甜瓜雄花的几个胚胎学特征
2017-06-15周雪夏正艳曲胜男葛姝君赵丹孙晴刘林
周雪,夏正艳,曲胜男,葛姝君,赵丹,孙晴,刘林
(临沂大学药学院山东临沂276005)
甜瓜雄花的几个胚胎学特征
周雪,夏正艳,曲胜男,葛姝君,赵丹,孙晴,刘林
(临沂大学药学院山东临沂276005)
为阐明甜瓜雄花的胚胎学特征,用光学显微技术观察雄花花粉和花药壁的发育过程。结果表明,花粉母细胞减数分裂伴随胼胝质壁发育,减数分裂产生的4个小孢子在共同的胼胝质壁中呈四面体形排列。成熟期花粉粒直径约45 μm,积累大量脂滴,故属于脂质花粉。花药壁由外向内依次为表皮、药室内壁、中层和绒毡层,各含1层细胞;绒毡层在原位解体,为腺质绒毡层。表皮产生单细胞表皮毛,细胞膨大、伸长,细胞质浓,不含大液泡,仅含小液泡,生存至花粉成熟时期。
甜瓜;雄花;花药;花粉
杂性同株植物的同一植株具有雄花、雌花和两性花,杂性同株是两性花丢失雄性或雌性结构的结果,是植物向雌雄同株、雌雄异株和杂性异株过渡的关键一步[1-3],显然,杂性同株具有重要进化地位。甜瓜绝大部分品种都是杂性同株、雄花两性花同株[4]。对甜瓜雄花进行胚胎学研究,不仅对甜瓜雄花进化和保持的研究具有一定意义,也为甜瓜栽培和育种提供理论依据。被子植物花药和花粉发育的基本途径已基本清楚[5],但是关于甜瓜雄花花药和花粉发育,仍有以下几个方面的问题尚需进一步研究。第一,按照花粉发育的基本模式,小孢子母细胞减数分裂后产生的4个小孢子包埋在共同的胼胝质壁内形成四分体,四分体中小孢子的排列方式有2种可能,一种是四面体形排列,另一种是十字形排列[5],甜瓜四分体小孢子的排列方式尚未见报道;第二,花粉在发育过程中积累贮藏物质,为将来萌发和花粉管伸长储备营养和能量,成熟花粉的贮藏物质在一些植物中以淀粉为主,在另一些植物中以脂滴[5-8]为主,甜瓜雄花花粉贮藏物质的主要形式尚未见报道;第三,被子植物花药壁由表皮、药室内壁、中层和绒毡层组成,表皮和药室内壁各由1层细胞构成,而中层和绒毡层的细胞层数因植物种类而变化[5],绒毡层因发育模式不同而分为腺质绒毡层和变形绒毡层2种类型,甜瓜雄花花药壁的细胞层数以及绒毡层的类型尚无报道;第四,多数瓜类作物如西瓜花药表面不具有表皮毛[6],而甜瓜雄花花药表面有表皮毛,该表皮毛有何显微特征,属于腺性还是非腺性,尚无报道。研究这些问题,为甜瓜胚胎学增添资料,也为甜瓜育种工作提供参考。
针对上述4个方面的问题,笔者对甜瓜各节位尚未开放雄花花蕾及开放当天雄花花蕾中的花粉母细胞、花粉和花药壁进行显微观察。
