Pseudoalteromonas carrageenovora芳香基硫酸酯酶突变文库热稳定性提高突变体的筛选及鉴定
2017-06-05乔超超王新侠李鹤宾肖安风朱艳冰
乔超超,王新侠,李鹤宾,倪 辉,肖安风,朱艳冰,*
(1.集美大学食品与生物工程学院,福建 厦门 361021;2.厦门医学院药学系,福建 厦门 361023)
Pseudoalteromonas carrageenovora芳香基硫酸酯酶突变文库热稳定性提高突变体的筛选及鉴定
乔超超1,王新侠1,李鹤宾2,倪 辉1,肖安风1,朱艳冰1,*
(1.集美大学食品与生物工程学院,福建 厦门 361021;2.厦门医学院药学系,福建 厦门 361023)
利用易错聚合酶链式反应技术引入随机诱变,构建一个Pseudoalteromonas carrageenovora芳香基硫酸酯酶突变体库。经过筛选,获得一个芳香基硫酸酯酶热稳定性提高的突变株4-153。序列分析表明,该突变体有2 个氨基酸替换,包括D84A和H260L。以对硝基苯硫酸钾为底物,突变酶4-153(M4-153)的最适反应温度为55 ℃,在45、50、55、60 ℃处理30 min后,M4-153分别保留85%、83%、48%和13%的残留酶活力。野生型酶(WT)在45、50、55、60 ℃处理30 min后,分别保留79%、68%、21%和1%的残留酶活力。M4-153与WT相比具有更好的热稳定性。M4-153的最适反应pH值为8.0,在pH 5.0~9.0范围内保持稳定。EDTA对突变酶的抑制作用表明,金属离子在突变酶的催化过程中起重要作用。M4-153对一些洗涤剂,包括Triton X-100、Tween 20、Tween 80和Chaps,有好的耐受性。M4-153对龙须菜粗多糖硫酸基团的脱硫率为79.5%。
芳香基硫酸酯酶;易错聚合酶链式反应;热稳定性提高;突变体性质
琼脂是构成红藻细胞壁的主要结构多糖,由琼脂糖和硫琼胶组成[1]。琼脂糖是由(1-3)-β-D-半乳糖和(1-4)-3,6-内醚-α-L-半乳糖相互交替连接而成的长链结构[1]。硫琼胶的结构与琼脂糖基本相同,主要差别是替换了半乳糖残基C6位置上的羟基,以硫酸基、甲氧基等基团进行替代[1]。琼脂的凝胶过程主要是由糖残基的相互交联作用所导致,交联越紧密,形成的琼脂凝胶强度就越大,而硫琼胶上具有的硫酸基等基团会影响分子间的相互交联作用,使得琼脂凝胶强度降低[2],琼胶产品质量下降。在琼脂的工业生产中,脱除硫酸基团是很重要的环节,最为广泛的方法为碱法[3]。国内外常用的碱法可分为3 种类型:低温浓碱法、常温浓碱法和高温稀碱法[4]。这类方法工艺简单、操作方便,但是由于处理过程中大量强碱的使用,使得该方法有反应难控制、产品得率低、胶质易损失和环境污染严重等缺陷。很多研究者开始尝试探索新的有效方法。芳香基硫酸酯酶能催化芳香基硫酸酯键的水解,生成芳基化合物和无机硫酸盐[5]。芳香基硫酸酯酶具有广泛的分布性,在真菌[6]、细菌[7]、海胆[8]、海藻[9]、蜗牛[10]、哺乳动物[11]和人体[12]中都有分离得到。其中对一些微生物来源的芳香基硫酸酯酶,这包括从米曲霉[13]、肺炎克雷伯杆菌[14]、肺炎克雷伯菌[15]、海单胞菌[16]、交替假单孢菌[17]、铜绿假单胞菌[7,18]、鼠伤寒沙门氏菌[19]、黏质沙雷氏菌[20]、鞘氨醇单胞菌[21]、链霉菌[22]和海栖热袍菌[23]中得到的芳基硫酸酯酶进行了研究,并发现某些芳香基硫酸酯酶具有可裂解琼脂上硫酸酯键的功能[5,16-17]。利用酶法去除硫酸基团具有反应条件温和、易控制、硫酸基团脱除率高,环境污染小等优点。芳香基硫酸酯酶在琼脂提取工业中具有潜在的应用价值[24]。
