基于高通量测序的郫县豆瓣后发酵期细菌多样性研究
2017-06-05赵红宇徐炜桢杨国华刘元福
赵红宇,徐炜桢,杨国华,刘元福,岳 鹏,张 良,*
基于高通量测序的郫县豆瓣后发酵期细菌多样性研究
赵红宇1,徐炜桢1,杨国华2,刘元福3,岳 鹏2,张 良1,*
(1.西华大学食品与生物工程学院,食品生物技术四川省高校重点实验室,四川 成都 610039;2.四川省丹 丹郫县豆瓣集团股份有限公司,四川省豆瓣酿制技术工程实验室,四川 成都 611732;3.四川友联味业食品有限公司,四川 成都 611732)
为研究郫县豆瓣后发酵过程中细菌变化规律,揭示其特有“日晒夜露”工艺的发酵本质,采用MiSeq测序分析其从后发酵1 周至后发酵6 a期间一共8 个时间点的细菌群落演替变化情况。结果表明,郫县豆瓣后发酵过程中共有57 个门类群、174 个纲类群、321 个目类群、535 个科类群、921 个属类群的细菌参与演替变化。后发酵时间对郫县豆瓣的细菌群落组成具有重要影响,随着后发酵的进行肠杆菌科和链形菌科持续减少,而鞘脂单胞菌科、芽孢杆菌科、梭菌科和消化链球菌科的细菌则出现先增加后减少,其峰值大多出现在1~3 a,但后发酵6 a的郫县豆瓣细菌群落较其他样品呈明显偏低。该方法发现了大量的非培养细菌和未报道细菌,所得细菌多样信息更接近于样品微生态,更能够全面解析自然发酵调味品—郫县豆瓣的细菌多样性,为传统产业的现代化改造和食品质量安全控制提供了科学支撑。
郫县豆瓣;细菌群落;高通量测序
郫县豆瓣属中国传统发酵食品,迄今为止已有300多年的历史,被列为中国非物质文化遗产。郫县豆瓣不仅生产工艺独特,也以其味辣香醇、黏稠绒实、红棕油亮、酱香浓郁等特点在我国酱类产品中独树一帜,堪称川菜之魂[1-2]。截止2015年末,“郫县豆瓣”品牌价值已达607.16亿 元,位列“加工食品类地理标志产品”全国第一;当年产品总产量达到110万t,实现工业产值102亿 元,出口世界绝大部分国家和地区,创汇超过4 000万美元[3]。
郫县豆瓣的生产包括前期发酵和后熟发酵两个阶段,前期发酵主要是指霉瓣子的制曲和辣椒坯的预处理[2]。后熟发酵主要是将成熟霉瓣子和成熟辣椒坯按比例配料混合,加入适量食盐和水,进入发酵池发酵,经过一定时期的翻晒和陈化,即是郫县豆瓣特有的日晒夜露工艺[4-5]。
近年来,国内利用变性梯度凝胶电泳(denaturing gradient gel electrophoresis,DGGE)、温度梯度凝胶电泳(temperature gradient gel electrophoresis,TGGE)[6]、DNA克隆文库[7]、末端限制性片段长度多态性(terminalrestriction fragment length polymorphism,T-RFLP)分析[8]、16S rRNA基因文库[9]等免培养技术,在分析研究发酵食品生产过程中微生物的功能作用方面,作出了大量卓有成效的成绩,其直接从分子水平上研究微生物资源,避免了传统基于微生物分离培养分析方法的局限性,在分析复杂环境微生物群落结构中具有优势。但这些方法都存在一些问题,如测序通量低、操作复杂、干扰因素多、准确率不高等,第2代基于宏基因组的深度测序技术又称为高通量测序(high-throughput sequencing,HTS)技术,具有检测通量更高、用时更少、准确度更高以及检测费用更低等优点,能更可靠、更全真、更直接地反映微生物的群落构成、功能特性、变化演替和多样性[10]。