1 材料与方法
试验于2014—2015年进行,所用甜瓜为‘绿宝’,来源于北京硕源种子有限公司。每年3月底用营养钵育苗,4月下旬移栽于临沂大学教学科研实习基地,露地栽培,常规管理,6月初取样。上午散粉前,取当天开放和未开放花蕾,包括顶芽中的幼蕾,取样植株数为8。自下而上各节位未开放花蕾长度:(23.9±2.7)、(14.0±2.9)、(9.5±1.0)、(7.7± 0.9)、(6.4±0.8)、(5.2±0.6)、(3.6±0.6)、(2.0±0.1)、(1.3±0.2)、(0.6±0.2)mm。为确保所取开放花蕾为取样当天开放,取样前一天下午天黑前将试验株上已开放的花蕾去掉,并做好标记。
从雄花蕾中分离出花药,较大的花药切成小块(长度小于1 mm),置于2%(φ)戊二醛中于室温下初固定4 h,再用1%(ω)锇酸于4°C下后固定4 h,然后用梯度酒精系列脱水,Epon-812树脂渗透、包埋。用玻璃刀切片,厚1.5 μm,切片用甲苯胺蓝染色,在光镜下观察并照相。
2 结果与分析
甜瓜雄花花药二室(图1-A),横切面上可观察到数个小孢子母细胞。小孢子母细胞呈多边形,边长不等,较长边20~25 μm,较短边约10 μm,细胞质浓,不含大液泡,小液泡分散在细胞质中,细胞核较大,球形,直径约10 μm,与周围花药壁细胞的核形成鲜明对比(图1-B)。通过花药纵切面观察,发育早期的小孢子母细胞较扁平(图1-C),该时期对应的花蕾长度在0.4~0.9 mm。
小孢子母细胞减数分裂开始时,形态方面发生两个标志性变化,一是产生胼胝质壁,二是原生质体由多边形变成椭球形(图1-D,E)。胼胝质壁在细胞角隅处较厚,厚度可达5 μm,而其他部位胼胝质壁相对较薄。原生质体椭球形,长约25 μm,宽约22 μm。小孢子母细胞减数分裂结束后,新产生的4个小孢子被共同的胼胝质壁包围,成为四分体,4个小孢子排列成四面体形(图1-F~H)。四分体直径约40 μm。减数分裂时期对应的花蕾长度在1~1.6 mm。
随着发育的进行,胼胝质壁消失,小孢子游离。刚从四分体游离出来的小孢子形状不稳定,多数较扁平,直径约40 μm,厚约20 μm(图1-I),后来变成椭球形,长约35 μm,宽约25 μm(图1-J~L)。小孢子是雄配子体的第1个细胞,即单核花粉。单核花粉经历不均等有丝分裂,产生1个大的营养细胞和1个小的生殖细胞。开花当天,花粉成熟,花粉粒直径约45 μm,含大量脂滴(图1-M,N)。小孢子时期对应的花蕾长度在1.6~4 mm,更大的花蕾则进入二细胞花粉时期,该时期花蕾长度范围较大。