来自Pseudoalteromonas carrageenovora的芳香基硫酸酯酶对对硝基苯硫酸钾(potassium 4-nitrophenyl sulfate,p-NPS)有活性,以及有脱除琼脂硫酸基团的能力[17]。从P. carrageenovora分离得到的芳香基硫酸酯酶基因(984 bp)克隆并在大肠杆菌中表达,发现45 ℃以上重组酶热稳定性不高。本研究以易错聚合酶链式反应(polymerase chain reaction,PCR)为基础定向进化提高该酶的热稳定性,并研究突变酶的酶学性质。
1 材料与方法
1.1 材料与试剂
E. coli BL21(DE3)、pET-28a质粒由集美大学食品与生物工程学院发酵工程实验室保藏。
质粒提取试剂盒、DNA凝胶回收试剂盒 天根生化科技(北京)有限公司;限制性内切酶BamHⅠ和HindⅢ、T4 DNA连接酶、rTaq酶、dNTPs TaKaRa公司;p-NPS 美国Sigma公司。
1.2 仪器与设备
双层全温度恒温摇床 上海智城分析仪器制造有限公司;双人单面净化工作台 苏州净化设备有限公司;酶标仪 美国BioTek公司;超声细胞破碎仪 美国Sonics公司;数显恒温水浴锅 常州国华电器有限公司;蛋白电泳仪 美国Bio-Rad公司;小型高速常温离心机、蛋白核酸测定仪 德国Eppendorf公司;大型高速冷冻离心机 美国Beckman公司;离子色谱 美国Dionex公司;凝胶成像系统 美国GE公司;数控超声波清洗器 昆山市超声仪器有限公司。
1.3 方法
1.3.1 芳香基硫酸酯酶随机突变体库的构建
P. carrageenovora芳香基硫酸酯酶含有信号肽,其长度为18 个氨基酸。在前期研究中,构建了不含信号肽的芳香基硫酸酯酶基因(930 bp)重组质粒pET-28a-ars。随机突变体库是基于易错PCR构建。以重组质粒pET-28aars为模板,使用上游引物ars-F(5’-CGCGGATCCTTT ACGTTTAACGGCAGC-3’)和下游引物ars-R(5’-CCC AAGCTTGCGTTTTAGTTCGTAAC-3’)(划线部分分别为BamHⅠ和HindⅢ的酶切位点)对目的基因进行扩增。总反应体系为50 μL,其中包含5 μL 10×缓冲液、0.2 μmol/L引物、0.5 mmol/L dTTP、0.5 mmol/L dGTP、0.1 mmol/L dATP、0.1 mmol/L dCTP、1 U rTaq 聚合酶、7 mmol/L MgCl2、2 ng重组质粒模板。热循环参数为:95 ℃ 5 min;94 ℃ 45 s,50 ℃ 45 s,72 ℃ 1 min(35 次);72 ℃ 10 min。BamHⅠ和HindⅢ消化后,PCR产物插入pET-28a(+)载体,构建重组质粒突变文库。质粒转化大肠杆菌BL21(DE3),生成突变文库。
1.3.2 热稳定性提高突变株的筛选
