红松林天然更新对连续小梯度海拔变化的响应1)
2017-06-05张凡兵龙婷金文斌李景文
张凡兵 龙婷 金文斌 李景文
(北京林业大学,北京,100083) (武警驻北京林业大学选培办) (北京林业大学)
红松林天然更新对连续小梯度海拔变化的响应1)
张凡兵 龙婷 金文斌 李景文
(北京林业大学,北京,100083) (武警驻北京林业大学选培办) (北京林业大学)
海拔是影响森林类型分布及其更新的重要生态因子,研究海拔梯度上森林更新的变化,对不同海拔梯度的森林经营与生态保护具有重要意义。以红松母树林群落的天然更新为研究对象,在小兴安岭地区东折棱河林业经营所56号林班,以连续小梯度海拔变化,沿林班内山坡下部、中下部、中部3个坡位,设置12块样地进行调查。结果表明:(1)无人为干扰条件下,随海拔梯度升高,林内针、阔叶树总株数逐增,立木层(红松、臭冷杉、鱼鳞云杉等)针叶树逐渐取代伴生阔叶树(青楷槭、花楷槭、黄桦、白桦等);林冠下层,针、阔叶树更新苗株数总体呈上升趋势,针叶树更新苗数量与阔叶树差距逐渐缩小,在海拔447 m时,针叶树更新数量首次超过阔叶树更新数量。(2)有人为干扰条件下,立木层林木稀疏,林内针、阔叶树更新不良,样地H主要树种更新频度均为0,林窗下以阔叶树更新为主(样地F、I仅有1种更新苗,分别为花楷槭与青楷槭),红松仅有极少幼树存在。(3)随着海拔梯度升高,红松断面积组成与重要值变化趋势基本一致,均保持稳定增长,红松群落优势更加明显。(4)海拔变化对立木层红松分布格局无明显影响,低海拔区红松无更新或更新极少且呈随机分布,中海拔区红松更新良好且多呈聚集分布,在演替过程中,最终以随机分布进入主林层。
小兴安岭;原始红松林;小梯度海拔;天然更新;空间格局
红松属阳性树种,是我国东北地区的主要树种之一[1],以红松为建群种的红松林是我国小兴安岭地区森林演替的顶级群落[2]。近年来,随着人为干扰活动加剧,小兴安岭地区原始红松林面积急剧缩小,关于红松天然更新的研究成了当前红松群落研究的热点[2]。目前,国内就东北地区红松林的群落特征[3]、物种组成[4-5]、物种多样性[6-8]、空间分布格局[9]及其主要伴生树种的生态学特征[10]、天然更新情况[11-12]等方面做了大量研究。对于红松天然更新,主要围绕不同林分或不同抚育(伐除)方式对红松天然更新[13-15]、格局及其年龄结构的影响[16]、林窗、林隙及光照条件与红松天然更新格局及动态的研究[17-25]、动物与红松天然更新机制[9]、红松种子传播与萌发的时空分布格局及红松更新的综合影响因子[2]等方面开展研究。