抗脱氧雪腐镰刀菌烯醇纳米抗体随机突变与筛选
2017-06-05吴红静曹冬梅付金衡
吴红静,曹冬梅,许 杨,,付金衡,涂 追,*
抗脱氧雪腐镰刀菌烯醇纳米抗体随机突变与筛选
吴红静1,2,曹冬梅1,许 杨1,3,付金衡3,涂 追1,*
( 1.南昌大学 食品科学与技术国家重点实验室,江西 南昌 330047;2.南昌大学科学技术学院,江西 南昌 330029;3.南昌大学中德联合研究院,江西 南昌 330047)
采用易错聚合酶链式反应技术,对来源于天然纳米抗体文库的抗脱氧雪腐镰刀菌烯醇(deoxynivalenol,DON)纳米抗体进行随机突变,通过优化和比较突变条件,构建了突变分布均匀的突变文库。噬菌体展示技术对突变文库进行了6 轮筛选,获得了8 种突变序列,其检测信号为野生型2.1 倍。多序列比对及三维结构建模分析表明,突变序列的突变位点集中于互补决定区与框架区连接的区域,并且部分序列出现了可以形成二硫键的氨基酸残基。研究结果为抗DON纳米抗体进一步的深度突变和改造提供了线索,同时可为其他抗小分子纳米抗体的改造提供借鉴。
纳米抗体;脱氧雪腐镰刀菌烯醇;易错聚合酶链式反应;噬菌体展示
纳米抗体是由天然缺失轻链的重链抗体(heavychain antibodies,HCAbs)可变区构成,又称为VHH(variable domain of heavy chain of heavy-chain)抗体[1]。纳米抗体具有分子质量小、易于表达以及稳定性好等优点,现已广泛应用于生物医学研究、诊断及食品安全等领域[2-5]。近年来,在食品安全检测领域,纳米抗体已成功用于抗原模拟、传统抗体替代等方面的应用[6-9]。虽然如此,目前针对小分子的高亲和力纳米抗体报道较少,有学者指出基于纳米抗体本身的结构特点,动物免疫系统可能并不适合产生高亲和力的抗小分子纳米抗体[10]。因此,采用体外突变和分子改造技术可为提高纳米抗体的亲和力、稳定性等特性提供有效途径[11-14]。
脱氧雪腐镰刀菌烯醇(deoxynivalenol,DON)属于B类单端孢霉烯族真菌毒素,是一种由真菌产生的次级代谢产物[15]。前期研究从天然纳米抗体噬菌体展示文库中筛选获得了可特异结合DON的纳米抗体,命名为DONIV2[16]。本研究采用易错聚合酶链式反应(error prone polymerase chain reaction,epPCR)对DONIV2进行随机突变,通过噬菌体展示技术对突变文库进行淘选,比较突变序列氨基酸残基变化以及突变位点三维结构的位置,分析抗DON纳米抗体与DON相互作用的机制和特点,为定向改造研究提供线索。
1 材料与方法
1.1 材料
1.1.1 菌株与质粒
大肠杆菌(Escherichia coli)TG1,含有抗DON纳米抗体编码基因的质粒pRX2-DONIV2,噬菌粒pHEN1和辅助噬菌体KM13均为实验室保藏[16]。
1.1.2 试剂