目前,HTS技术在发酵调味品研究中得到了一定的应用,如韩国学者对海鲜酱[11]、酱粉[12]、泡菜[13]、鱼酱[14]和豆瓣辣酱[15]的发酵微生物多样性已经进行了解析;欧洲学者更是从宏基因组的角度评述了该技术给食品微生物群落变化研究带来的重大影响变化[16]。但是,国内外尚鲜有利用该技术分析郫县豆瓣后发酵期细菌多样性的研究报道。
本研究采用MiSeq测序,对郫县豆瓣后发酵过程中从入池到发酵6 a合计8 个时间点的样品进行分析,以期揭示郫县豆瓣不同后发酵阶段的细菌多样性,加深对传统本土调味品发酵机制的认识,为传统产业的现代化改造和食品质量安全控制提供科学支撑。
1 材料与方法
1.1 材料
样品取自郫县当地某郫县豆瓣生产企业。选取后发酵1 周、3 个月、6 个月、1、2、3、5、6 a的郫县豆瓣样品(共计8 个样品,按发酵时间长短分别编号为1W、3M、6M、1Y、2Y、3Y、5Y及6Y)。每个时间节点的样品选取同一后发酵时间的不同发酵池进行3 次重复取样。郫县豆瓣在后发酵时,每隔一定时间需进行搅拌、翻晒,故取样时从每个发酵池的上层(距表面0~20 cm)、中层(距表面30~50 cm)、下层(距池底0~20 cm)各取25 g左右,混合均匀后密封,低温运至实验室,于-70 ℃保存备用。测序结果表明来源于同一批次豆瓣的菌群非常相似(数据未列出),所以把同一批次的3 个重复样品的序列合并为一个数据以便进行后续分析。1.2 试剂
土壤DNA提取试剂盒 上海Sangon Biotech公司;Taq DNA聚合酶、dNTPs、16S rRNA通用引物 宝生物工程(大连)有限公司;二代测序快速DNA建库试剂盒美国Illumina公司。
1.3 仪器与设备
NanoDrop 2000测定仪 美国Thermo公司;台式离心机、聚合酶链式反应(polymerase chain reaction,PCR)仪 德国Eppendorf公司;MiSeq测序仪 美国Illumina公司。
1.4 方法
1.4.1 DNA提取
豆瓣基因组DNA采用土壤DNA提取试剂盒提取。DNA纯度(A260nm/280nm及A260nm/230nm)采用NanoDrop 2000测定仪测定。
1.4.2 PCR扩增
PCR扩增采用16S rRNA的V4~V5高变区的通用引物515F(5’-GTGCCAGC MGCCGCGGTAA-3’)和909R(5’-CCCCGYCAATTCMTTTRAGT-3’)[17]并在515F引物5’端加不同的barcode。详细PCR步骤参考Li Xiangzhen等[18]的方法略作改进,PCR扩增条件为:94 ℃预变性5 min,94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸50 s,30 个循环;最后72 ℃再延伸10 min。
1.4.3 MiSeq测序
1% 琼脂糖凝胶电泳检测PCR产物,试剂盒回收纯化PCR产 物,按等物质的量比例混合成测序文库后,用MiSeq测序仪进行双端测序。
1.5 数据统计分析
采用QIIME pipeline去除低质量原始测序数据,使用Uchime去除嵌合体序列[19]。以16S rRNA序列97%相似度作为操作分类单元(operational taxonomic units,OTUs)的划分标准。使用QIIME平台计算PD Whole Tree和Chao1多样性指数,并进行主坐标分析(principal coordinate analysis,PcoA)[20]。
2 结果与分析
2.1 郫县豆瓣不同后发酵阶段细菌多样性分析

表 1 基于97%相似性水平所计算的多样性指数Table 1 Diversity indices calculated based on a cutoff of 97%similarity