刘晓东[14]等人对小兴安岭红松天然更新的影响因子进行灰色关联分析,表明红松在中低海拔生长、更新良好,为该区域优势种,其天然更新受郁闭度、海拔、坡度、光照、林,聂志文等[15]认为海拔及坡度差异对红松分布具有重要影响。相关研究更侧重于不同生态因子对红松天然更新影响的综合排序,而选取的指标值对红松天然更新的具体影响并未做详尽分析。因此,本文以原始红松林为研究对象,选择影响红松天然更新的重要因子海拔高度,沿山坡下部、中下部、中部3个坡位,设置了连续小梯度海拔变化的12块样地,对样方内红松天然更新情况进行调查,以期进一步探究红松天然更新与海拔变化的动态关系。通过对红松天然更新的主要参考指标(更新密度、更新频度、更新组成、更新格局)与影响红松天然更新的重要因子(林木株数、针叶树株数、冠下植被盖度、红松断面积组成与重要值)随连续小梯度海拔变化的响应进行分析。揭示了中低海拔区红松群落天然更新与连续小梯度海拔变化的关系,为红松林保护与管理提供科学依据。
1 研究地概况
研究地点设在小兴安岭东折棱河自然保护区56号林班内,海拔340~542 m,该林班主要以低山缓坡为主,平均坡度为12°,最大坡度为20°,土壤为典型暗棕壤。东折棱河经营所(128°30′~129°24′E,46°29′~47°6′N)始建于1975年,施业区总面积7 039 hm2,区内最高海拔1 180 m,属北温带大陆性湿润季风气候。夏季最高气温达36 ℃,冬季最低气温达-37 ℃,冬季极端气温达-41 ℃,年平均气温1 ℃,无霜期95~120 d,≥0 ℃的积温1 900~2 200 ℃,年平均降水量610 mm。2005年东折棱河林业经营所被批准为省级自然保护区,经营所从2004年第4季度停止木材采伐,目前以季节性的营林生产作业和多种经营生产为主要工作。全施业区共划分为22个林班,有林地面积6 668 hm2。天然红松种子林1 641 hm2,主要分布于56、58、60等林班,其中56林班为红松母树林。天然云杉种子林37 hm2,其它天然林1 300 hm2。施业区内主要以红松(Pinuskoraiensis)、紫椴(Tiliaamurensis)、糠椴(T.mandshurica, Tm)、水曲柳(Fraxnusmandshurica)、核桃楸(Juglansmandashurica)、臭冷杉(Abiesnephrolepis)、青楷槭(Acertegmentosum)、花楷槭(Acerukurunduense)等为主的针阔混交林组成。