盐、碱胁迫对药用植物碱地肤生长及其茎叶离子含量的影响
2017-06-13郭立泉张淑芳田小海孙吉凤陆静梅
麻 莹,郭立泉,张淑芳,田小海,孙吉凤,陆静梅
(1.长春医学高等专科学校基础医学部,吉林 长春 130031;2.东北师范大学环境学院,国家环境保护湿地生态与植被恢复重点实验室,吉林 长春 130117;3.东北师范大学生命科学学院,吉林 长春 130024)
盐、碱胁迫对药用植物碱地肤生长及其茎叶离子含量的影响
麻 莹1,2,郭立泉2,张淑芳1,田小海1,孙吉凤1,陆静梅3
(1.长春医学高等专科学校基础医学部,吉林 长春 130031;2.东北师范大学环境学院,国家环境保护湿地生态与植被恢复重点实验室,吉林 长春 130117;3.东北师范大学生命科学学院,吉林 长春 130024)
对碱地肤(Kochiasieversiana)幼苗进行盐胁迫或碱胁迫动态处理,通过动态监测碱地肤的生长及无机离子含量等指标的变化情况,探讨了碱地肤生长及离子积累的动态特点.结果表明:随着盐、碱胁迫的延长,碱地肤含水量、鲜重和干重等生长指标均不同程度地受到影响,且碱胁迫影响更大.盐、碱胁迫均抑制碱地肤的生长,碱胁迫抑制更为严重.碱地肤并非肉质化植物,其在长时间的渗透胁迫和高pH下仍能保持高含水量,此特殊现象可能是决定碱地肤具有高度抗盐碱特性的生理特点之一.随着盐、碱胁迫的延长,碱地肤植株对K+,Na+吸收不存在拮抗竞争,且在144 h的盐、碱胁迫下仍保持着较低的bNa+/bK+比值,这种排Na+、吸K+的特殊吸收机制对碱地肤的抗盐、碱性具有重要意义.此外,与盐胁迫相比,碱胁迫下其bNa+/bK+比值更低,这可能是碱地肤适应高pH盐碱生境的重要原因之一.
碱地肤;盐、碱胁迫;无机离子;生长

碱地肤(Kochiasieversianus)是一种能够在pH高达10以上的土壤生境下生存的抗盐碱的藜科牧草[3].碱地肤还具有一定的药用价值,其全草及种子均可以当作中药材使用.[4]由于其既具有极强的耐盐碱能力又具有很好的药用价值,因而成为中国东北治理碱化草地的首选牧草之一.目前,有关碱地肤耐盐碱性的研究,主要涉及盐、碱胁迫下碱地肤的种子萌发、生长、茎叶溶质积累及其有机酸的分布等生理特点方面.[5-7]然而以上研究,主要以不同浓度的盐、碱处理为胁迫变量,本文突破以往盐碱处理方法,采用相同浓度的盐或碱进行胁迫,以不同胁迫时间作为胁迫变量,探讨了碱地肤生长及其茎叶无机离子含量的动态变化情况.
碱地肤属富集 Na+,K+等离子的盐碱植物之一[3],主要通过选择吸收耗能低的Na+,K+,Cl-等无机离子来降低细胞渗透势[6].然而,有关盐碱生境中碱地肤体内无机离子动态积累的研究尚未见报道,了解盐碱胁迫对碱地肤生长及离子含量的动态变化特点,将对盐碱草原的治理及恢复有着极其重要的科学和经济价值.
鉴于分布有碱地肤的中国东北碱化草地土壤含盐特点,本文分别用中性盐(NaCl,Na2SO4)和碱性盐(NaHCO3,Na2CO3)模拟盐、碱胁迫条件,并对碱地肤(K.sieversiana)的幼苗进行胁迫处理[7],通过动态监测碱地肤生长及其茎叶无机离子积累的变化特点,探讨了碱地肤特殊的离子积累的动态特点,以为碱地肤适应盐碱生境的特殊生理机制研究提供参考.
1 材料与方法
1.1 材料培养
碱地肤(K.sieversiana)种子采集于吉林省西部天然草原.将种子播种于直径17 cm盛有干净细砂的塑料花盆内,出苗后每2天用Hoagland营养液透灌1次,其他时间用蒸馏水补充水分蒸发.每盆定苗16株.实验在室外进行,3次重复[7],处理期间人工遮雨.