限制性内切酶、Taq DNA聚合酶、TA克隆载体(pMD18-T)及各种DNA试剂盒 TaKaRa公司;T4连接酶 New England Biolab公司;酵母膏、蛋白胨英国Oxoid公司;辣根过氧化物酶偶联的抗M13单克隆抗体(HRP/Anti-M13 Monoclonal Conjugate) 美国GE Healthcare公司;氨苄青霉素、卡那霉素、四甲基联苯胺、牛血清白蛋白(bovine serum albumin,BSA)、卵清白蛋白(ovalbumin,OVA)、脱脂乳 上海生物工程有限公司;其他化学试剂均为国产分析纯。
1.1.3 引物
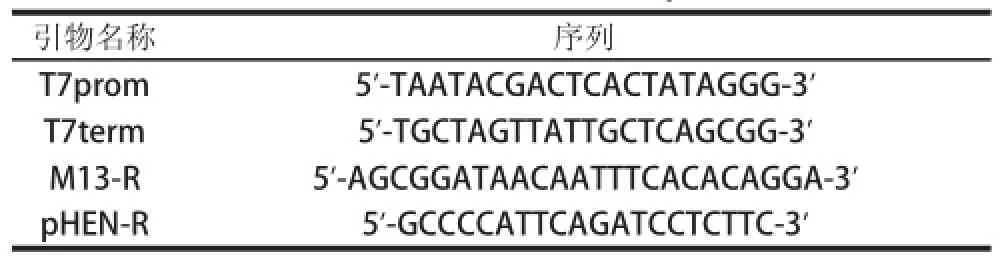
表 1 随机突变及鉴定所用的引物Table 1 Primers used for epPCR
随机突变以及突变文库鉴定所用的引物见表1。引物T7prom和T7term为载体pRX2的通用引物,分别位于抗DON纳米抗体编码基因两侧;引物M13和pHEN-R为载体pHEN1通用引物。PCR引物合成和DNA测序委托上海英俊生物技术有限公司。
1.2 方法
1.2.1 易错PCR
常规PCR体系(25 μL)中含有模板DNA(pRX2-DONIV2)10 ng、引物各10 pmol、10×PCR缓冲液(不含Mg2+)、TritonX-100 0.01%、MgCl21.5 mmol/L、dNTP各0.16 mmol/L、Taq聚合酶1.25 U。PCR扩增条件:94 ℃预变性3 min;94 ℃、30 s,60 ℃、30 s,72 ℃、1 min,10 个循环;94 ℃、30 s,55 ℃、30 s,72 ℃、1 min,25 个循环;72 ℃延伸10 min。
易错PCR体系在常规PCR体系的基础上进行,扩增条件相同,对Mg2+浓度、Mn2+浓度、TritonX-100浓度进行单因素试验。单因素优化后的条件下,设置Ⅰ和Ⅱ两组不同的dNTP浓度,Ⅰ组的4 种dNTP浓度均为0.16 mmol/L,Ⅱ组dATP和dGTP浓度均为0.16 mmol/L、dTTP和dCTP浓度均为1 mmol/L。
Ⅰ和Ⅱ两组易错PCR条件下的产物分别以1.2%的琼脂糖凝胶电泳分离,DNA凝胶回收试剂盒回收目的条带,然后按照TA克隆试剂盒说明,分别将两种纯化的易错PCR产物克隆到pMD18-T载体,氯化钙法转化大肠杆菌TG1感受态细胞,分别随机挑选10 个克隆进行测序。对两组易错PCR产物的突变位点、突变率以及突变类型等指标进行比较分析,选择适当突变率、突变分布均匀的易错PCR产物构建文库。
1.2.2 易错PCR文库的构建与鉴定
将易错PCR产物与噬菌粒pHEN1分别用限制性内切酶SfiⅠ和NotⅠ双酶切,经琼脂糖凝胶回收、定量后,以物质的量比3∶1,16 ℃条件下过夜连接。连接产物经乙醇沉淀后,溶于10 µL无菌水,电穿孔转化大肠杆菌TG1,感受态细胞的制备与转化,按照参考文献[18]进行。取10 µL电击、培养后的菌液倍比稀释,涂布LB(含100 μg/mL氨苄青霉素)培养板,计算库容。其余部分全部涂布于24 cm×24 cm LB(含100 μg/mL氨苄青霉素)培养板,37 ℃条件下倒置培养16 h。用5 mL LB培养基将培养板上的菌苔刮洗后,加入终含量25%甘油,分装,-80 ℃条件下保存备用。从计算库容的平板上随机挑取10 个克隆,采用通用引物M13-R和pHEN-R进行菌落PCR验证,计算有效库容量。