主要灌木有:东北溲疏(Deutziaparviflora)、刺五加(Acanthopanaxsenticosus)、毛榛(Corylusmandshurica)、堇叶山梅花(Philadelphushenryi)、曲萼绣线菊(Spiraeaflexuosa)等。主要草本有:人字果(Dichocarpumhypoglaucun)、小叶芹(Aegopodiumalpestre)、粗茎鳞毛蕨(Dryopteriscrassirhizoma)等。中低海拔主要形成以红松为优势的针阔混交林。红松纯林仅分布在坡度较陡的高海拔地段,大部分位于相邻林班界限处。
2 研究方法
2.1 样地设置与调查
采用线路踏查和样地实测法相结合,在东折棱河林业经营所56号林班中低海拔区,选择红松母树林长势较好且分布均匀的林分,进行连续小梯度(3 m<Δh<15 m)海拔变化的样地设置。在林班内,沿山坡下部(样地A~F)、山坡中下部(样地G~I)、山坡中部(J~L)3个坡位,设置12块样地,各样地面积均为30 m×30 m,经坡度矫正后,各样方闭合差均小于1/200,采用相邻格子法进行群落特征与更新调查。将每个样地分为36个5 m×5 m小样方,调查并记录各小样方内,乔木的种名与数量,应用上限排外法对胸径大于4 cm的林木进行每木检尺,测量树高、胸径、冠幅,死枝下高,活枝下高。在每个5 m×5 m的小样方内,随机选取1个2 m×2 m的小样方为灌木样方,于样方内4个角落各设置1 m×1 m的小样方为草本样方,按物种记录株数、平均高、盖度、生长状况(优、良、中、差)、分布状况(随机,聚集,均匀)。采用百步抬头法记录林内郁闭度。此外,样地记录内容还包括样地位置(海拔、坡向、坡位、坡度),林内干扰方式及强度(样地E~I均存在不同程度的干扰活动。样地E、F的干扰源于林区择伐、松果采摘、冬季运输枯死木、伐倒木等人为活动,最终形成部分林窗。样地G~I的干扰分别源于枯死木、残存雪压木形成的小面积林隙,其干扰程度较样地E、F轻)。最后,进行土壤与群落最小面积调查。更新调查中,根据高度把更新对象分为幼苗、幼树,针叶树其高度小于10 cm为幼苗,相当于1~3年生,更新层以下,树高大于10 cm的为幼树;阔叶树其高度小于30 cm为幼苗,更新层以下,高于30 cm为幼树。在外业调查过程中,幼苗计入草本层,幼树计入灌木层(见表1)。所列主要树种取自各样方调查树种,经计算后重要值前五名,考虑到部分样方红松重要值较低,则按重要值大小取至红松处。