测序数据经软件拼接、过滤等处理后,8 个不同后发酵阶段郫县样品平均获得17 155 条细菌序列。不同后发酵阶段的样品细菌多样性指数如表1所示。从表1可以看出,随着发酵时间的延长,样品当中的细菌多样性总体呈现先下降后上升再下降的趋势。但后发酵时间过长(6 a)的样品细菌多样性最低,与其他后发酵阶段的细菌多样性存在明显差异。

图 1 豆瓣样品的稀释性曲线Fig. 1 Rarefaction curves for Pixian soybean paste samples
样品稀释曲线图1,也与表2情况类似,其曲线在小于6 000 条序列时OTU)数量随样品序列数的增加迅速变化,在大于14 000 条序列后OTU数量变化逐渐趋于平缓,这说明本测序深度条件下已覆盖了8 个样本中绝大部分的测序数据,能够较为全面地反映郫县豆瓣后发酵阶段的细菌多样性变化情况。
群落生态学可通过样品的OTU和Chao1来反映微生物群落的丰度和多样性。产业界通常将后发酵6 月以上的郫县豆瓣视为成熟,但是为保障产品的均一性,出厂前还会将不同后发酵期的豆瓣进行混合。若将业界普遍公认后发酵期6 个月至2 a的郫县豆瓣作为成熟样本计算,本研究通过MiSeq测序获得的平均序列数和划分的OTU均显著高于韩国辣酱[15]和韩国泡菜[13](平均序列数分别为15 733、1 606、2 656 条,OTU分别为781、144、121),这说明本土郫县豆瓣细菌的丰富度和多样性均较 高。
郫县豆瓣后发酵过程中细菌的多样性是 特有的区域环境条件、霉瓣子制曲工艺、自然接种过程(“日晒夜露”工艺)与发酵坯高盐含量(盐 含量大于15%)之间强烈选择和协同进化的结果。中国地理标志产品郫县豆瓣的主产区郫县,在川西平原腹心地带,属盆地中亚热带湿润气候,年均气温15.7 ℃,相对湿度70%,年降雨量960 mm,多集中在5~9 月,全年无霜期282 d,平均日照1 286.9 h[21],该条件为微生物菌群的生存提供了极好的环境,使细菌群落丰富性和多样性极高,造就了郫县豆瓣独特的外观品质和内在风味。
2.2 郫县豆瓣后发酵期细菌群落结构变化分析
对不同后发酵阶段郫县豆瓣的8 个样品中的细菌群落,利用Illumina双末端高通量测序,发现不同样品中共有57 个门类群、174 个纲类群、321 个目类群、535 个科类群、921 个属类群。

图 2 门水平下郫县豆瓣细菌群落变化Fig. 2 Changes in bacteria l community in Pixian soybean paste during fermentation at phylum level
如图2所示,细 菌群落中门水平平均含量大于1%的是变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)、蓝藻门(Cyanobacteria)、拟杆菌门(Bacter oidetes)、泉古菌门(Crenarchaeota)、绿弯菌门(Chloroflexi)、放线菌门(Actinobacteria)和广古菌门(Euryarchaeota),分别平均占总细菌数的46.15%、32.22%、4.12%、3.97%、2.86%、2.41%、2.28%和1.11%。发酵前期主要是厚壁菌门,而发酵后期主要是变形菌门。

图 3 纲水平下郫县豆瓣细菌群落变化Fig. 3 Changes in bacterial community in Pixian soybean paste during fermentation at class level