接种至少10 倍库容量的活细胞于5 mL的LB(含2%葡萄糖,100 μg/mL氨苄青霉素),30 ℃、220 r/min振摇培养至OD600nm达0.5,按感染复数20∶1加入辅助噬菌体,37℃、220 r/min振摇培养60 min。将培养物离心,用50 mL LB(含100 μg/mL 氨苄青霉素、50 μg/mL卡那霉素)重悬沉淀,30 ℃、220 r/min振摇培养过夜后,3 000×g离心取上清液,按标准的聚乙二醇/NaCl法纯化噬菌体[18],即得到噬菌体抗体文库,取10 μL测定滴度,其余分装于-80 ℃条件下保存备用。
1.2.3 噬菌体扩增、纯化和滴度测定
[18]进行测定。
1.2.4 易错PCR文库的淘选
以甲基化牛血清白蛋白(methylated BSA,MBSA)为载体,参考文献[17]制备DON人工抗原DON-MBSA。采用竞争洗脱法对突变文库进行淘选,方法参见文献[19],人工抗原DON-MBSA(100 μg/mL)包被酶标板,每轮3% BSA和3% OVA交替封闭,竞争洗脱采用50 ng/mL的DON溶液孵育1 h。
1.2.5 间接phage-ELISA
采用phage-ELISA法对淘选后的克隆进行亲和力鉴定[20]。酶标板中包被磷酸盐缓冲液(phosphate buffered saline,PBS)稀释的人工抗原DON-MBSA(1 μg/mL),3%脱脂乳封闭后,分别加入淘选后的噬菌体克隆和未突变的原始噬菌体克隆DONIV2,PBST(含0.5%吐温20的PBS溶液)洗板后,以HRP/Anti-M13(用3%脱脂乳,按1∶5 000(V/V)稀释)检测吸附的噬菌体。
1.2.6 三维结构模拟与对比
采用I-TASSER4.4软件建立纳米抗体的三维结构[21-22]。预测结果采用软件UCSF Chimera进行显示和编辑[23]。
2 结果与分析
2.1 随机突变文库的构建与鉴定
2.1.1 易错PCR条件的优化
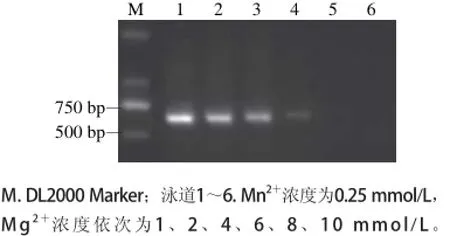
图 1 不同Mg2+浓度条件下易错PCR电泳图Fig. 1 Effects of Mg2+concentration on the epPCR
为了获得随机突变的编码基因,同时保证扩增产物特异性和产量,在普通PCR的基础上,首先对Mg2+、Mn2+浓度、TritonX-100浓度进行了单因素优化试验。如图1所示,随着Mg2+浓度的增加,扩增产物浓度下降,当Mg2+浓度大于6 mmol/L时,凝胶电泳未检测到扩增产物。高浓度的Mg2+能稳定错配的碱基对,因此选择较高的Mg2+浓度6 mmol/L。

图 2 不同Mn2+浓度条件下易错PCR电泳图Fig. 2 Effects of Mn2+concentration on the epPCR
反应体系中加入Mn2+可以降低聚合酶对模板的特异性,提高错配率。Mn2+浓度优化结果如图2所示,当Mn2+浓度降至0.2 mmol/L时,凝胶电泳出现特异性目的条带,选择Mn2+适宜浓度为0.2 mmol/L。