2.2 数据处理
分别采用Excle 2007、SPSS 19.0H以及SigmaPlot 12.5等软件进行数据整理、回归分析与统计检验以及图形绘制。对各海拔区样方内数据进行整理与分析,围绕描述红松天然更新的重要指标(更新频度、更新密度、更新组成与红松幼苗分布格局)与红松天然更新的重要影响因子(林内针、阔叶乔木株数,针、阔叶更新苗株数,群落内乔木、灌木、草本层各物种重要值,红松母树重要值、红松断面积组成),对红松天然更新随连续小梯度海拔变化进行分析。部分指标计算公式如下:
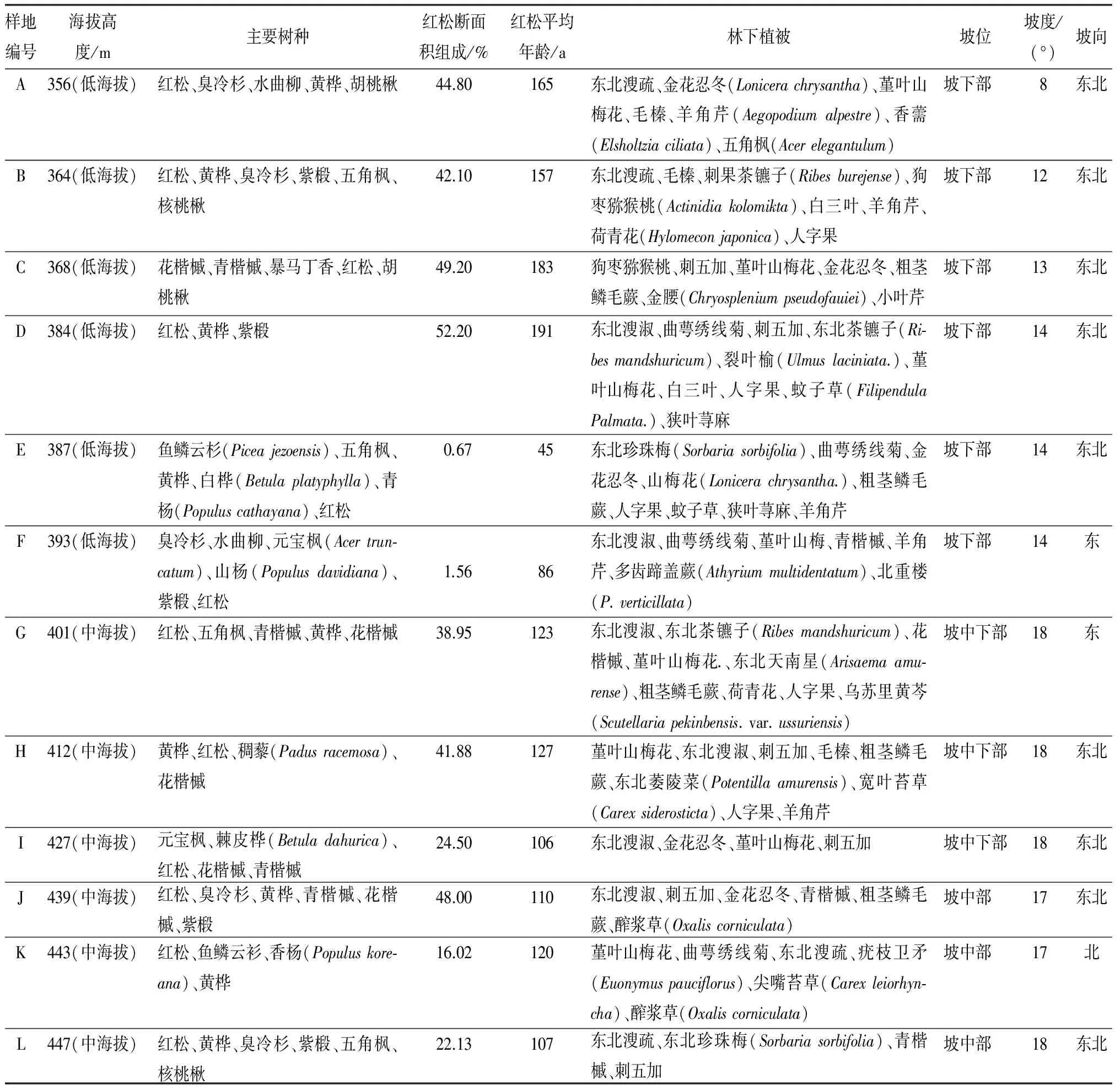
表1 调查样地的基本情况
乔木树种的重要值=(相对密度+相对频度+相对显著度)/3;
灌木或草本的重要值=(相对密度+相对频度+相对盖度)/3;
更新密度=株数合计×10000/(样方面积×样方数);
更新频度=(某种幼树出现样方数/调查样方数)×100%。
植物种的空间分布格局计算公式:
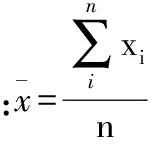
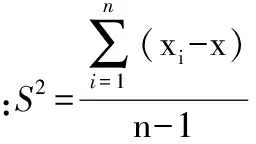

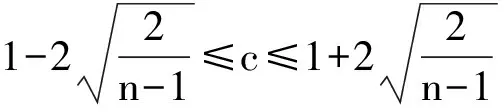
3 结果与分析
3.1 低海拔地段原始红松林的天然更新
由表1、表2可知,样地A~D主要以红松为主。4个样方有红松33株,树高最高达26.8m,平均年龄大于150a。断面积组成分别为44.8%、42.1%、49.2%、52.2%,重要值分别为18.9、9.5、17.41、36.4。其他混交阔叶树主要有:花楷槭、青楷槭、核桃楸、水曲柳、紫椴等伴生种,偶有香杨等偶见种,阔叶树合计173株。混交针叶树主要是臭冷杉,偶有五角枫,合计60株。可见,在山坡下部低海拔区,红松母树虽然数量不多,但长势良好,在林分中占据着重要地位,控制林分主林层。在演替层,阔叶树凭借大量株数占据着林下,而混交针叶树则很少。林内郁闭度在0.7左右。阔叶树更新苗81株,最大更新密度花楷槭(1 296株/hm2),更新频度19%,其他针叶树更新苗15株,最大更新密度为臭冷杉(926株/hm2),更新频度22%,可见林内阔叶树更新占优。林下青楷槭、花楷槭等阔叶树在更新组成中占4~6成,而红松占1成或没有更新。