如图3所示,纲水平平均含量大于1%的分别是隶属变形菌门的γ-变形菌纲(Gamma Proteobacteria)、α-变形菌纲(Alpha Proteobacteria)和β-变形菌纲(Beta Proteobacteria),分别占总细菌群数的26.77%、15.59%和2.82%;隶属厚壁菌门的芽孢杆菌纲(Bacilli)和梭菌纲(Clostridia),分别平均占总菌群数的19.99%和12.18%;隶属蓝藻门的Chloroplast,占4.05%;隶属泉古菌门的奇古菌纲(Thaumarchaeota),占1.83%;隶属拟杆菌门的拟杆菌纲(Bacteroidia),占1.81%;隶属放线菌门的放线菌纲(Actinobacteria)占1.60%;隶属绿弯菌门的厌氧绳菌纲(Anaerolineae),占1.05%。随着后发酵的进行,γ-变形菌纲、Chloroplast、厌氧绳菌纲逐渐变少,α-变形菌纲、芽孢杆菌纲、梭菌纲、β-变形菌纲、奇古菌纲、拟杆菌纲出现先增多后减少,其峰值大多出现在后发酵1~2 a。

图 4 目水平下郫县豆瓣细菌群落变化Fig. 4 Changes in bacterial community in Pixian soybean paste during fermentation at order level
如图4所示,目水平平均含量大于1%的分别有肠杆菌目(Enterobacteriales)20.04%、芽孢杆菌目(Bacillales)14.12%、鞘脂单胞菌目(Sphingomonadales)13.56%、梭菌目(Clostridiales)11.78%、乳杆菌目(Lactobacillales)5.23%、链形菌目(Streptophyta)3.98%、假单胞菌目(Pseudomonadales)2.46%、伯克氏菌目(Burkholderiales)2.37%、 海洋螺菌目(Oceanospirillales)2.31%、拟杆菌目(Bacteroidales)1.81%、亚硝基球形菌目(Nitrososphaerales)1.78%、放线菌目(Actinomycetales)1.53%和根瘤菌目(Rhizobiales)1.08%。随着后发酵的进行,肠杆菌目、链形菌目均逐渐变少,芽孢杆菌目、鞘脂单胞菌目、梭菌目、乳杆菌目、伯克氏菌目、海洋螺菌目均出现先增多后减小,其中芽孢杆菌目、乳杆菌目、海洋螺菌目的峰值出现在后发酵3 个月,而鞘脂单胞菌目、梭菌目和伯克氏菌目的峰值出现在1~3 a。

图 5 科水平下郫县豆瓣细菌群落变化Fig. 5 Changes in bacterial community in Pixian soybean p aste during fermentation at family level
如图5所示,科水平平均含量大于1%分别有肠杆菌科(Enterobacteriaceae)20.04%、鞘脂单胞菌科(Sphingomonadaceae)13.48%、芽孢杆菌科(Bacillaceae)9.13%、梭菌科(Clostridiaceae)6.91%、链形菌科(Streptophyta)3.98%、葡萄球菌科(Staphylococcaceae)3.48%、消化链球菌科(Peptostreptococcaceae)2.68%、杆状盐单胞菌科(Halomonadaceae)2.28%、明串珠菌科(Leuconostocaceae)2.19%、乳杆菌科(Lactobacillaceae)2.06%、假单胞菌科(Pseudomonadaceae)1.87%、亚硝基球形菌科(Nitrososphaeraceae)1.78%、产碱菌科(Alcaligenaceae)1.03%和草酸杆菌科(Oxalobacteraceae)1.02%。