将文库的突变子涂布于LB固体培养基(含50 mg/mL卡那霉素和0.05 mmol/mL异丙基-β-D-硫代半乳糖苷(isopropyl β-D-1-thiogalactopyranoside,IPTG),37 ℃过夜培养后,根据产芳香基硫酸酯酶菌株水解底物p-NPS的显色反应来进行突变文库的初筛。含有菌株的平板经过50 ℃处理2 h后,用含20 mmol/L p-NPS的软琼脂覆盖,这时野生型菌株无显色反应,具有显色反应的菌落认为是热稳定性提高的突变株。菌株过夜培养后,按1∶100(V/V)转接到5 mL LB液体培养基(含50 mg/mL卡那霉素)中,37 ℃、180 r/min培养至OD600nm达到0.6,加入终浓度为0.05 mmol/mL IPTG,25 ℃、180 r/min培养10 h。离心收集菌体,重悬于50 mmol/L Tris-HCl缓冲液(pH 7.5)中,在冰浴条件下进行超声波破碎处理,10 000 r/min离心20 min获得的上清液即为粗酶液。粗酶液在50 ℃处理2 h后,按下述1.3.5节的方法检测酶的残余活力,进行复筛验证。复筛后得到的热稳定性提高突变株,送至铂尚生物技术(上海)有限公司进行基因测序分析。
1.3.3 重组酶的表达与纯化
突变型和野生型菌株分别接种至200 mL含50 μg/mL卡那霉素的LB培养基中,37 ℃摇动培养至OD600nm达到0.5,然后加入IPTG至终浓度为0.05 mmol/L。在25 ℃孵育10 h后,诱导细胞通过6 000 r/min离心5 min收获。纯化带有His标签的蛋白质,使用Ni Sepharose 6 Fast Flow在自然条件下进行亲和层析。纯化后的蛋白用Bradford法测定蛋白质浓度[25],十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)分析纯化的酶,并对其分子质量进行分析。
1.3.4 芳香基硫酸酯酶的活性测定
根据Kim等[17]的方法稍作修改后,测定芳香基硫酸酯酶的活性(文中进行酶学性质分析使用的酶为纯化后的酶,除非另有说明)。取80 μL用50 mmol/L Tris-HCl缓冲液(野生型使用pH 7.5,突变体使用pH 8.0)配制的20 mmol/L p-NPS底物溶液,加入20 μL酶液(400 ng),55 ℃孵育10 min后,加入25 μL 5 mol/L NaOH溶液终止反应。在410 nm波长处使用分光光度计测定吸光度。在此条件下,每分钟催化生成1 μmoL对硝基苯酚所需的酶量定义为一个酶活力单位(U)。
1.3.5 反应温度对酶活性的影响
突变酶的最适反应温度在30~80 ℃的范围内测定。突变型与野生型酶的热稳定性在45、50、55、60 ℃下进行。将酶在不同温度条件下处理30 min后,测定酶的残余活力,以未经处理的酶活力作为100%。
1.3.6 pH值对突变酶活性的影响
将酶在pH3.0~10.0的缓冲液中于37 ℃放置 1 h后,测定酶的残余活力。以未经处理的酶活力定义为100%。测定使用的缓冲液为:50 mmol/L 磷酸二氢钠-柠檬酸(pH 3.0~5.0)、50 mmol/L柠檬酸-柠檬酸钠缓冲液(pH 5.0~7.0),50 mmol/L Tris-HCl缓冲液(pH 7.0~9.0),50 mmol/L甘氨酸-氢氧化钠缓冲液(pH 9.0~10.0)。