1.2 盐胁迫和碱胁迫条件
分别将2种中性盐NaCl,Na2SO4和2种碱性盐NaHCO3,Na2CO3按照物质的量比1∶1混合,分别作为盐胁迫组和碱胁迫组[7].盐、碱胁迫组的总盐浓度均为200 mmol/L,以保证两种胁迫处理液的Na+浓度和总离子浓度相同,只有pH不同,而且碱胁迫组溶液的pH值与典型的碱地肤生境的pH值相近(9.80~10.10)[8].以不含胁迫盐的Hoagland营养液作为对照组.盐胁迫组、碱胁迫组和对照组溶液的pH值分别为6.58,9.96及6.56.
1.3 胁迫处理
苗龄6周后,选取66盆长势一致的碱地肤苗,随机分为22组,每组3盆(3次重复).其中:8个组为对照组(CK),只浇营养液,在8个时间点(0,16,24,36,48,72,96,144 h)取样;7个组为盐胁迫组,用中性盐处理,在7个时间点(16,24,36,48,72,96,144 h)取样;7个组为碱胁迫组,用碱性盐处理,在7个时间点(同盐胁迫组)取样.进行处理时,以含有相应胁迫盐的营养液作为处理液,每盆苗用500 mL盐或碱的处理液浇灌,2次/d透灌,使植株处于完全胁迫状态;对照组只浇500 mL营养液.
1.4 取样
分别按不同的处理时间点逐盆取出碱地肤植株,分别用自来水、蒸馏水冲洗植株,用滤纸吸干植株表面水分.在子叶痕处剪开,将植株分成茎叶和根2个部分,分别称取以上部分的鲜重(WF).分别称取5 g 茎叶部分冻干,并记录其干重,冻干样品用于离子含量测定;其余鲜样105℃杀青15 min后,40℃下真空干燥至恒重,计算总干重(WD).
1.5 指标测定
(1) 生长指标:含水量(%)=(WF-WD)/WF×100%.干重、鲜重均以每株茎叶部分质量表示(g).
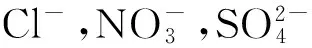
1.6 数据分析
数据处理等统计学分析均用统计学软件SPSS完成.结果数据均用3次重复数据的平均值±标准误差表示,检验水平P<0.05.
2 结果分析
2.1 盐、碱胁迫下碱地肤动态生长情况
盐、碱胁迫下碱地肤的地上部分含水量、鲜重、干重的动态变化情况见图1.从图1A可以看出,盐、碱胁迫条件下,碱地肤地上部分含水量均降低(P<0.01).而碱胁迫(F=191.80)下降幅度明显大于盐胁迫(F=29.57),但在长时间(144 h)的碱胁迫下仍能保持较高的含水量(75%),而这不是肉质化的结果,这一特殊的耐盐机制值得深入研究.
从图1B可以看出,三种生长条件对碱地肤地上部分鲜重的影响不同:非胁迫条件下,碱地肤每株地上部分鲜重随生长时间显著增加(P<0.01);盐胁迫下,鲜重增加不明显(P>0.05);而碱胁迫下,鲜重显著降低(P<0.01),这可能与碱胁迫下含水量显著降低有关.
从图1C可以看出,随着生长时间的延长,非胁迫和盐胁迫条件下,碱地肤每株地上部分干重显著增加(P<0.01),且非胁迫条件下其增加的幅度(F=25.43)大于盐胁迫(F=5.62);而碱胁迫下,其干重增加不明显(P>0.05).可见,盐碱胁迫特别是碱胁迫不利于碱地肤地上部分干重的增加.综上,盐、碱胁迫均抑制碱地肤的生长,而碱胁迫抑制的更为严重.

图1 盐、碱胁迫下碱地肤的地上部分含水量、鲜重、干重的动态变化
2.2 盐、碱胁迫下碱地肤茎叶的阳离子动态积累特点
盐、碱胁迫下Na+,K+的含量及bNa+/bK+比值的动态变化情况见图2.从图2可以看出,碱胁迫下植株茎叶部分对K+,Na+离子的吸收高于盐胁迫.在盐、碱胁迫48 h内,Na+含量随着胁迫时间的延长有所增加,且两种胁迫的作用相近,一旦胁迫时间超过48 h,其含量急剧增加(P<0.01),其中碱胁迫的作用更大(见图2A).K+变化较为特殊,在16 h的盐、碱处理下其含量均升高,而16 h后随之下降;当胁迫时间超过36 h后,盐胁迫下其含量基本保持不变(P>0.05),碱胁迫下随着时间的延长其含量有所增加(见图2B).在胁迫的整个过程中,碱胁迫下的K+含量始终大于盐胁迫.可见,碱胁迫下Na+的急剧积累,并没有抑制K+的吸收.从图2C还可看出,随着胁迫时间的增加,bNa+/bK+比值有所增加,但碱胁迫下bNa+/bK+小于盐胁迫(96 h除外).动态观测到Na+大量积累而K+含量并不随着降低,即碱地肤对K+,Na+吸收并不存在拮抗竞争现象[6],这与多数盐生植物的K+,Na+吸收机制不同[10].