图 3 不同TritonX-100体积分数条件下易错PCR电泳图Fig. 3 Effects of TritonX-100 concentration on the epPCR
反应体系中TritonX-100体积分数对易错PCR的影响结果如图3所示,PCR产物的量随着TritonX-100体积的提高而增加,当体积分数达到0.04%时,PCR产物产量最大(泳道4)。
2.1.2 易错PCR产物突变分析
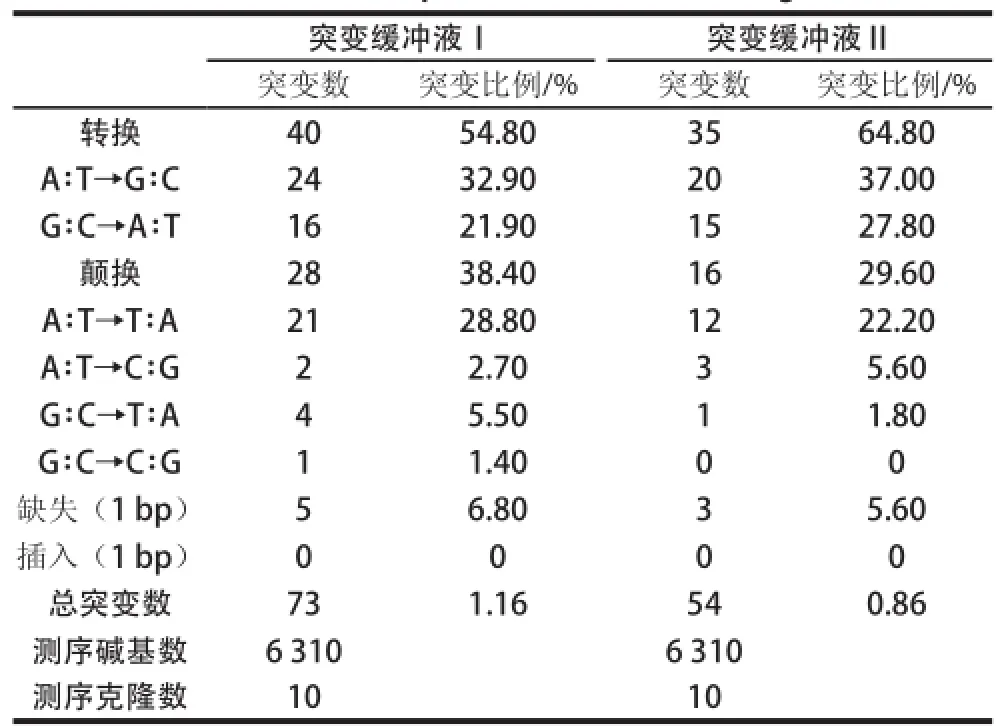
表 2 突变产物序列分析Table 2 Mutation spectrum of random mutagenesis
分别随机挑取10 个两组不同dNTP组分条件下的易错PCR产物进行序列分析,突变条件Ⅰ的总突变率为1.16%,突变条件Ⅱ的总突变率为0.86%,发生的突变绝大多数为点突变,缺失和插入较少(表2)。突变条件Ⅱ的转换比例(A∶T→G∶C/G∶C→A∶T)较突变条件Ⅰ小,即转换的偏好性更低,同时条件Ⅱ产物突变点的分布比条件Ⅰ突变点的分布更为均匀,综合考虑,采用突变条件Ⅱ的产物进行文库构建,平均每个克隆突变3~4 个核苷酸位点。
2.2 随机突变文库的构建、淘选与鉴定
2.2.1 文库的构建与淘选
将易错PCR产物用SfiⅠ和NotⅠ双酶切,凝胶电泳回收纯化,连接至噬菌粒载体pHEN1,连接产物电转化大肠杆菌TG1感受态细胞,菌落计数结果显示,获得约6×106个独立克隆,辅助噬菌体救援后得到噬菌体展示文库滴度约为2.7×1013CFU/mL。
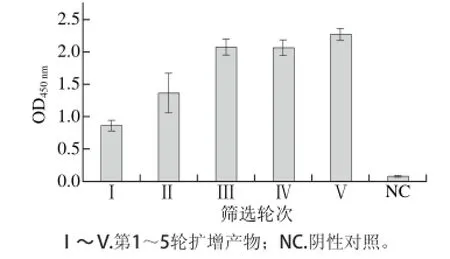
图 4 间接phage-ELISA测定每轮扩增产物Fig. 4 Indirect phage-ELISA of amplified phages from each round of panning
采用固相筛选的方法,对随机突变文库进行了6 轮筛选,phage-ELISA检测扩增后的每轮竞争洗脱物,结果显示在同样的实验条件下,OD450nm随着淘选的进行而升高,提示亲和力提高的突变得到了富集(图4)。
2.2.2 phage-ELISA鉴定阳性克隆

图 5 阳性克隆多序列比对结果Fig. 5 Multi-sequence alignment of mutants
从第6轮淘选竞争洗脱产物测定滴度的平板上,随机挑取96 个克隆,经辅助噬菌体救援后,进行phage-ELISA检测。以未突变的野生型克隆DONIV2作为对照,设置OD450nm为对照的2.1 倍为阳性标准,测定的96 个克隆中,共有30 个克隆为阳性,阳性率达31.3%。阳性克隆测序分析结果显示,共获得了8 条不同的突变序列(图5)。
2.3 三维结构模拟与分析
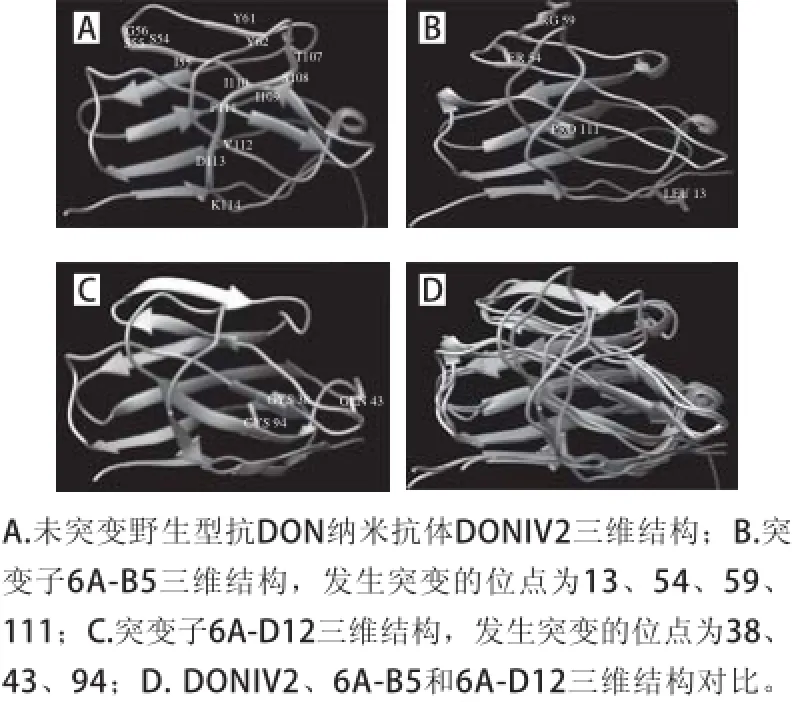
图 6 抗DON纳米抗体三维结构模拟与对比Fig. 6 Three-dimensional structure modeling