1.3.7 金属离子对突变酶活性的影响
在突变酶中分别添加终浓度为1 mmol/L或10 mmol/L的不同金属盐离子,包括NaCl、KCl、CaCl2、MgCl2、ZnCl2、CuCl2、MnCl2、CoCl2和CdCl2。37 ℃条件下放置1 h后,测定酶的残余活力,研究金属离子对突变酶活性的影响。以未添加金属离子的酶活力为100%。
1.3.8 抑制剂和洗涤剂对突变酶活性的影响
在突变型芳香基硫酸酯酶中分别添加终浓度为1 mmol/L或10 mmol/L的抑制剂(包括乙二胺四乙酸(ethylenediaminetetraacetic acid,EDTA)、β-巯基乙醇(β-mercaptoethano,β-ME)、二硫苏糖醇(DL-dithiothreitol,DTT)和苯甲基磺酰氟(phenylmethanesulfonyluoride,PMSF))、0.1%或1%去垢剂(包括Triton X-100、Tween 20、Tween 80、Chaps和SDS)。置于37 ℃条件下放置1 h后,测定酶的残余活力,研究抑制剂和洗涤剂对突变酶活性的影响。以未添加抑制剂或去垢剂的酶活力为100%。
1.3.9 动力学参数的测定
400 ng突变酶与不同浓度(0.1~3.0 mmol/L)p-NPS在最适条件下反应10 min后,测定酶的活力。用Lineweaver-Burk双倒数作图,得到线性回归方程,并分析计算酶的动力学参数米氏常数(Km)和最大反应速率(vmax)。
1.3.10 脱除龙须菜粗多糖硫酸基团的研究
称取0.1 g龙须菜粗多糖溶解于20 mL 50 mmol/L Tris-HCl(pH 7.5)缓冲液中。分别加入60 U和90 U的野生型或突变型酶,45 ℃条件下处理4 h。将处理后的混合溶液置于尼龙布上,用超纯水充分洗涤后,烘干粉碎。称取等质量的经过酶处理后与未处理的龙须菜粗多糖,高温碳化后,置于马弗炉中550 ℃灰化4 h,将所得灰分全部溶于超纯水中,定容至25 mL。取1 mL溶液用0.22 μm的膜过滤后,利用ICS-2100离子色谱测定样品的硫酸基团含量。以未经酶处理的粗多糖为对照,根据以下公式计算脱硫率:

2 结果与分析
2.1 芳香基硫酸酯酶随机突变体库的构建与筛选
芳香基硫酸酯酶基因利用易错PCR技术引入随机诱变,库容量约为2.0×105。经过两步筛选,发现突变体4-153与野生型相比热稳定性提高。基因序列分析发现,野生型与突变型酶基因相比有3 个碱基被替换,导致4-153有2 个氨基酸残基发生改变,分别为D84A和H260L(野生型和突变型芳香基硫酸酯酶4-153分别命名为WT和M4-153)。突变酶4-153被选定做进一步的酶学性质研究。
2.2 突变型和野生型酶的表达与纯化
经过诱导表达后,分别对重组的WT和M4-153进行纯化。对纯化后的蛋白质进行SDS-PAGE分析显示,突变后的芳香基硫酸酯酶大小与野生型一致,并测得WT和M4-153的酶活力分别为11.3、7.8 U/mg。
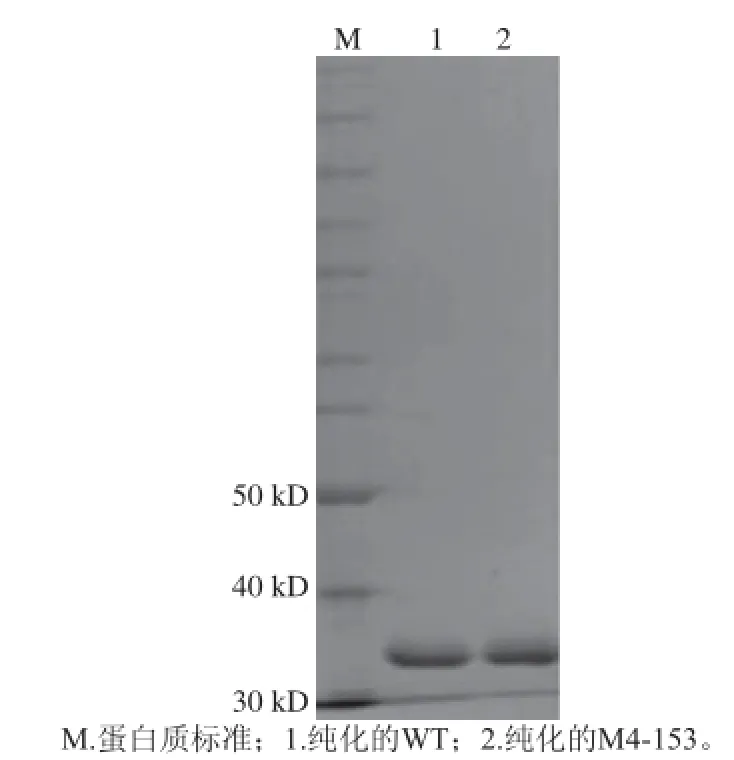
图 1 芳香基硫酸酯酶在大肠杆菌中的表达Fig. 1 Expression of the arylsulfatase genes in E. coli
2.3 温度对芳香基硫酸酯酶的影响