本文研究进一步表明林木株数特别是同一类型的母树株数对相应类型的更新苗数量影响较大,一定数量的针叶树使得林内光照条件得到改善,在生长条件适宜情况下,以优势数量存在的伴生阔叶树,其更新苗数量将远超林内优势树种的更新苗数量。林内红松更新密度最大为370株/hm2。经分析,距离母树较近的杨桦林使得松鼠等动物传播种源几率增大,同时杨桦林内初期庇荫条件好,中后期充足光照的特点适宜红松生长习性,林下更新苗年龄跨度大(5~65a),而母树林内更新苗年龄跨度小(2~30a),这种由种源数量相差不大,但更新苗年龄跨度差异显著的特点,造就了红松母树林与杨桦红松林更新密度的差距。后者主要与林内更新植株盖度有关,该样方调查的红松母树林与水曲柳红松林群落特征相近,林下植被茂盛,郁闭度较大,茂密的植被阻挡了红松种子与土壤的接触几率,即使接触也因光照不足而影响种子生长、发育。
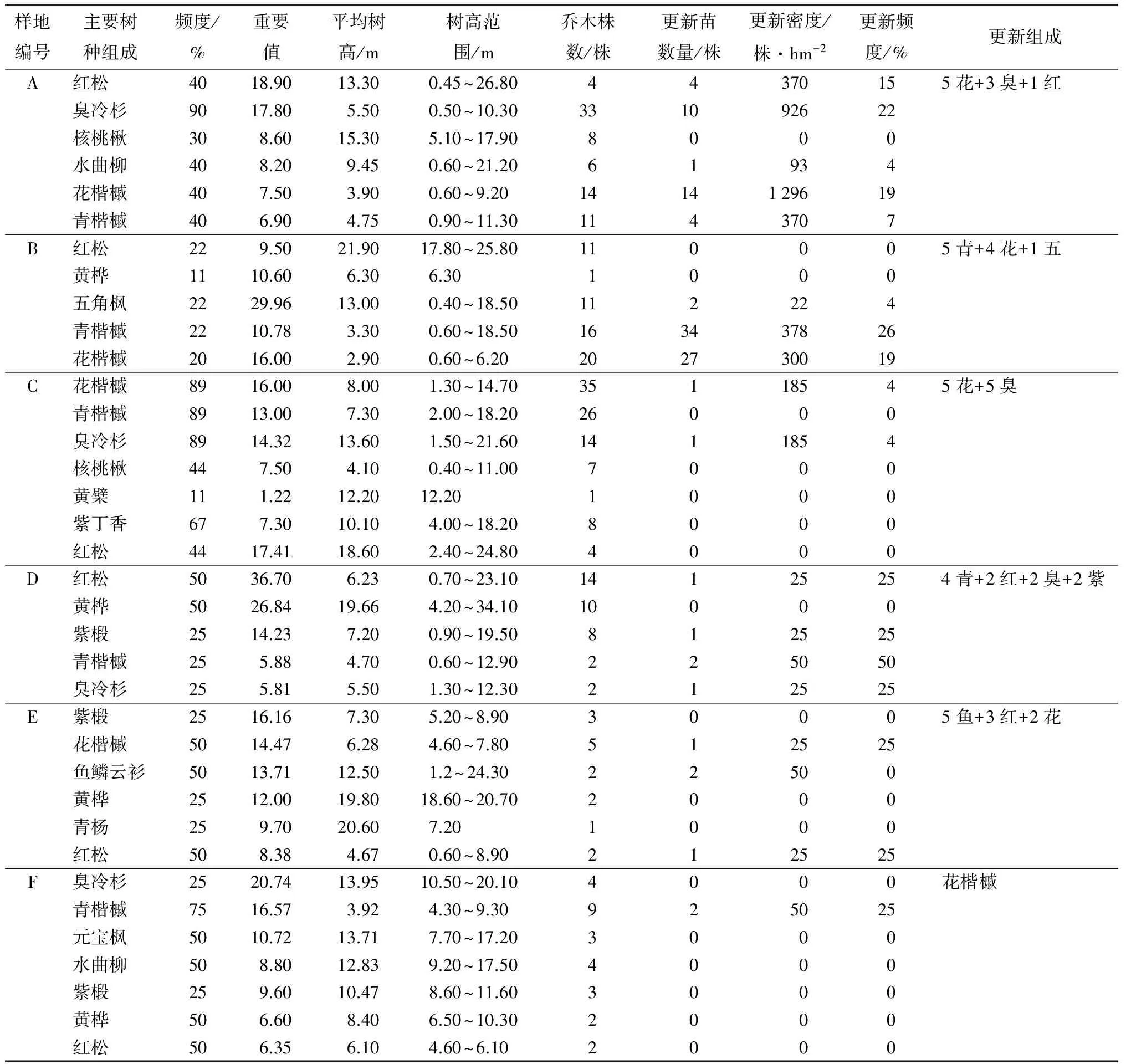
表2 低海拔地段原始红松林天然更新情况
样地E、F现存立木主要以青楷槭、黄桦、花楷槭等阔叶树为主,并伴有元宝枫、紫椴等,阔叶树合计32株。针叶树共10株,红松4株。2块样地中红松断面积组成分别为0.67%、1.56%,重要值分别为8.38、6.35。可见,在低海拔地区,经过择伐(高强度人为干扰)下的红松母树林,红松已不占优势,优势树种主要是枫桦、青楷槭、紫椴等阔叶树,这类阔叶树数量相对样方A~D中阔叶树仍较少。林内上方郁闭度小,林窗明显。林下灌木生长极为茂盛,更新样方植株盖度分别为20.9%,7.71%,均高于前4个样方。林下灌木以毛榛、堇叶山梅花、金花忍冬、东北珍珠梅为主,盖度8%~80%,平均盖度26.5%;林下草本平均盖度15.17%。林内针、阔叶树株数较少,加之人为干扰严重,使得林内更新不良。林木株数是影响天然更新的重要因子,同一林型林木株数(针叶树株数)愈少,群落内更新愈差。此外,人为干扰活动是影响本样方内红松天然更新的重要因子。尽管样地更新不良,但在更新组成中,样方E为针叶树占优,而样方F则为阔叶树占优且更新苗仅为花楷槭。这与林内母树有关外(样地E主要针叶树占优8株,而阔叶树仅3株,样地F主要阔叶树占优17株,针叶树仅2株),还与各样方林下植被盖度有关(样地E盖度远大于样地F)。研究表明,林下植被形成的一定盖度对耐荫树种的幼苗生长具有促进作用,这与郁闭的林下环境严重阻碍草本或其他树种生长,有利耐荫树种形成竞争优势。