花药壁分化出4层细胞,最外一层细胞为表皮,向内是药室内壁,再向内是单层细胞的中层,最里面是单层细胞的绒毡层。绒毡层细胞变化大,在小孢子母细胞时期呈扁平状,径向长度约4 μm,细胞质浓,液泡较小(图1-B,C);到减数分裂开始时,绒毡层细胞明显膨大,径向长度增至10 μm,液泡变大,位于细胞内侧(图1-D,E);四分体时期,绒毡层细胞进一步膨大,径向长度增至20 μm,液泡变小,细胞质内出现大量脂滴(图1-H);小孢子时期,绒毡层部分细胞凝缩、死亡,其他细胞横向膨大,向着药室的一侧形成曲面(图1-K,L)。开花前,绒毡层细胞基本消失,只剩下部分极度凝缩的细胞,呈凸透镜形(图1-M)。绒毡层细胞始终保持原来的位置,直到完全解体、消失。
花药表皮分化出大量表皮毛,表皮毛一直生存到花粉成熟时期。表皮毛为单细胞结构,细胞肥大,细胞质浓,不含大液泡,只含小液泡,具有腺性细胞的特征,属于腺毛。小孢子母细胞时期,腺毛长约20 μm(图1-O),随着花粉发育进展,腺毛逐渐伸长,至小孢子时期,长达150 μm(图1-P~R)。花粉成熟时期,腺毛细胞内小液泡增多,细胞表面发生凹陷,含有小液泡(图1-S)。
3 讨论
被子植物小孢子母细胞减数分裂伴随着胼胝质壁的发生,减数分裂产生的4个小孢子被共同的胼胝壁包围在一起,即形成四分体,四分体内小孢子排列成四面体形或十字形。笔者研究表明,甜瓜四分体内小孢子排列成四面体形,这与西瓜[6]和南瓜[8]等瓜类作物中的情况一样,表明小孢子在四分体中的四面体形排列可能是瓜类作物的普遍特征。
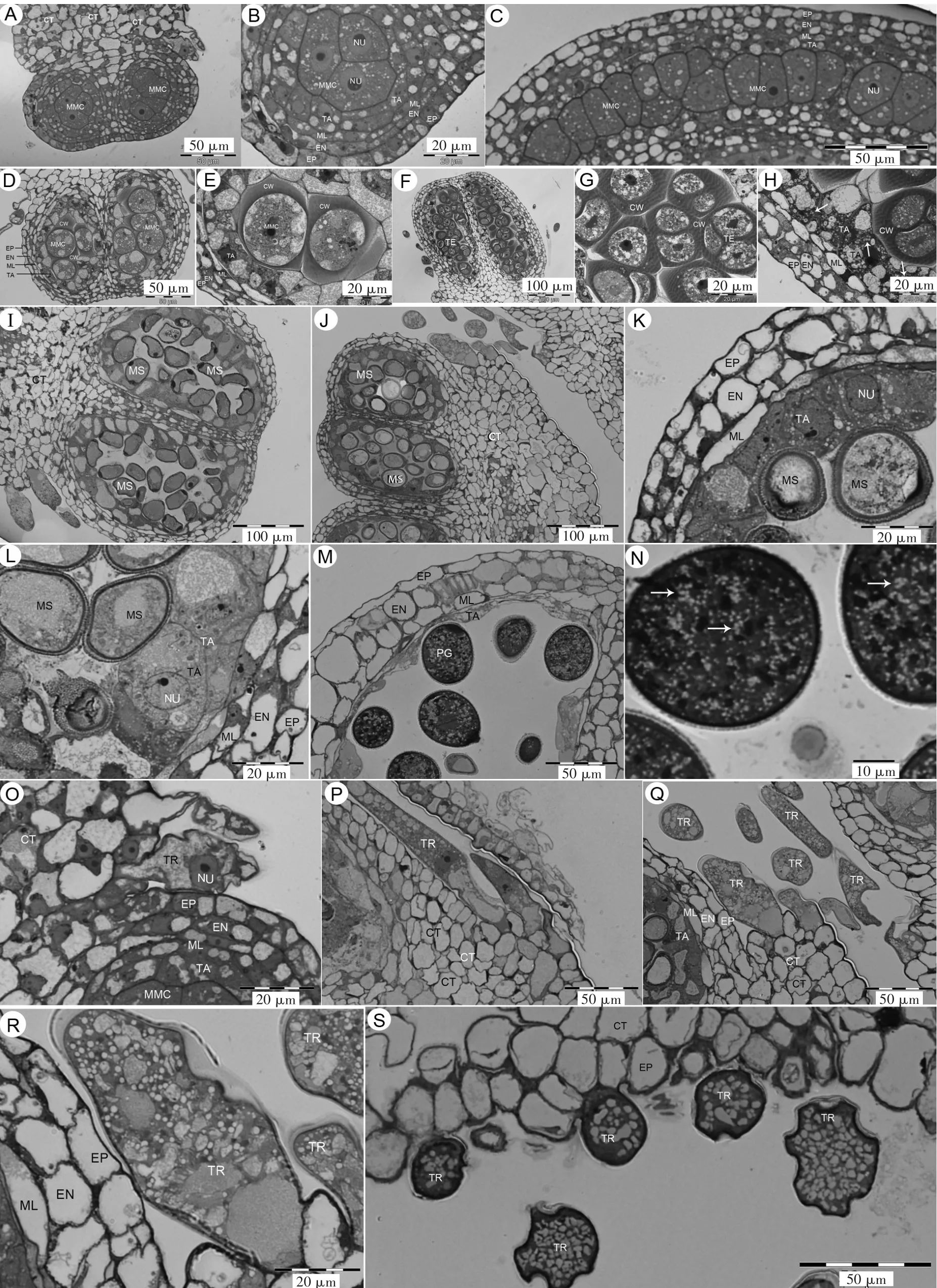
图1 甜瓜花药与花粉
被子植物花粉粒由营养细胞和生殖细胞或2个精子构成,营养细胞为花粉萌发和花粉管的伸长储备营养和能量。贮藏物质的主要形式包括淀粉和脂滴,如果成熟花粉中的贮藏物质以淀粉为主,这样的花粉属于淀粉质花粉;如果以脂滴为主,这样的花粉就属于脂质花粉[5]。本研究结果表明,甜瓜成熟花粉中的贮藏物质是脂滴,因而甜瓜花粉属于脂质花粉。西瓜[9]花粉也是脂质花粉,而南瓜花粉是淀粉质花粉。贮藏物质形式似乎与花粉粒大小有相关性,体积大的花粉倾向于积累能量较低的淀粉,体积较小的花粉倾向于积累能量较高的脂滴,例如,南瓜[8]花粉粒直径达150 μm,积累淀粉;西瓜[9]花粉粒直径60 μm,积累脂滴;甜瓜花粉粒直径45μm,积累脂滴;玉凤花属[10-11]花粉粒直径小于25μm,既不含淀粉粒也不含脂滴。玉凤花属花粉粒营养细胞含有无数小泡,这些小泡可能是贮藏可溶性物质的场所。