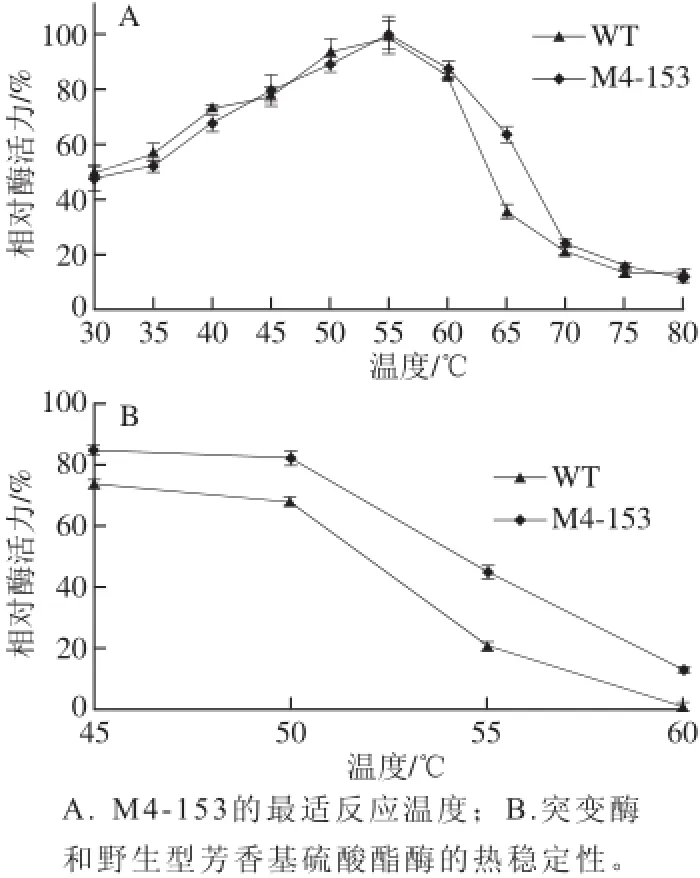
图 2 温度对M4-153和WT的影响Fig. 2 Effect of temperature on the activity of M4-153 and WT
M4-153的最适温度在30~80 ℃内进行测定。如图2所示,M4-153和WT的最适反应温度为55 ℃,在45~60 ℃温度范围内仍可以保持超过80%活性(图2A)。热稳定性分析显示,在45、50、55、60 ℃条件下处理30 min后,M4-153分别保留了85%、83%、48%、13%的残余活性,而WT在45、50、55、60 ℃处理30 min后,分别保留了79%、68%、21%、1%的残余活性(图2B)。以上结果表明,突变酶M4-153比野生型酶WT具有更好的热稳定性。
2.4 pH值对突变酶活性的影响
芳香基硫酸酯酶M4-153和WT的最适pH值在5.0~10.0的范围内测定,结果如图3A所示,M4-153的最适pH值为8.0,WT的最适pH值为7.5。当pH值低于6.5或高于9.5,M4-153和WT的酶活性急剧降低。pH值稳定性实验结果表明,M4-153在pH 5.0~9.0范围内保持稳定,在37 ℃放置1 h后仍能保留其原活性的60%以上,而WT在pH 6.0~9.0范围内保持稳定,在37 ℃放置1 h后仍能保留其原活性的60%以上(图3B)。
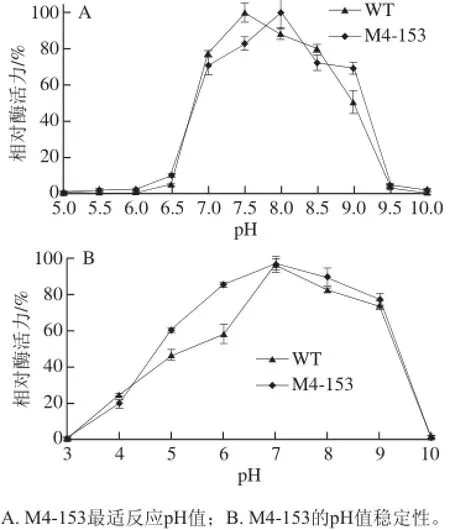
图 3 pH值对M4-153和WT活性和稳定性的影响Fig. 3 Effect of pH on the activity and stability of M4-153 and WT
2.5 金属离子对突变酶活性的影响
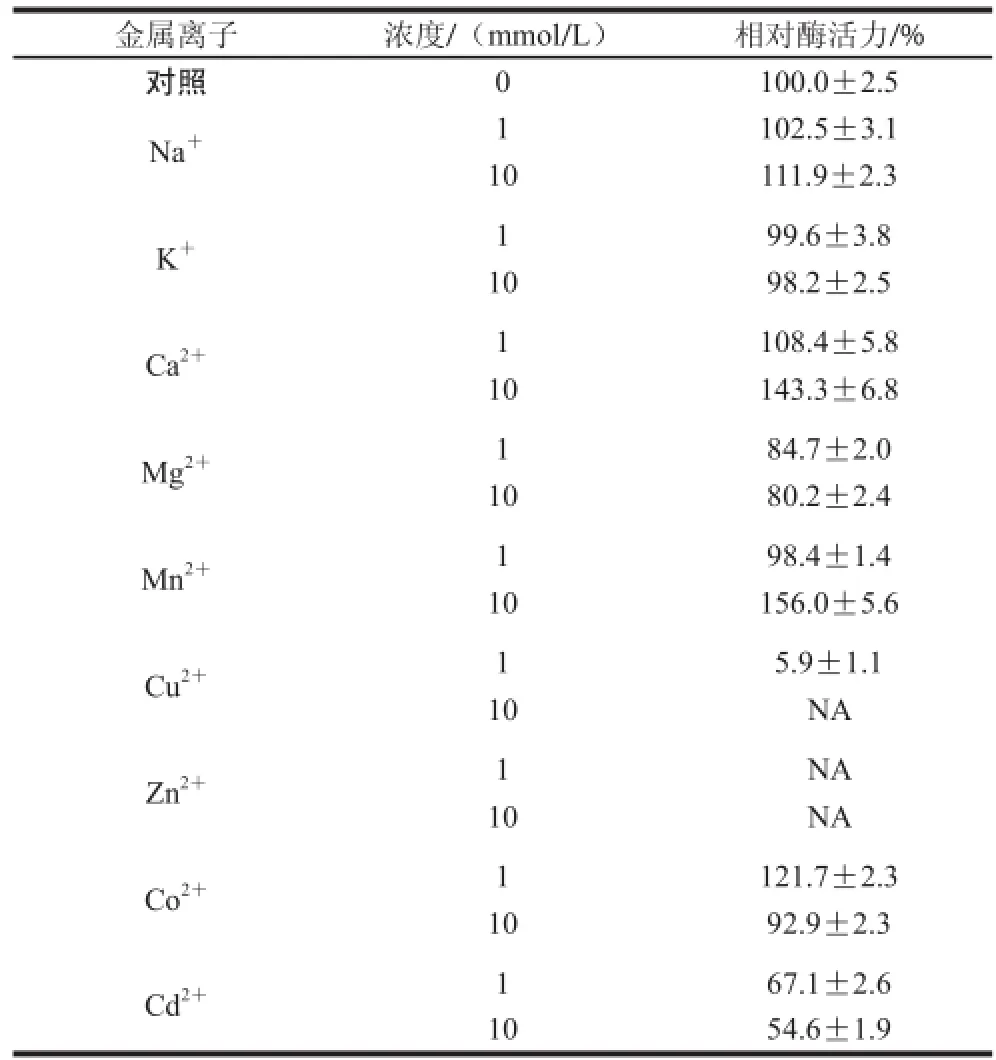
表 1 金属离子对突变酶活性的影响Table 1 Effects of metal ions on mutant arylsulfatase activity
由表1可知,K+对突变芳香基硫酸酯酶活力没有显著的影响,而Mg2+、Cu2+、Zn2+、Cd2+会抑制酶的活性,浓度越高抑制越明显,Zn2+和高浓度的Cu2+会导致酶完全失活。Ca2+对突变酶的活力有一定的促进作用。1 mmol/L的Na+和Mn2+对酶活力没有影响,而10 mmol/L的Na+、和Mn2+对酶活力具有促进作用。Co2+在1 mmol/L对酶活性有促进作用,当浓度增大到10 mmol/L时会稍微抑制酶活力。
2.6 抑制剂和去垢剂对突变酶活性的影响
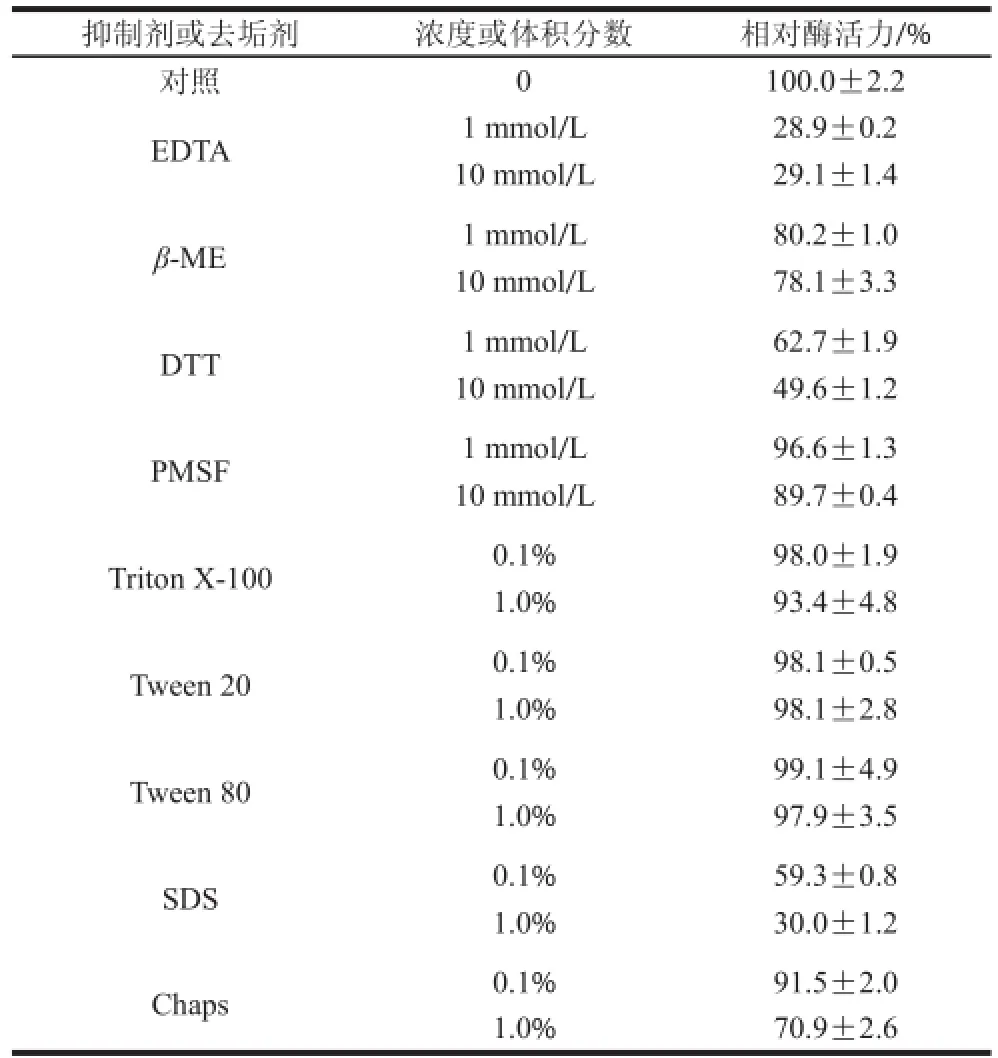
表 2 抑制剂和去垢剂对突变酶的影响Table 2 Effects of inhibitors and detergents on mutant arylsulfatase activity
由表2可知,抑制剂EDTA、β-ME和DTT都对酶活力有抑制作用,其中最明显的是EDTA,使得芳香基硫酸酯酶失去超过70%的酶活力。PMSF对酶活性没有强烈的抑制效果,在浓度为10 mmol/L时,芳香基硫酸酯酶仍可保留89.7%的相对酶活力。去垢剂SDS和1%的Chaps对酶活力有抑制作用,其中SDS抑制效果显著,在加入1%的SDS时,芳香基硫酸酯酶失去近70%的酶活力。突变酶对Triton X-100、Tween 20和Tween 80有良好的抗性。