当这些幼苗的生长超过草本层,冠下盖度依然过大,会影响其对光照的需求,此时,林窗的出现有助于耐荫树种由幼苗向幼树进行转变[15]。调查发现,样方E中更新的主要阔叶树仅1株,其余4株为均为耐荫针叶树,树高均大于1 m的幼树,样方F中仅有2株花楷槭幼树。在合理经营这一海拔段时,应严格控制人为活动,减小人为干扰,尤其是减少对红松等母树的破坏,以保证林内足够种源及幼苗。同时,合理保留部分林隙或林窗,以促进群落内幼树生长。
3.2 中海拔地段原始红松林天然更新
由表3可知,样地G、H、I现存立木以红松为主,3块样地红松断面积组成分别为38.95%、41.88%、24.50%;重要值分别为7.48、23.44、17.40。红松株数虽然较少,却是这一区域的优势种,控制着主林层,青楷槭、花楷槭、黄桦、棘皮杹、元宝枫等阔叶树作为伴生树种得以大量存在。在调查过程中,这一区域有部分干扰,源于部分雪压木,造成了林中一定面积的林隙。林下植被光照充足,长势较好,三块样地林下植株盖度均大于9%,盖度较大。灌木以东北茶镳子、毛榛、刺果茶镳子、东北溲淑、堇叶山梅花为主,草本以东北萎陵菜、羊角芹、粗茎鳞毛蕨等为主。林下阔叶树更新比针叶树更新良好,其原因与样地E、F一致。红松更新较差,这一点从更新密度、更新频度与更新组成中即可看出。究其原因可能与地面种源减少(2012年当地红松结果为小年,居民秋末入山采摘松果,鼠类动物缺食影响种源运输与储藏,进而影响红松更新)有关。此外,林隙面积过大,灌草茂盛,进而影响红松幼苗生长的光照条件。
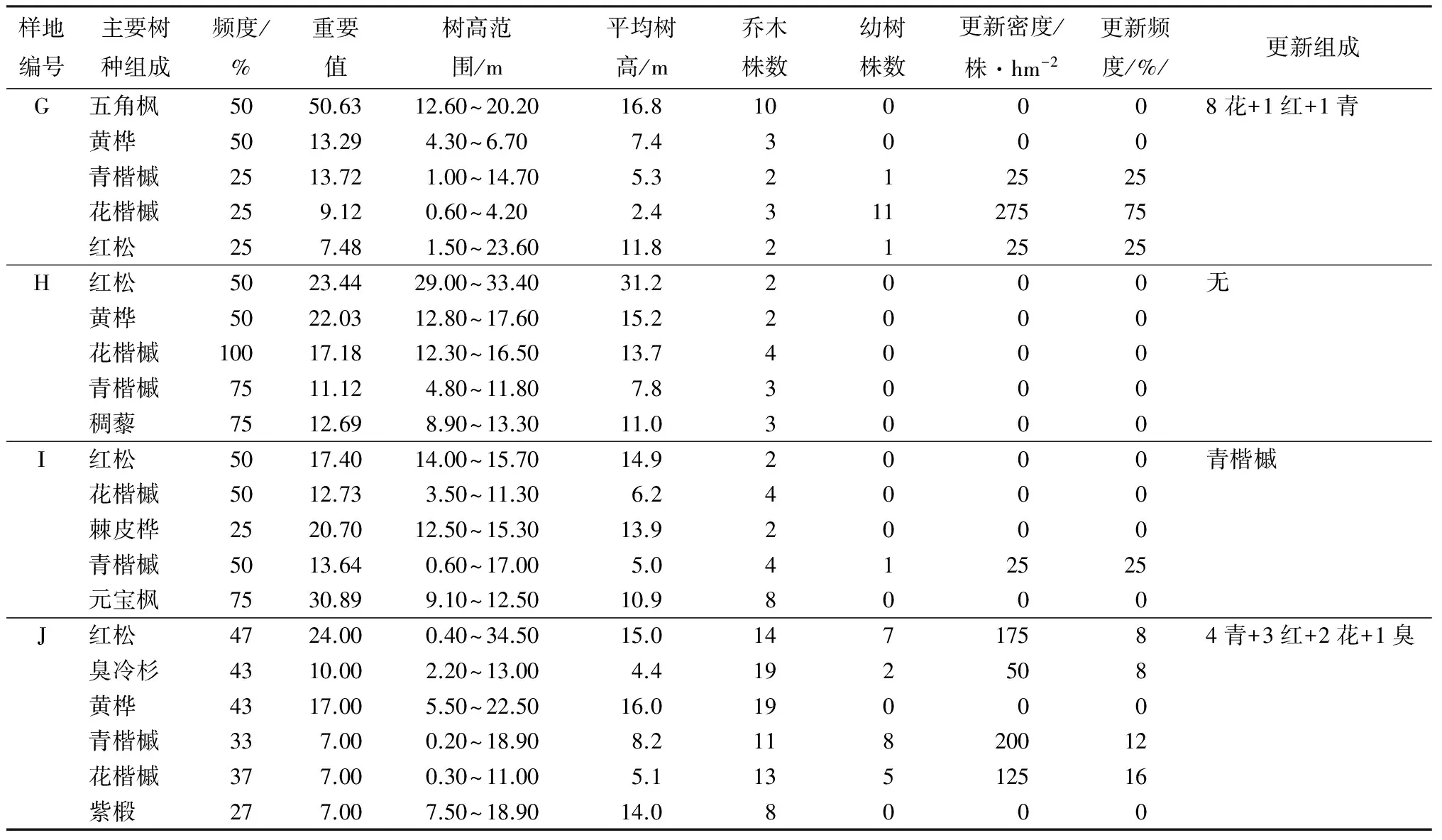
表3 中海拔地段原始红松林天然更新情况
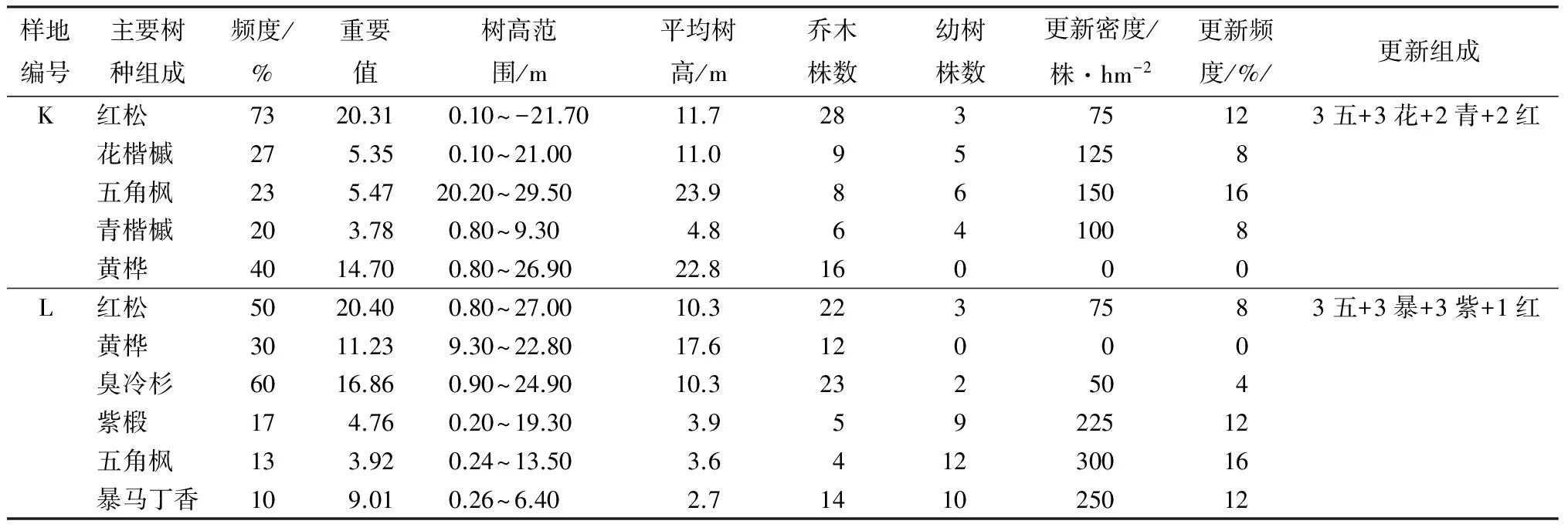
续(表3)
样地J、K、L,立木主要以红松为主,3块样地的红松合计64株,断面积组成最大为48%,最小也达16.02%。红松重要值均在20%以上。样地内伴生针叶树多为五角枫、臭冷杉,伴生阔叶树多为青楷槭、花楷槭、暴马丁香、紫椴,两者株数相差不大。林内郁闭度为0.8,林下植被盖度小,灌木以狗枣猕猴桃、东北溲淑、珍珠梅为主,草本以醡浆草、白三叶、尖嘴苔草、异叶金腰居多,形成了荫庇的条件。阔叶树更新情况与针叶树更新情况相差不大,由于本样方红松母树较多,红松更新较快。此外,该样地红松天然更新良好还和样地所处的海拔、坡位、坡度有关。