采用蛋白建模软件I-TASSER4.4,对突变前野生型抗DON纳米抗体DONIV2和2 个出现频率最高的突变子6A-B5和6A-D12进行三维结构建模,结果如图6A~C所示。通过结构对比发现,突变子的CDR3区域的结构与突变前的CDR3区域结构差异较大,其他部分主链结构基本一致(图6D)。
3 结 论
突变方法及反应条件对目的序列的突变率、突变位点等结果具有明显的影响。Rasila等[24]对易错PCR、化学突变和高突变菌株3 种突变方法的效果进行了详细的比较研究,发现易错PCR的突变率高和突变范围更广。本研究对易错PCR突变的条件进行了优化和比较,选择突变率适宜、突变位点分布均匀的突变条件,为鉴定影响结合活性的关键区域提供了基础。
通过比较突变前后氨基酸序列的变化,发现突变位点明显集中在CDR与FR连接的区域(图5),而CDR形成loop结构的位置发生突变较少,说明抗原结合表位可能直接由CDR-loop组成,这与分子对接结果吻合[25]。Min等[12]通过酵母展示系统对抗黄曲霉毒素B1的亲和力成熟的研究中,也发现大多数发生突变的位点不在CDR区域。值得注意的是,部分突变子(6A-D12和6A-H8)同时突变得到2 个半胱氨酸残基,三维结构模型显示,2 个半胱氨酸残基空间位置相邻(图6C),可形成新的二硫键,稳定蛋白质的结构,提示在蛋白质的设计和改造中,引入二硫键有利于稳定分子结构。
参考文献:
[1] MUYLDERMANS S. Nanobodies: natural single-domain antibodies[J]. Annual Review of Biochemistry, 2013, 82(2): 775-797. DOI:10.1146/annurev-biochem-063011-092449.
[2] DMITRIEV O Y, LUTSENKO S, MUYLDERMANS S. Nanobodies as probes for protein dynamics in vitro and in cells[J]. Journal of Biological Chemistry, 2016, 291(8): 3767-3775. DOI:10.1074/jbc. R115.679811.
[3] HELMA J, CARDOSO M C, MUYLDERMANS S, et al. Nanobodies and recombinant binders in cell biology[J]. Journal of Cell Biology, 2015, 209(5): 633-644. DOI:10.1083/jcb.201409074.
[4] de MEYER T, MUYLDERMANS S, DEPICKER A. Nanobody-based products as research and diagnostic tools[J]. Trends in Biotechnology, 2014, 32(5): 263-270. DOI:10.1016/j.tibtech.2014.03.001.