2.7 动力学参数
以p-NPS为底物,用Lineweaver-Burk双倒数法做图,计算突变酶的动力学参数。M4-153的Km和vmax值分别为0.73 mmol/L和12.21 μmol/(mg·min)。
2.8 脱除龙须菜粗多糖的硫酸基团
由表3可知,在45 ℃条件下处理4 h的条件下,随着加酶量的增多,龙须菜粗多糖被脱除的硫酸基团就越多。当加酶量为90 U时,M4-153能脱除79.5%的硫酸基团,而野生型脱除率为76.3%。
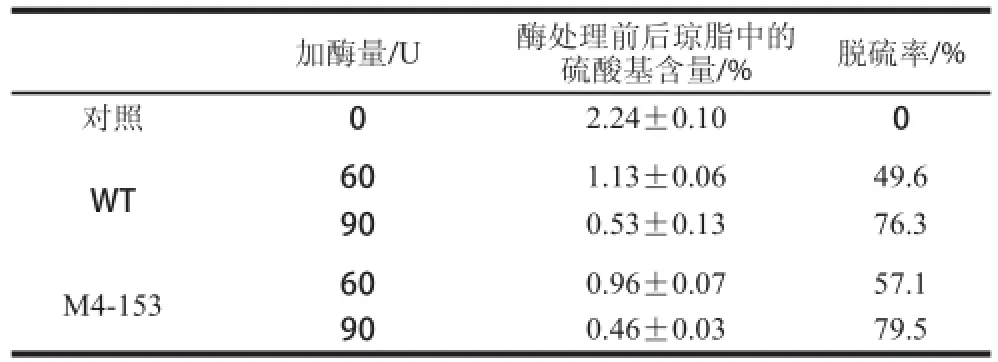
表 3 突变型和野生型芳香基硫酸酯酶对龙须菜粗多糖硫酸基团的脱除Table 3 Effects of enzymatic treatment with purified mutant and wide-type arylsulfatases on the desulfuration efficiency of crude polysaccharides from Gracilaria lemaneiformis
3 讨 论
定向进化是一种提高酶热稳定性、底物特异性等的有效方法。基于易错PCR的定向进化方法,由于简单易行而被广泛应用[26-27]。本研究利用易错PCR方法构建了一个P. carrageenovora芳香基硫酸酯酶突变体库,并筛选得到热稳定性提高的突变酶M4-153。
琼脂是典型的热融性分子,需要加热以成为液体状态,它的融解温度为43 ℃,温度保持在45 ℃以上时不会形成凝胶。因此,水解琼脂硫酸基团的酶在45 ℃以上保持稳定才能在工业应用上有良好的前景。本研究中,突变酶的热稳定性分析结果显示,酶在高于45 ℃时热稳定性提高。在相同的条件下,M4-153对龙须菜琼脂硫酸基团的脱硫率高于WT,并且M4-153在pH 5.0~9.0范围内保持稳定,这些提高了突变酶的工业应用前景。
EDTA对M4-153的酶活性有较强的抑制效果,这表明金属离子在酶的催化过程中起主要作用。SDS对酶活力也有着抑制作用,推测是由于SDS与芳香基硫酸酯酶的疏水性氨基酸残基上的烷基结合并造成影响,来自Marinomonas sp. FW-1的芳香基硫酸酯酶也有着相似的结果[16]。M4-153在一些洗涤剂的存在下仍然保持相对稳定,如Triton X-100、Tween 20、Tween 80和Chaps,这些性质提高了该突变酶在工业利用过程中的潜在价值。
硫酸基含量是琼脂的一个重要质量标准[28],芳香基硫酸酯酶是一种可在温和条件下进行琼脂脱硫的理想候选酶。本研究中当突变酶M4-153的加酶量达90 U,在45 ℃处理4 h后,龙须菜粗多糖硫酸基含量下降了79.5%,该突变芳香基硫酸酯酶是琼脂脱硫应用中的潜在催化剂。
[1] DUCKWORTH M, YAPHE W. The structure of agar: part Ⅰ. Fractionation of a complex mixture of polysaccharides[J]. Carbohydrate Research, 1971, 16(1): 189-197. DOI:10.1016/S0008-6215(00)86113-3.