研究表明,随着海拔升高,针叶树因独特的生物学特性而适应于高海拔的耐寒条件,在红松适生中低海拔区,海拔升高对红松等针叶树更新更加有利[22];山坡中部的侧方缓坡有利于土壤排水,相对于山坡下部的潮湿土壤,山坡中部的湿润土壤更适合红松更新、生长[26-29]。综合样地A~I,在红松适生的中低海拔区,排除人为干扰,原始红松林天然更新情况并不乐观,大部分在50株/hm2,很多情况下基本无更新,这与许多人研究结论一致[30-31]。究其原因,原始红松林不缺少种源[32-33],但其林冠郁闭度大,林内光线弱,早春幼苗易受鼠害影响。此外,阴湿的林内环境致使球蚜,立枯病等病虫害发生,严重威胁红松幼苗生长[34]。这一海拔区,合理经营原始红松林,要充分考虑到营造既适合于动物传播种子又适合红松幼苗生长的生境上,以10~15 a为界限,适当考虑对林内择伐[35-36]。合理调控林内光线,郁闭度控制在0.6左右[29,37-38],红松的天然更新才可能成功。
3.3 原始红松林群落天然更新的主要影响因子对海拔变化的响应
3.3.1 林内主要针、阔叶树株数对海拔变化的响应
不同的海拔梯度适生不同的林型。由图1可知,在小兴安岭原始红松林适生的中低海拔区,排除部分样地人为干扰后,林木株数随小梯度海拔升高呈小幅度递增;就株数而言,阔叶树是这一区域的主要树种,这种现象在山坡下部更为明显。随着海拔升高,相较于山坡下部,山坡中部立木层针叶树株数与阔叶树株数差距逐渐缩小。在海拔447 m时,该样地内针叶树株数超过了阔叶树株数。不难预测,随着海拔逐渐升高,在原始红松林内针叶树将逐渐取代阔叶树,成为林内的主要树种,这显然是针叶树的生物学特性与生态学特性更适应于高海拔区所致。这一结论与吴晓蒲等[17]的研究结论一致,均表明在红松适宜生长的海拔区,随海拔升高,以红松为优势的针叶树,群落优势将更为明显。样地E~I的林分,由于受人为干扰严重,林木株数锐减,立木层稀疏。样地G因大量五角枫存在,使针叶树多于阔叶树外,其他样方均以大量伴生阔叶树存在。可见,上林层稀疏致使林冠光照条件改变,此时林内主要以喜光阔叶树为主,在演替后期,这些阔叶树逐渐被林冠下耐荫树种代替(如红松等)演变为阔叶红松林。
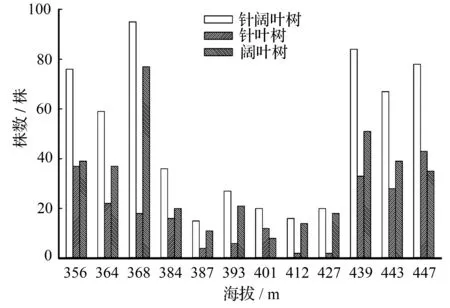
图1 针、阔叶树株数与连续小梯度海拔变化的关系
3.3.2 林内主要针、阔叶树更新苗对海拔变化的响应