Nam等[15]利用高通量测序技术研究韩国辣酱发现了与以往报道不一致的结果,以往认为枯草芽孢杆菌(Bacillus subtilis)是韩国辣椒发酵的优势微生物,但该研究认为芽孢杆菌属(Bacillus species)的微生物在韩国中心区域生产的辣酱当中是优势微生物,而其他区域生产的辣酱当中乳酸菌(Lactic acid)才是优势微生物。Park等[13]利用焦磷酸测序分析泡菜发酵过程中细菌群落的演替情况,发现人工接种和自然发酵的泡菜细菌群落在发酵初期存在差异,但随着发酵的进行差异越来越小,发酵末期几乎没有明显差异;造成泡菜当中细菌群落的差异主要原因是生产工艺、泡菜种类不同。邹艳玲[22]利用16S rRNA基因文库技术从后发酵期的郫县豆瓣中共得到了102 个克隆子,发现芽孢杆菌科、葡萄球菌科和肠杆菌科是主要细菌。张琦等[6]利用PCR-DGGE技术发现郫县豆瓣后发酵过程中优势细菌为乳酸菌,魏斯氏菌属的Weissella confusa和乳酸菌属的Lactobacillus lactis是发酵初期的优势菌,而发酵后期优势菌则为乳酸菌属的Lactobacillus namurensis。赵辉平等[23]应用纯培养方法和16S rRNA基因系统发育分析郫县豆瓣中可培养细菌的多样性,从所得的209 株纯培养细菌中筛选出35 株典型菌株,共包含8 个属,其中Bacillus是优势类群。张大凤等[24]将酿酒酵母、产朊假丝酵母、植物乳杆菌和耐盐四联球菌共培养制成复合微生物菌剂用于郫县豆瓣后熟发酵,显著提高了产品品质。
本研究结果表明随着郫县豆瓣后发酵的进行,肠杆菌科和链形菌科持续减少,而鞘脂单胞菌科、芽孢杆菌科、梭菌科和消化链球菌科则出现先增加后减少的情况,其峰值大多出现在1~3 a。值得注意的是,后发酵时间6 a的郫县豆瓣细菌群落与其他样品呈现明显差异,其多样性较小,细菌群落主要集中于肠杆菌科(89.31%)。鉴于郫县豆瓣后发酵期细菌变化研究报道较少的现状,本研究结果与以往研究结果存在较大差异,可能因为郫县豆瓣后发酵期的细菌群落组成与发酵管理、取样季节、原料来源、组成配比、所在区域均具有密切关系。以往研究由于样品采集(采集季节差异和产地差异)、研究方法(分离纯化与免培养)等差异造成了不一致,还需要长期追踪并积累细菌演替变化的数据,以期进一步认识郫县豆瓣后发酵过程当中细菌群落组成差异的原因。
已有研究结果,无论是取样追踪的时间跨度,还是所得微生物菌群数据丰富程度,以及确定优势菌群的全面性,与本结果相比都具有一定局限。这说明HTS技术可以快速、高效地对郫县豆瓣中细菌结构进行精确的分析,尤其是为揭示其一些功能作用的机制提供了新的研究方法。陈玲等[9]利用16S rDNA克隆文库法与高通量测序法在浓香型大曲微生物群落结构分析对比研究过程中,也得出了与本研究类似的结论。
但是,该技术仍有一定的不足,尽管该技术可以提供海量的数据,但质量有待提高, 而且存在序列读长;另外,由于所得测序数据量巨大,经软件拼接、过滤等处理后,难免出现冗余数据和有效数据丢失的情况[25]。因此,还需要结合宏蛋白质组学、风味物质分析等其他技术,才能够更加全面地分析发酵食品中微生物的作用机制以及提升传统产业的现代化水平。
2.3 郫县豆瓣后发酵期细菌演替主坐标分析
由于不同后发酵期郫县豆瓣细菌群落构成复杂,差异较大。为反映长达6 a的郫县豆瓣后发酵“日晒夜露”过程中细菌群落的演替变化情况,本实验组对测序数据进行了基于unweighted unifrac的PcoA,既考虑细菌序列在后发酵期间的群落中出现过程,也考虑该相关序列的丰度变化[26]。具体的说,先根据最大近似法使用FastTree构建进化树,再利用Fastunifrac分析得到样品间距离矩阵,结果如图6所示 。