[2] ARNOTT S, FULMER A, SCOTT W E, et al. The agarose double helix and its function in agarose gel structure[J]. Journal of Molecular Biology, 1974, 90(2): 269-284. DOI:10.1016/0022-2836(74)90372-6.
[3] GUISELEY K B. The relationship between methoxyl content and gelling temperature of agarose[J]. Carbohydrate Research, 1970, 13(2): 247-256. DOI:10.1016/S0008-6215(00)80831-9.
[4] 黄婷婷, 叶李艺, 沙勇, 等. 龙须菜提取琼胶碱处理工艺条件优化[J]. 化学工程与装备, 2010(10): 12-15. DOI:10.3969/ j.issn.1003-0735.2010.10.004.
[5] BOLTES I, CZAPINSKA H, KAHNERT A, et al. 1.3 ☒ structure of arylsulfatase from Pseudomonas aeruginosa establishes the catalytic mechanism of sulfate ester cleavage in the sulfatase family[J]. Structure, 2001, 9(6): 483-491. DOI:10.1016/S0969-2126(01)00609-8.
[6] PAIETTA J V. Molecular cloning and regulatory analysis of the arylsulfatase structural gene of Neurospora crassa[J]. Molecular and Cellular Biology, 1989, 9(9): 3630-3637. DOI:10.1128/ MCB.9.9.3630Mol.Cell.Biol.
[7] BEIL S, KEHRLI H, JAMES P, et al. Purification and characterization of the arylsulfatase synthesized by Pseudomonas aeruginosa PAO during growth in sulfate-free medium and cloning of the arylsulfatase gene (atsA)[J]. European Journal of Biochemistry, 1995, 229(2): 385-394. DOI:10.1111/j.1432-1033.1995.0385k.x.
[8] YAMADA K, AKASAKA K, SHIMADA H. Structure of sea-urchin arylsulfatase gene[J]. European Journal of Biochemistry, 1989, 186(1/2): 405-410. DOI:10.1111/j.1432-1033.1989.tb15223.x.
[9] HOSTOS E L, SCHILLING J, GROSSMAN A R. Structure and expression of the gene encoding the periplasmic arylsulfatase of Chlamydomonas reinhardtii[J]. Molecular and General Genetics, 1989, 218(2): 229-239. DOI:10.1007/BF00331273.
[10] WITTSTOCK U, FISCHER M, SVENDSEN I, et al. Cloning and characterization of two cDNAs encoding sulfatases in the Roman snail, Helix pomatia[J]. International Union of Biochemistry and Molecular Biology Life, 2000, 49(1): 71-76. DOI:10.1080/713803591.
[11] WAHEED A, RISLEY J M, van ETTEN R L. Structural and immunological relationships among mammalian arylsulfatase a enzymes[J]. Comparative Biochem istry and Physiology Part B: Comparative Biochemistry, 1985, 82(4): 855-862. DOI:10.1016/0305-0491(85)90535-8.
[12] STEVENS R L, FLUHARTY A L, KILLGROVE A R, et al. Microheterogeneity of arylsulfatase a from human tissues[J]. Biochimica et Biophysica Acta-Enzymology, 1976, 445(3): 661-671. DOI:10.1016/0005-2744(76)90118-2.
[13] BENKOVIC S J, VERGARA E V, HEVEY R C. Purification and properties of an arylsulfatase from Aspergillus oryzae[J]. Journal of Biological Chemistry, 1971, 246(16): 4926-4933.
[14] OKAMURA H, YAMADA T, MUROOKA Y, et al. Purification and properties of arylsulfatase of Klebsiella aerogenes identity of the enzymes formed by non-repressed and de-repressed synthesis[J]. Agricultural and Biological Chemistry, 1976, 40(10): 2071-2076. DOI: 10.1080/00021369.1976.10862350.
[15] MIECH C, DIERKS T, SELMER T, et al. Arylsulfatase from Klebsiella pneumoniae carries a formylglycine generated from a serine[J]. Journal of Biological Chemistry, 1998, 273(9): 4835-4837. DOI:10.1074/jbc.273.9.4835.
[16] WANG X Y, DUAN D L, XU J C, et al. Characterization of a novel alkaline arylsulfatase from Marinomonas sp. FW-1 and its application in the desulfation of red seaweed agar[J]. Journal of Industrial Microbiology and Biotechnology, 2015, 42(10): 1353-1362. DOI:10.1007/s10295-015-1625-6.
[17] KIM D E, KIM K H, BAE Y J, et al. Purification and characterization of the recombinant arylsulfatase cloned from Pseudoalteromonas carrageenovora[J]. Protein Expression and Purification, 2005, 39(1): 107-115. DOI:10.1016/j.pep.2004.09.007.
[18] MARINO T, RUSSO N, TOSCANO M. Catalytic mechanism of the arylsulfatase promiscuous enzyme from Pseudomonas aeruginosa[J]. Chemistry, 2013, 19(6): 2185-2192. DOI:10.1002/chem.201201943.
[19] HENDERSON M J, MILAZZO F H. Arylsulfatase in Salmonella typhimurium: detection and influence of carbon source and tyramine on its synthesis[J]. Journal of Bacteriology, 1979, 139(1): 80-87.
