal contribution of CO2fluxes in the annual C budget of a northern bog. Global Biogeochemical Cycles, 2003, 17(1): 29- 1- 29- 9.
[67] Wickland K P, Striegl R G, Mast M A, Clow D W. Carbon gas exchange at a southern Rocky Mountain wetland, 1996- 1998. Global Biogeochemical Cycles, 2001, 15(2): 321- 335.
[68] Alm J, Schulman L, Walden J, Nykänen H, Martikainen P J, Silvola J. Carbon balance of a boreal bog during a year with an exceptionally dry summer. Ecology, 1999, 80(1): 161- 174.
[69] Heikkinen J E P, Virtanen T, Huttunen J T, Elsakov V, Martikainen P J. Carbon balance in East European tundra. Global Biogeochemical Cycles, 2004, 18(1): GB1023.
[70] Waddington J M, Roulet N T. Carbon balance of a boreal patterned peatland. Global Change Biology, 2000, 6(1): 87- 97.
[71] Lafleur P M, Moore T R, Roulet N T, Frolking S. Ecosystem respiration in a cool temperate bog depends on peat temperature but not water table. Ecosystems, 2005, 8(6): 619- 629.
[72] Macdonald J A, Fowler D, Hargreaves K, Skiba U, Leith I D, Murray M B. Methane emission rates from a northern wetland; response to temperature, water table and transport. Atmospheric Environment, 1998, 32(19): 3219- 3227.
[73] Van Den Pol-Van Dasselaar A, Van Beusichem M L, Oenema O. Methane emissions from wet grasslands on peat soil in a nature preserve. Biogeochemistry, 1999, 44(2): 205- 220.
[74] Andrews J A, Matamala R, Westover K M, Schlesinger W H. Temperature effects on the diversity of soil heterotrophs and the δ13C of soil-respired CO2. Soil Biology and Biochemistry, 2000, 32(5): 699- 706.
[75] Zogg G P, Zak D R, Ringelberg D B, White D C, MacDonald N W, Pregitzer K S. Compositional and functional shifts in microbial communities due to soil warming. Soil Science Society of America Journal, 1997, 61(2): 475- 481.
[76] 宋长春, 王毅勇. 湿地生态系统土壤温度对气温的响应特征及对CO2排放的影响. 应用生态学报, 2006, 17(4): 625- 629.
[77] 毛德华, 王宗明, 罗玲, 韩佶兴. 1982- 2009年东北多年冻土区植被净初级生产力动态及其对全球变化的响应. 应用生态学报, 2012, 23(6): 1511- 1519.
[78] 张宪洲. 我国自然植被净第一性生产力的估算与分布. 自然资源, 1993, 15(1): 15- 21.
[79] 周广胜, 张新时. 全球气候变化的中国自然植被的净第一性生产力研究. 植物生态学报, 1996, 20(1): 11- 19.
[80] Campbell C, Vitt D H, Halsey L A, Campbell I D, Thormann M N, Bayley S E. Net primary production and standing biomass in northern continental wetlands. Information Report NOR-X- 369. Edmonton, Alberta: Northern Forestry Centre, Canadian Forest Service, 2000.
[81] Trettin C C, Jurgensen M F, Gale M R, McLaughlin J W. Soil carbon in northern forested wetlands: impacts of silvicultural practices//McFee W, Kelly J, eds. Carbon Forms and Functions in Forest Soils. Madison: Soil Science Society of America Inc, 1995.
[82] 何浩, 潘耀忠, 朱文泉, 刘旭拢, 张晴, 朱秀芳. 中国陆地生态系统服务价值测量. 应用生态学报, 2005, 16(6): 1122- 1127.
[83] 李银鹏, 季劲钧. 全球陆地生态系统与大气之间碳交换的模拟研究. 地理学报, 2001, 56(4): 379- 389.
Carbon source or sink of natural forested wetland ecosystem in Xiaoxing′anling region of China
LI Na,MU Changcheng*,WANG Biao,ZHANG Yan,MA Li
CenterforEcologicalResearch,NortheastForestryUniversity,Harbin150040,China
Carbon (CO2and CH4) emission fluxes, annual net carbon sequestration of vegetation, were studied from seven kinds of natural wetlands (Carexschmidtiimarsh-C,Betulaovalifolia-C.schmidtiishrub swamp-G,Alnussibirica-C.schmidtiiswamp-M,B.platyphylla-C.schmidtiiswamp-B,Larixgmelinii-C.schmidtiiswamp-L-T,L.gmelinii-moss swamp-L-X, andL.gmelinii-Sphagnumspp. swamp-L-N) in Xiaoxing′anling region of China using the static chamber gas chromatography method and the relative growth equation method to evaluate the carbon source or sink of natural forested wetlands based on net ecosystem carbon balance. The results showed that: 1) seasonal dynamics of CH4emission fluxes from seven kinds of natural wetlands could be divided into three types (single-peak, multi-peak, and emission and absorption alternating type) and CH4emission flux (0.006—7.756 mg m-2h-1) exhibited a different trend in M (which was 1.0—1291.7 times greater than the others,P< 0.05) >C, G, B (17.7—649.0 times greater than L-T, L-X, and L-N;P>0.05) >L-T, L-X, and L-N; 2)seasonal dynamics of CO2emission fluxes from the seven kinds of natural wetlands could be divided into two types (single-peak and double peak), and CO2emission flux (157.40—231.06 mg m-2h-1) exhibited an different trend in G (which was greater 28.7%—46.8% than forested wetlands,P< 0.05) >C(7.4%—22.5% than forested wetlands,P>0.05) >five kinds of forested wetlands; 3)CH4emission fluxes from C, G, M, B, and L-N were controlled by soil temperature, but that of L-T and L-X had no significant correlation with soil temperature at 0—40 cm. CO2emission fluxes from C, G, M, and L-T were controlled by air temperature and soil temperature, and that of B, L-X, and L-N were controlled by air temperature, soil temperature, and the water table level; 4) the annual net carbon sequestration of vegetation ((2.05±0.09)—(6.75±0.27) tC hm-2a-1) had a different trend in C (greater than the others by 65.4%—229.3%,P< 0.05)>G, B, L-T, L-X, and L-N (greater than M by 80.0%—99.0%,P< 0.05) >M; and 5) the sources and sinks of carbon were significantly different among the seven kinds of natural wetland types (-2.32—2.09 tC hm-2a-1). For example, C, B, and L-N exhibited varying degrees of carbon sinks, yet M, G, L-T, and L-X exhibited varying degrees of carbon sources. Therefore, marsh wetlands were greater carbon sinks and shrub wetlands were greater carbon sources, yet forested wetlands maintained overall carbon balance (except M) in the Xiaoxing′anling region of China.
Xiaoxing′anling; natural wetlands; carbon emission; carbon sequestration; carbon source/sink
国家自然科学基金项目(31370461)
2016- 01- 25; 网络出版日期:2016- 12- 19
10.5846/stxb201601250174
*通讯作者Corresponding author.E-mail: muccjs@163.com
李娜,牟长城,王彪,张妍,马莉.小兴安岭天然森林沼泽湿地生态系统碳源/汇.生态学报,2017,37(9):2880- 2893.
Li N,Mu C C,Wang B,Zhang Y,Ma L.Carbon source or sink of natural forested wetland ecosystem in Xiaoxing′anling region of China.Acta Ecologica Sinica,2017,37(9):2880- 2893.