图2 盐、碱胁迫下Na+(A),K+(B)的含量及bNa+/bK+(C)的动态变化
2.3 盐、碱胁迫下碱地肤茎叶的阴离子动态积累特点
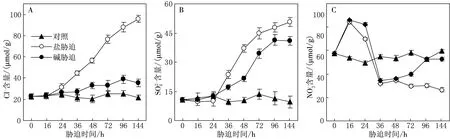
图3 盐、碱胁迫下等阴离子含量的动态变化
3 讨论
3.1 盐、碱胁迫对碱地肤生长的影响
生长受到抑制是植物对盐、碱胁迫普遍而直接的反应[11],碱地肤也是这样.与非胁迫条件相比,盐、碱胁迫均不利于碱地肤干重、鲜重的积累,其中碱胁迫的抑制作用更强.碱地肤要适应碱胁迫(高pH)生境必然要消耗更多的物质和能量,因此,其生长受到抑制更严重.这也许是碱地肤在逆境下的重要生存对策之一.
降低自身含水量是植物响应渗透胁迫进行渗透调节的一种既迅速又经济的途径[12],碱地肤也不例外,随着盐、碱胁迫强度的增大其含水量逐渐下降(见图1A).与芦苇(Phragmitesaustralis)相比[12],在长时间(144 h)的碱胁迫下碱地肤仍能保持较高的含水量(75%),而碱地肤并非肉质化植物,其在长时间的渗透胁迫和高pH下仍能保持高含水量,这一特殊现象可能正是碱地肤具有高度抗盐碱特性的生理特点之一.细胞含水量高必然引起液泡体积增大、细胞表面积增大、原生质层变薄等后果,而这些因素更加能够促进Na+,K+(见图2A和图2B)的进入,同时高含水量还可以最大限度地浓缩细胞质中溶质浓度,稀释液泡中有害离子的浓度[12].
3.2 盐碱胁迫对碱地肤茎叶无机离子动态积累的影响
植物的主要抗盐机制就是Na+及其他离子的代谢机制.碱地肤通过体内大量积累Na+来降低细胞渗透势,缓解盐害[6].Na+主要通过非选择性阳离子通道和高亲和K+通道两种途径进入根细胞[13].由于Na+和K+水合半径相似,致使大量Na+竞争K+通道,从而抑制了K+的吸收.而盐、碱胁迫下碱地肤对K+,Na+吸收不存在拮抗竞争现象,这与我们以前的研究结果一致[6].K+作为植物生长的必需元素,其选择吸收既可以降低细胞渗透势,保持细胞质内外渗透平衡,还可缓解盐、碱胁迫导致的K+亏缺而引起的生长抑制[6],因此,K+特殊的吸收机制可能对碱地肤的抗盐、碱特性具有重要意义,尚有待进一步研究.
越来越多的结果表明,在盐胁迫条件下,限制Na+吸收、增加Na+外排,同时保证K+的吸收,维持细胞质低bNa+/bK+是植物抗盐性的关键[14].众所周知,高pH的碱胁迫对植物的伤害远远大于盐胁迫.而碱胁迫下碱地肤的bNa+/bK+低于盐胁迫,这对碱胁迫下碱地肤可维持正常的生理代谢是非常有利的.这可能也是碱地肤适应高pH(9.8~10.2)天然盐碱生境的重要原因之一[8].

[1] YANG C W,CHONG J N,LI C Y,et al.Osmotic adjustment and ion balance traits of an alkali resistant halophyteKochiasieversianaduring adaptation to salt and alkali conditions [J].Plant Soil,2007,294:263-276.
[2] 石德成,殷立娟.盐(NaCl)与碱(Na2CO3)对星星草胁迫作用的差异[J].植物学报,1993,35(2):144-149.
[3] 郑慧莹,李建东.松嫩平原盐生植物及盐碱草地恢复[M].北京:科学出版社,1999:12.
[4] 翟延君,冯夏红,康延国,等.碱地肤的生药鉴定[J].中药材,1996,19(6):283-285.