[20] MUROOKA Y, YIM M H, HARADA T. Formation and purification of Serratia marcescens arylsulfatase[J]. Applied and Environmental Microbiology, 1980, 39(4): 812-817.
[21] KIM J H, BYUN D S, GODBER J S, et al. Purification and characterization of arylsulf atase from Sphingomonas sp. AS6330[J]. Applied Microbiology and Biotechnology, 2004, 63(5): 553-559. DOI:10.1007/s00253-003-1463-8.
[22] UEKI T, SAWADA Y, FUKAGAWA Y, et al. A new type of st reptomycete arylsulfatase with high affinity to the sulfuryl moiety of the substrate[J]. Bioscience, Biotechnology, and Biochemistry, 1995, 59(6): 1069-1075. DOI:10.1271/bbb.59.1069.
[23] LEE D G, SHIN J G, JEON M J, et al. Heterologous expression and characterization of a recombinant thermophilic arylsulfatase from Thermotoga maritima[J]. Biotechnology and Bioprocess Engineering, 2013, 18(5): 897-902. DOI:10.1007/s12257-013-0094-x.
[24] SHUKLA M K, KUMAR M, PRASAD K, et al. Partial characterization of sulfohydrolase from Gracilaria dura and evaluation of its potential application in improvement of the agar quality[J]. Carbohydrate Polymers, 2011, 85(1): 157-163. DOI:10.1016/ j.carbpol.2011.02.009.
[25] BRADFORD M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of proteindye binding[J]. Analytical Biochemistry, 1976, 72(1/2): 248-254. DOI:10.1016/0003-2697(76)90527-3.
[26] MABROUK S B, AYADI D Z, HLIMA H B, et al. Thermostability improvement of maltogenic amylase MAUS149 by error prone PCR[J]. Journal of Biotechnology, 2013, 168(4): 601-606. DOI:10.1016/ j.jbiotec.2013.08.026.
[27] LIN L, FU C G, HUANG W Q. Improving the activity of the endoglucanase, Cel8M from Escherichia coli by error-prone PCR[J]. Enzyme and Microbial Technology, 2016, 86: 52-58. DOI:10.1016/ j.enzmictec.2016.01.011.
[28] GUISELEY K B, KIRKPATRICK F H, PROVONCHEE R B, et al. A further fractionation of agarose[J]. Hydrobiologia, 1993, 260(1): 505-511. DOI:10.1007/BF00049063.
Screening and Characterization of Mutant with Improved Thermostability from a Random Mutant Library of Pseudoalteromonas carrageenovora Arylsulfatase
QIAO Chaochao1, WANG Xinxia1, LI Hebin2, NI Hui1, XIAO Anfeng1, ZHU Yanbing1,*
(1. College of Food and Biological Engineering, Jimei University, Xiamen 361021, China; 2. Department of Pharmacy, Xiamen Medical College, Xiamen 361023, China)
A library of Pseudoalteromonas carrageenovora arylsulfatase mutants was constructed by random mutagenesis using error-prone PCR. After screening, one mutant strain named 4-153 was obtained whose arylsulfatase had improved thermal stability. It was found that there were two amino acid substitutions in the mutant, including D84A and H260L. When p-nitrophenyl sulfate was used as a substrate, the optimal reaction temperature for the mutant enzyme was 55 ℃. Mutant arylsulfatase 4-153 (M4-153) retained 85%, 83%, 48%, and 13% of its initial activity after incubation at 45, 50, 55 and 60 ℃for 30 min, respectively. Meanwhile, wild-type arylsulfatase (WT) retained 79%, 68%, 21%, and 1% of its initial activity after incubation at 45, 50, 55 and 60 ℃ for 30 min, respectively. These results showed that M4-153 had a better thermal stability than WT. M4-153 had an optimum pH of 8.0, and it was stable over the pH range of 5.09.0. Inhibition assay with EDTA indicated that metal ions played an important role in the catalytic process of the mutant enzyme. The recombinant arylsulfatase 4-153 showed a relatively strong tolerance to some detected detergents including Triton X-100, Tween 20, Tween 80, and Chaps. The desulfuration ratio of the crude polysaccharides from Gracilaria lemaneiformis by M4-153 was 79.5%.
arylesterase; error-prone PCR; improved thermostability; mutant property
10.7506/spkx1002-6630-201710004
Q814.9
A
1002-6630(2017)10-0018-06
乔超超, 王新侠, 李鹤宾, 等. Pseudoalteromonas carrageenovora芳香基硫酸酯酶突变文库热稳定性提高突变体的筛选及鉴定[J]. 食品科学, 2017, 38(10): 18-23. DOI:10.7506/spkx1002-6630-201710004. http://www.spkx.net.cn
QIAO Chaochao, WANG Xinxia, LI Hebin, et al. Screening and characterization of mutant with improved thermostability from a random mutant library of Pseudoalteromonas carrageenovora arylsulfatase[J]. Food Science, 2017, 38(10): 18-23. (in Chinese with English abstract)
10.7506/spkx1002-6630-201710004. http://www.spkx.net.cn
2016-09-02
国家自然科学基金青年科学基金项目(31401632);福建省高校新世纪优秀人才支持计划项目(B15139)
乔超超(1991—),男,硕士研究生,主要从事食品科学研究。E-mail:451028523@qq.com
*通信作者:朱艳冰(1976—),女,副教授,博士,主要从事食品科学研究。E-mail:yanbingzhu@163.com