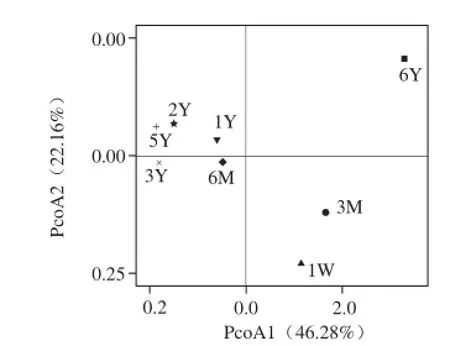
图 6 主坐标分析结果Fig. 6 Result of principal coordinate analysis
由图6可知,后发酵时间 对郫县豆瓣的细菌群落组成具有重要影响,大致可以分为3 个变化阶段,即是前3 个月、发酵6 个月至5 a和发酵6 a。同时,第2变化阶段又可以被分为2 个过程,即是发酵6 个月至1 a间的细菌群落,相比发酵2~5 a第1个阶段细菌群落多样性和丰度也存在明显差异。
目前,业内有一种片面的认识:认为后熟周期越长越好。因此 常出现由于销售制约而无限期延长后发酵周期的现状。已有学者指出,在现有的发酵条件下,后熟完成后,各种有效成分将会从高峰降至低谷,综合成分整体劣变,同时卫生指标和理化指标也会相应降低,因此应严格控制发酵周期为6 个月至1 a[1]。本研究结果发现后发酵期6 个月至5 a的郫县豆瓣细菌群落演替变化相较于其他阶段变化较小,为该观点从微生物角度提供了一定理论支撑。
3 结 论
本研究利用HTS高通量测序技术对郫县豆瓣后发酵过程中的细菌群落进行了分析研究,结果表明该技术可以较为有效、全面地评价郫县豆瓣中的细菌群落。本研究丰富了对传统本土调味品-郫县豆瓣中细菌的认识,细菌群落的多样性和丰度均显著高于同属东亚国家的韩国泡菜、辣酱等传统发酵调味品。发酵时间对郫县豆瓣后发酵期的细菌群落组成有重要影响,无限期延长后发酵周期会带来微生物群落丰度和多样性的迅速降低。
[1] 李幼筠. 郫县豆瓣剖析[J]. 中国酿造, 2008, 27(11): 83-86. DOI:10.3969/j.issn.0254-5071.2008.06.027.
[2] 余浪, 阚建全. 传统豆瓣的研究进展[J]. 中国调味品, 2008, 33(5): 26-31. DOI:10.3969/j.issn.1000-9973.2008.05.002.
[3] 杜敬秋, 刘浏. 607亿 元郫县豆瓣跻身中国品牌价值榜[N]. 成都日报, 2015-12-14(8).
[4] 石磊, 李诚. HACCP在郫县豆瓣生产中的应用[J]. 中国调味品, 2007, 32(6): 25-27. DOI:10.3969/j.issn.1000-9973.2007.06.002.
[5] 高岭. 郫县豆瓣的生产工艺改进[J]. 中国调味品, 2006, 31(5): 34-39. DOI:10.3969/j.issn.1000-9973.2006.05.009.
[6] 张琦, 汪先丁, 杨虎, 等. 郫县豆瓣自然发酵过程中细菌群落结构的变化[J]. 食品与发酵科技, 2010, 46(6): 16-18. DOI:10.3969/ j.issn.1674-506X.2010.06.005.
[7] 聂志强, 韩玥, 郑宇, 等. 宏基因组学技术分析传统食醋发酵过程微生物多样性[J]. 食品科学, 2013, 34(15): 198-203. DOI:10.7506/ spkx1002-6630-201315041.
[8] 左勇, 王小龙, 张晶, 等. 基于高通量测序技术的宜宾芽菜真菌多样性研究[J]. 中国调味品, 2016, 41(4): 19-22. DOI:10.3969/ j.issn.1000-9973.2016.04.004.
[9] 陈玲, 袁玉菊, 曾丽云, 等. 16S rDNA 克隆文库法与高通量测序法在浓香型大曲微生物群落结构分析中的对比研究[J]. 酿酒科技 , 2015(12): 33-40. DOI:10.13746/j.njkj.2015145.