[5] TU Z, CHEN Q, LI Y P, et al. Identification and characterization of species-specific nanobodies for the detection of Listeria monocytogenes in milk[J]. Analytical Biochemistry, 2016, 493: 1-7. DOI:10.1016/j.ab.2015.09.023.
[6] XU Y, XIONG L, LI Y P, et al. Anti-idiotypic nanobody as citrinin mimotope from a naive alpaca heavy chain single domain antibody library[J]. Analytical and Bioanalytical Chemistry, 2015, 407(18): 5333-5341. DOI:10.1007/s00216-015-8693-3.
[7] SHU M, XU Y, WANG D, et al. Anti-idiotypic nanobody: a strategy for development of sensitive and green immunoassay for fumonisin B1[J]. Talanta, 2015, 143: 388-393. DOI:10.1016/ j.talanta.2015.05.010.
[8] QIU Y L, HE Q H, XU Y, et al. Deoxynivalenol-mimic nanobody isolated from a naive phage display nanobody library and its application in immunoassay[J]. Analytica Chimica Acta, 2015, 887: 201-208. DOI:10.1016/j.aca.2015.06.033.
[9] LIU X, XU Y, WAN D B, et al. Development of a nanobody-alkaline phosphatase fusion protein and its application in a highly sensitive direct competitive fluorescence enzyme immunoassay for detection of ochratoxin A in cereal[J]. Analytical Chemistry, 2015, 87(2): 1387-1394. DOI:10.1021/ac504305z.
[10] MAKVANDI-NEJAD S, FJALLMAN T, ARBABI-GHAHROUDI M, et al. Selection and expression of recombinant single domain antibodies from a hyper-immunized library against the hapten azoxystrobin[J]. Journal of Immunological Methods, 2011, 373(1/2): 8-18. DOI:10.1016/j.jim.2011.07.006.
[11] AKAZAWA-OGAWA Y, UEGAKI K, HAGIHARA Y. The role of intra-domain disulfide bonds in heat-induced irreversible denaturation of camelid single domain VHH antibodies[J]. Journal of Biochemistry, 2016, 159(1): 111-121. DOI:10.1093/jb/mvv082.
[12] MIN W K, KIM S G, SEO J H. Affinity maturation of singlechain variable fragment specific for aflatoxin B1using yeast surface display[J]. Food Chemistry, 2015, 188: 604-611. DOI:10.1016/ j.foodchem.2015.04.117.
[13] LIU J L, GOLDMAN E R, ZABETAKIS D, et al. Enhanced production of a single domain antibody with an engineered stabilizing extra disulfide bond[J]. Microbial Cell Factories, 2015, 14: 158. DOI:10.1186/s12934-015-0340-3.
[14] TURNER K B, ZABETAKIS D, GOLDMAN E R, et al. Enhanced stabilization of a stable single domain antibody for SEB toxin by random mutagenesis and stringent selection[J]. Protein Engineering Design & Selection, 2014, 27(3): 89-95. DOI:10.1093/protein/gzu001. [15] HASSAN Y I, WATTS C, LI X Z, et al. A novel peptide-binding motifs inference approach to understand deoxynivalenol molecular toxicity[J]. Toxins (Basel), 2015, 7(6): 1989-2005. DOI:10.3390/ toxins7061989.
[16] 涂追, 许杨, 刘夏, 等. 驼源天然单域重链抗体库的构建与鉴定[J]. 中国生物工程杂志, 2011, 31(4): 31-36. DOI:10.13523/ j.cb.20110406.
[17] 邓舜洲, 游淑珠, 许杨. 脱氧雪腐镰刀菌烯醇人工抗原的研制[J]. 食品科学, 2007, 28(2): 149-152. DOI:10.3321/ j.issn:1002-6630.2007.02.036.
[18] SAMBROOK J, RUSSELL D W. 分子克隆实验指南[M]. 黄培堂, 译. 3版. 北京: 科学出版社, 2008: 251-269.
[19] 刘夏, 许杨, 涂追, 等. 抗AFB1单域重链抗体文库淘选方法的研究[J]. 食品与生物技术学报, 2011, 30(6): 950-955. DOI:10.3321/ j.issn:1673-1689.2011.06.028.