[5] 颜宏,赵伟,秦峰梅,等.盐碱胁迫对碱地肤、地肤种子萌发以及幼苗生长的影响[J].东北师大学报(自然科学版),2006,38(4):117-123.
[6] 麻莹,曲冰冰,郭立泉,等.盐碱混合胁迫下抗碱盐生植物碱地肤的生长及其茎叶中溶质积累特点[J].草业学报,2007,16(4):25-33.
[7] 麻莹,郭立泉,张淑芳,等.盐碱胁迫下抗碱牧草碱地肤溶质积累、分布特点及有机酸的生理贡献[J].草业学报,2013,22(1):193-200.
[8] MA Y,GUO L Q,WANG H X,et al.Accumulation,distribution,and physiological contribution of oxalic acid and other solutes in an alkali-resistant forage plant,Kochiasieversiana,during adaptation to saline and alkaline conditions[J].Journal of Plant Nutrition and Soil Science,2011,174(4):655-663.
[9] 王保山,赵可夫.小麦叶片中Na+和K+提取方法比较[J].植物生理学通讯,1995(3):50-52.
[10] SHI D C,WANG D L.Effect s of various salt-alkali mixed stresses onAneurolepidiumchinense(T rin.)Kitag[J].Plant and Soil,2005,271:15-26.
[11] NALIWAJSKI M.R,SKLODOWSKA,M.The oxidative stress and antioxidant systems in cucumber cells during acclimation to salinity [J].J Plant Physiol,2014,58:47-54.
[12] LISSNER J,SCHIERUP H,COMN F A,et al.Effect of climate on the salt tolerance of twoPhragmitesaustralispopulations.-I.Growth,inorganic solutes,nitrogen relations and osmoregulation[J].Aquat Bot,1999,64:317-333.
[13] MUNNS R,TESTER M.Mechanisms of salinity tolerance[J].Annu Rev Plant Biol,2008,59:651-681.
[14] ZHU J K.Regulation of ion homeostasis under salt stress[J].Curr Opin Plant Biol,2003(6):441-445.
(责任编辑:方 林)
Effects of salt and alkali stresses on growth and ions accumulation in shoot of medicinal plantKochiasieversiana
MA Ying1,2,GUO Li-quan2,ZHANG Shu-fang1,TIAN Xiao-hai1,SUN Ji-feng1,LU Jing-mei3
(1.Department of Basic Medicine,Changchun Medical College,Changchun 130031,China;2.State Environmental Protection Key Laboratory of Wetland Ecology and Vegetation Restoration,School of Environment,Northeast Normal University,Changchun 130117,China;3.School of Life Sciences,Northeast Normal University,Changchun 130024,China)
In this study,seedlings ofKochiasieversiana(Pall.)C.A.Mey were exposed to the dynamic treatment:salt stress and alkali stress.The dynamic characteristics ofK.sieversianagrowth and ions accumulation were investigated by determining and analyzing the growth index,the concentrations of ions and so on.The results showed that the growth indexes,such as water content,dry weight,fresh weight and so on,inK.sieversiana,were affected to various degrees with increasing stress time under salt and alkali stresses,and alkali stress was greater than salt stress.Clearly,both salt and alkali stresses inhibited the growth ofK.sieversiana,and the inhibition of alkali stress was more serious than salt stress.K.sieversianais not a kind of succulence plant,whereas it could still keep high water content under long-time osmotic stress and high pH condition,which may endow it with high saline-alkali tolerance as a primary physiological characteristic.There is no antagonism competition between absorption of K+and Na+inK.sieversianawith increasing stress time under salt and alkali stresses,and it could still low value ofbNa+/bK+under 144 h alkali stress.This kind of special absorption mechanism of K+and Na+may have great significance in saline-alkali tolerance ofK.sieversiana.Moreover,the value ofbNa+/bK+is lower under alkali stress than salt stress,which may be one of the important reasons for adaption to saline-alkaline habitat contained high pH inK.sieversiana.
Kochiasieversiana;salt/alkali stresses;inorganic ion;growth
1000-1832(2017)02-0111-05
10.16163/j.cnki.22-1123/n.2017.02.021
2016-10-17
国家自然科学基金资助项目(41271231);东北师范大学教育部重点实验室开放课题(130028691);吉林省科技发展计划项目(20140204023NY);吉林省教育厅“十三五”科学技术项目(JJKH20171071KJ).
麻莹(1981—),女,博士,副教授,主要从事植物形态及逆境生理学研究;通讯作者:陆静梅(1952—),女,教授,博士研究生导师,主要从事植物形态及生理学研究.
Q 945.78 [学科代码] 180·5140
A