[10] 韩 齐, 李媛媛, 孙方达, 等. 新一代测序技术在食品微生物学中的应用[J]. 食品工业, 2016, 37(1): 278-283.
[11] G UAN L, CHO K H, LEE J H. Analysis of the cultivable bacterial community in jeotgal, a Korean salted and fermented seafood, and identification of its dominant bacteria[J]. Food Microbiology, 2011, 28(1): 101-113. DOI:10.1016/j.fm.2010.09.001.
[12] JUNG J Y, LEE S H, JEON C O. Microbial community dynamics during fermentation of doenjang-meju, traditional Korean fermented soybean[J]. International Journal of Food Microbiology, 2014, 185: 112-120. DOI:10.1016/j.ijfoodmicro.2014.06.003.
[13] PARK E J, CHUN J, CHA C J, et al. Bacterial community analysis during fermentation of ten representative kinds of kimchi with barcoded pyrosequencing[J]. Food Microbiology, 2012, 30(1): 197-204. DOI:10.1016/j.fm.2011.10.011.
[14] LEE S H, JUNG J Y, JEON C O. Bacterial community dynamics and metabolite changes in myeolchi-aekjeot, a Korean traditional fermented fish sauce, during fermentation[J]. International Journal of Food Microbiology, 2015, 203: 15-22. DOI:10.1016/ j.ijfoodmicro.2015.02.031.
[15] NAM Y D, LEE S Y, LIM S I. Microbial community analysis of Korean soybean pastes by next-generation sequencing[J]. International Journal of Food Microbiology, 2012, 155(1/2): 36-42. DOI:10.1016/ j.ijfoodmicro.2012.01.013.
[16] KERGOURLAY G, TAMINIAU B, DAUBE G, et al. Metagenomic insights into the dynamics of microbial communities in food[J]. International Journal of Food Microbiology, 2015, 213: 31-39. DOI:10.1016/j.ijfoodmicro.2015.09.010.
[17] TAMAKI H, WRIGHT C L, LI X, et al. Analysis of 16S rRNA amplicon sequencing options on the Roche/454 next-generation titanium sequencing platform[J]. PLoS ONE, 2011, 6(9): e25263. DOI:10.1371/journal.pone.0025263.
[18] LI X Z, RUI J P, MAO Y J, et al. Dynamics of the bacterial community structure in the rhizosphere of a maize cultivar[J]. Soil Biology and Biochemistry, 2014, 68: 392-401. DOI:10.1016/j.soilbio.2013.10.017. [19] EDGAR R C, HAAS B J, CLEMENTE J C, et al. UCHIME improves sensitivity and speed of chimera detection[J]. Bioinformatics, 2011, 27(16): 2194-2200. DOI:10.1093/bioinformatics/btr381.
[20] WANG Q, GARRITY G M, TIEDJE J M, et al. Na☒ve bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy[J]. Applied and Environmental Microbiology, 2007, 73(16): 5261-5267. DOI:10.1128/aem.00062-07.
[21] 陈佐国, 叶振钟. 郫县-成都市的对外窗口 川西平原璀灿的明珠[J].中国西部, 1994(1): 26-27.
[22] 邹艳玲. 后熟期郫县豆瓣细菌多样性分析及产品护色研究[D]. 成都: 西华大学, 2013: 12-14.
[23] 赵辉平, 关统伟, 董丹, 等. 郫县豆瓣中可培养细菌多样性分析及酶活性初筛[J]. 中国调味品, 2015, 40(8): 5-9. DOI:10.3969/ j.issn.1000-9973.2015.08.002.
[24] 张大凤, 邹艳玲, 李明元. 郫县豆瓣生产用复合微生物菌剂的研制[J]. 中国调味品, 2012, 37(7): 62-64; 68. DOI:10.3969/ j.issn.1000-9973.2012 .07.017.