被子植物花药壁的中层和绒毡层的细胞层数因植物种类而变化,中层的细胞层数在有些植物中为2层,如西瓜[6]和南瓜[7],而在有些植物中仅1层,本研究结果表明,甜瓜花药壁的中层只含1层细胞。除表皮外,花药壁各层细胞均由壁细胞衍生,壁细胞分裂次数决定花药壁的细胞层数,即壁细胞分裂次数多,则花药壁细胞层数多,显然,与西瓜[6]和南瓜[7]相比,甜瓜花药壁细胞分裂次数较少。多数植物绒毡层由1层细胞组成,如西瓜[6]和南瓜[7];仅少数植物绒毡层由2层细胞组成[5]。甜瓜绒毡层只含1层细胞,与西瓜、南瓜等瓜类作物一致。绒毡层可分为腺质绒毡层和变形绒毡层2种类型,前者在整个发育过程中始终保持在原来的位置,通过细胞的内表面产生分泌物,为花粉发育提供营养和能量,直到细胞完全自溶;后者的主要特征是细胞较早地发生内壁和径向壁解体,原生质突出并移动到花药室中并融合形成周原质团,被发育中的花粉粒吸收利用。甜瓜绒毡层细胞始终保持在原来的位置,直到完全自溶,因而为腺质绒毡层。
植物表皮毛由表皮细胞发育而成,有多细胞表皮毛和单细胞表皮毛,有分支表皮毛或不分支表皮毛,有具分泌能力的表皮毛(腺毛),有不具分泌能力的非腺性表皮毛[12-15]。单细胞腺毛膨大、细胞质丰富,多细胞腺毛通常分化为头部和柄部[16-17]。表皮毛通常发生在幼茎、叶、花萼等器官表面,本研究结果还表明,表皮毛还可发生在花药表面,甜瓜花药表皮产生单细胞表皮毛,细胞肥大,细胞质浓,属于单细胞腺毛。甜瓜花药壁腺毛的功能尚不清楚,可能具有生态学意义,有助于吸引传粉昆虫。
[1]CUEVAS J,POLITO V S.The role of staminate flowers in the breeding systems ofOlea europaea(Oleaceae):an andromone⁃cious,wind-pollinated taxon[J].Annals of Botany,2004,93(5): 547-553.
[2]TALLEJO-MARIN M,RAUSHER M D.The role of male flowers in andromonoecious species:energetic cost and siring success in Solanum carolinenseL[J].Evolution,2007,61(2):404-412.