[20] TU Z, XU Y, HE Q H, et al. Isolation and characterisation of deoxynivalenol affinity binders from a phage display library based on single-domain camelid heavy chain antibodies (VHHs)[J]. Food and Agricultural Immunology, 2012, 23(2): 123-131. DOI:10.1080/095401 05.2011.606560.
[21] YANG J Y, YAN R X, ROY A, et al. The I-TASSER Suite: protein structure and function prediction[J]. Nature Methods, 2015, 12(1): 7-8. DOI:10.1038/nmeth.3213.
[22] ROY A, KUCUKURAL A, ZHANG Y. I-TASSER: a unified platform for automated protein structure and function prediction[J]. Nature Protocols, 2010, 5(4): 725-738. DOI:10.1038/nprot.2010.5.
[23] PETTERSEN E F, GODDARD T D, HUANG C C, et al. UCSF Chimera-a visualization system for exploratory research and analysis[J]. Journal of Computational Chemistry, 2004, 25(13): 1605-1612. DOI:10.1002/jcc.20084.
[24] RASILA T S, PAJUNEN M I, SAVILAHTI H. Critical evaluation of random mutagenesis by error-prone polymerase chain reaction protocols, Escherichia coli mutator strain, and hydroxylamine treatment[J]. Analytical Biochemistry, 2009, 388(1): 71-80. DOI:10.1016/j.ab.2009.02.008.
[25] 涂追, 许杨, 付金衡, 等. 抗DON单域重链抗体序列分析及三维建模与对接[J]. 江苏农业学报, 2011, 27(4): 893-898. DOI:10.3969/ j.issn.1000-4440.2011.04.036.
Random Mutation and Mutant Screening of Anti-Deoxynivalenol Nanobody
WU Hongjing1,2, CAO Dongmei1, XU Yang1,3, FU Jinheng3, TU Zhui1,*
(1. State Key Laboratory of Food Science and Technology, Nanchang University, Nanchang 330047, China; 2. College of Science and Technology, Nanchang University, Nanchang 330029, China; 3. Jiangxi-OAI Joint Research Institution, Nanchang University, Nanchang 330047, China)
In this work, an anti-deoxynivalenol nanobody isolated from a naive phage display library was adopted to investigate the relationship between bioactivity and the composition of amino acid residues using error prone polymerase chain reaction (epPCR) and phage display. The epPCR was firstly optimized to produce a phage display library with balanced distribution of mutation. After six rounds of panning of the mutant library, 30 out of 96 picked clones exhibited 8 different mutations, whose signal was 2.1 folds higher than that of the wild-type clone. Multi-sequence alignment analysis showed that mutant sites were prone to be found in the junction region between complementarity determining regions (CDRs) and framework regions (FRs). Three-dimensional structure modeling results indicated that some mutants were capable of forming an intramolecular disulphide bond, which benefits to stabilize the molecular structure. These data can provide a clue for further mutation and modification of anti-deoxynivalenol nanobody and also provide a guideline for the design of nanobody recognizing other low molecular weight compounds.
nanobody; deoxynivalenol; error prone PCR; phage display
10.7506/spkx1002-6630-201710001
TS201.6
A
1002-6630(2017)10-0001-05
吴红静, 曹冬梅, 许杨, 等. 抗脱氧雪腐镰刀菌烯醇纳米抗体随机突变与筛选[J]. 食品科学, 2017, 38(10): 1-5. DOI:10.7506/spkx1002-6630-201710001. http://www.spkx.net.cn
WU Hongjing, CAO Dongmei, XU Yang, et al. Random mutation and mutant screening of anti-deoxynivalenol nanobody[J]. Food Science, 2017, 38(10): 1-5. (in Chinese with English abstract)
10.7506/spkx1002-6630-201710001. http://www.spkx.net.cn
2016-06-12
国家自然科学基金青年科学基金项目(31301479);江西省教育厅科学技术研究项目(GJJ151502);南昌大学科学技术学院自然科学基金项目(2013-ZR-05)
吴红静(1981—),女,讲师,硕士,研究方向为食品安全与营养。E-mail:51531843@qq.com
*通信作者:涂追(1982—),男,副研究员,博士,研究方向为食品安全与生物技术。E-mail:tuzhui@ncu.edu.cn