[25] GOODWIN S, McPHERSON J D, McCOMBIE W R. Coming of age: ten year s of next-generation sequencing technologies[J]. Nature Reviews Genetics, 2016, 17(6): 333-351. DOI:10.1038/nrg.2016.49. [26] NAM Y D, YI S H, LIM S I. Bacterial diversity of cheonggukjang, a traditional Korean fermented food, analyzed by barcoded pyrosequencing[J]. Food Control, 2012, 28(1): 135-142. DOI:10.1016/ j.foodcont.2012.04.028.
Bacterial Community Analysis of Pixian Soybean Paste during Post-Fermentation by High-Throughput Sequencing
ZHAO Hongyu1, XU Weizhen1, YANG Guohua2, LIU Yuanfu3, YUE Peng2, ZHANG Liang1,*
(1. Key Laboratory of Food Biotechnology of Sichuan Province, School of Food and Bioengineering , Xihua University, Chengdu 610039, China; 2. Soybean Paste Brewing Technology and Engineering Laboratory of Sichuan Province, Sichuan Dandan Pixian Soybean Paste Co. Ltd., Chengdu 611732, China; 3. Sichuan Youlian Condiment Food Co. Ltd., Chengdu 611732, China)
This study was undertaken to analyze the microbial communities in Pixian soybean paste during the postfermentation process by high-throughput sequencing method. The results revealed that at the taxonomic levels of microbes including phylum, class, order, family, and genus, high-throughput sequencing could detect a total of 57 phyla, 174 classes, 321 orders, 535 families and 921 genera. Post-fermentation time had a significant effect on the bacterial community composition of Pixian soybean paste. During post-fermentation, the quantities of Enterobacteriaceae and Streptophyta continued to decrease whereas Sphingomonadaceae, Bacillaceae, Clostridiaceae and Peptostreptococcaceae increased first and then decreased, most of which reached their peak values during 1 to 3 years. However, after 6 years of fermentation, the bacterial community of Pixian soybean paste was significantly lower than that of other samples. With this method, a large number of non-cultureable bacteria and unclassified bacteria were detected. The re sults of bacterial diversity analysis were close to the microecology of the samples, which were helpful to better understand the bacterial diversity of Pixian soybean paste and might provide a scientific support for the modernization of the traditional industry and the control of food quality and safety.
Pixian soybean paste; bacterial community; high-throughput sequencing
10.7506/spkx1002-6630-201710020
Q933
A
1002-6630(2017)10-0117-06
2016-08-02
四川省战略性新兴新产品重点研发项目(2015GZX0021);四川省重点研发项目(2016NZ0093);成都市科技惠民技术研发项目(2015-HM01-00003-SF);成都市农业技术成果应用示范项目(2015-NY01-00001-NC);成都市农业技术研发项目(2015-NY02-00097-NC);成都市产学研联合实验室项目(2015-YF04-00047-JH);西华大学食品生物技术重点实验室建设项目(川教2006-313)
赵红宇(1992—),男,硕士研究生,研究方向为食品生物技术。E-mail:redrain.z@foxmail.com
*通信作者:张良(1982—),男,副教授,博士,研究方向为食品生物技术。E-mail:zhang-liang@foxmail.com
赵红宇, 徐炜桢, 杨国华, 等. 基于高通量测序的郫县豆瓣后发酵期细菌多样性研究[J]. 食品科学, 2017, 38(10): 117-122. DOI:10.7506/spkx1002-6630-201710020. http://www.spkx.net.cn
ZHAO Hongyu, XU Weizhen, YANG Guohua, et al. Bacterial community analysis of pixian soybean paste during postfermentation by high-throughput sequencing[J]. Food Science, 2017, 38(10): 117-122. (in Chinese with English abstract)
10.7506/spkx1002-6630-201710020. http://w ww.spkx.net.cn