[3]ZHANG T,TAN D Y.An examination of the function of male flowers in an andromonoecious shrubCapparis spinosa[J].Journal of Integrative Plant Biology,2009,51(3):316-324.
[4]KOUONON L C,JACQUEMART A L,BI A I Z,et al.Repro⁃ductive biology of the andromonoeciousCucumis melosubsp. agrestis(Cucurbitaceae)[J].Annals of Botany,2009,104(6): 1129-1139.
[5]胡适宜.被子植物胚胎学[M].北京:高等教育出版社,1984:20-66.
[6]刘林,白华举.西瓜花药与花粉显微结构研究[J].中国西瓜甜瓜,2003,16(5):13-14.
[7]张涛,刘林.南瓜花粉发育过程中绒毡层细胞的变化[J].中国瓜菜,2014,27(3):20-21.
[8]张涛,董文才,刘林.南瓜花粉发育的解剖观察[J].中国瓜菜,2013,26(6):44-46.
[9]LIU L.Ultrastructural study on dynamics of plastids and mito⁃chondria during microgametogenesis in watermelon[J].Micron,2012,43:412-417.
[10]LIU L.Ultramicroscopic examination of mature massulae of Habenaria arinaria(Orchidaceae)[J].Micron,2015,74:1-7.
[11]刘林.十字兰成熟花粉的超微特征[J].园艺学报,2015,42(9):1831-1836.
[12]AKERS C P,WEYBREW J A,LONG R C.Ultrastructure of glandular trichomes of leaves ofNicotiana tabacumL.cv.XAN⁃THI[J].American Journal of Botany,1978,65(3):282-292.
[13]FIGUEIREDO A C,PAIS M S.Ultrastructural aspects of the glandular cells from the secrtory trichomes and from the cell suspension cultures ofAchillea millefoliumL.ssp.millefolium[J].Annals of Botany,1994,74(2):179-190.
[14]ASCENSAO L,MARQUES N,PAIS M S.Peltate glandular tgrichomes ofLeonotis leonurusleaves:ultrastructure and histo⁃chemical characterization of secretions[J].International Journal of Plant Sciences,1997,158(3):249-258.
[15]DUKE S O,PAUL R N.Development and fine structure of glandular trichomes ofArtemisia annuaL[J].International Journal of Plant Sciences,1993,154(1):107-118.
[16]GRAVANO E,TANI C,BENNICI A,et al.The ultrastructure of glandular trichomes ofPhillyrea latifoliaL.(Oleaceae)leaves[J].Annals of Botany,1998,81(2):327-335.
[17]刘林.杏叶腺毛的结构和功能研究[J].果树学报,2012,29(3):355-358.
Embryological characteristics of male flowers of Cucumis melo
ZHOU Xue,XIA Zhengyan,QU Shengnan,GE Shujun,ZHAO Dan,SUN Qing,LIU Lin
(College of Pharmacy,Linyi University,Linyi 276005,Shandong,China)
In order to understand the anther wall structure and pollen developmental features of Cucumis melo,anthers of different developmental stages were observed by microscopy.The meiosis of the microspore mother cell gave rise to four microspores,which were arranged in a tetrahedroid pattern within a common callose wall.Mature pollen grains,which were 45 μm in diameter,contained numerous lipid droplets and therefore belonged to lipidic type.The anther wall was comprised of epidermis,endothecium,middle layer and tapetum,each consisting of a single celled layer.The tapetal cell was degenerated on its original location and so the tapetum belonged to glandular type.The epidermis developed a lot of single celled trichomes.The trichome cell was enlarged and contained rich cytoplasm and survived until the pollen grains got mature.
Cucumis melo;Male flower;Anther;Pollen
2016-03-03;
:2017-03-23
山东省自然科学基金(ZR2015HL115);国家级大学生创新创业训练计划项目(201610452025)
:周雪,女,在读本科生,研究方向为药用植物资源学。E-mail:zhouxue_1020@163.com
刘林,男,教授,研究方向为植物发育与细胞生物学。E-mail:liulin@lyu.edu.cn