海拔能影响植被类型的分布,也会影响海拔梯度上不同植被类型的天然更新。此外,针、阔叶总株数与针叶树株数,分别是影响原始红松林天然更新的两个重要因子。林木株数愈多,林下更新愈好,林内针叶树愈多,林分天然更新愈好[14]。由图2可知,在排除部分样地人为干扰后(样地E~I),随着海拔梯度升高,针、阔叶树更新苗总数呈上升趋势,且针叶树更新苗也逐渐上升(在图2中阔叶树与针叶树更新苗所示折线间距逐步缩短),海拔447 m时,针叶树更新株数超过阔叶树更新株数。更新苗随小梯度海拔变化趋势与其母树随海拔变化趋势一致。样方A、B更新苗总数远高于样地J、K、L并未与海拔梯度增高表现出严格一致的线性关系,究其原因,样地J~L在立木层针阔叶树数量,显著多于样地A、B,上方林层过于荫庇致使冠下光照削弱,更新树木难以生长,而样地A、B更新植株盖度稍大,可见,林内光照条件是较好的。除外,有可能与这一地域活动的动物有关。仔细参照徐振邦等[12]对长白山阔叶红松林天然更新条件的研究,发现针阔叶树更新苗随大尺度海拔变化差异与本文所示趋势线基本一致。在人为干扰明显地区,林窗大面积形成,林下灌草盖度较小,林内多以喜光阔叶树更新为主,且林内更新株数总体较林冠下更新减少。此时林窗下,更新株数多为幼树,这一结论与徐振邦等[12]的研究结果一致,可见,林窗形成的光照条件,不利于红松幼苗而利于其幼树的成长,这与红松在不同年龄段对光照的适应有关。
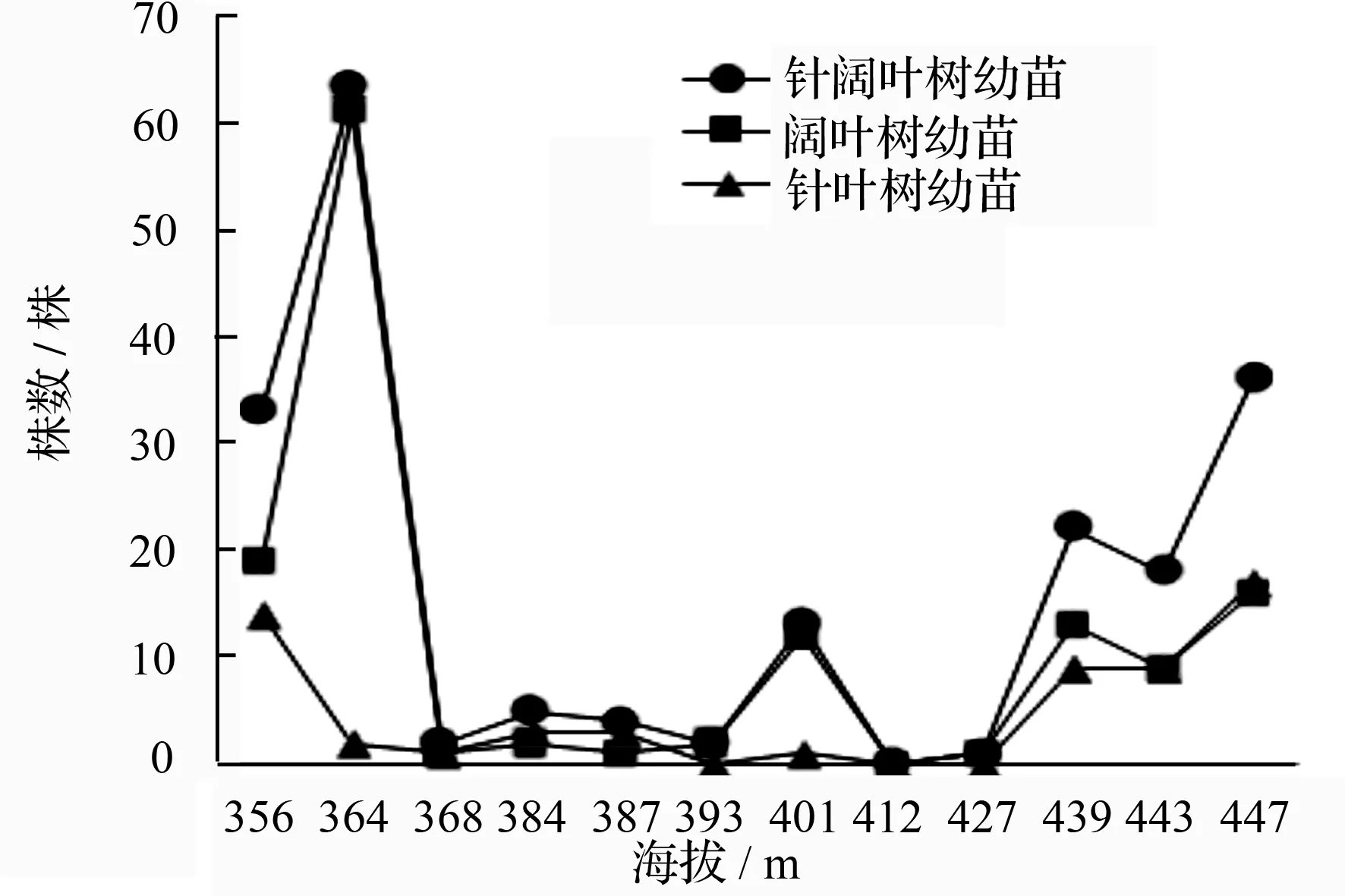
图2 针、阔叶树更新苗数量与连续小梯度海拔变化的关系
3.3.3 红松乔木株数、更新苗株数、断面积组成与重要值对海拔变化的响应
由图3可知,在56号林班内,排除人为干扰因素后(样地A~D,J~L),由山坡下部、山坡中下部至山坡中部,沿着连续小梯度海拔变化,红松更新苗株数与红松母树株数变化趋势基本一致,即红松母树较多区域,红松更新情况相对良好。在人为干扰严重地区(样地E~I),红松母树减少(均为2株),林窗或林隙面积较大,较强光照条件影响耐荫树种红松幼苗的生长,此时,红松更新仅以极少幼树存在。这一结论与有关学者的研究结果一致[18-19]。
红松是小兴安岭东折棱河林场区域森林群落的建群种。红松断面积组成与重要值变化保持一致,随着连续小梯度海拔变化升高,在无人为干扰条件下,两者均稳步上升;虽然在调查地红松株数并未占据最优,最多仅28株(样地K)但并不影响其重要值为20.31%。这与在红松适宜海拔区,随海拔升高,红松凭借较强的生物学与生态学特性适应环境。在人为干扰严重地区,红松虽株数不多,仅2株立木,但过半样地显示其重要值均大于20%,充分显示了红松对该地区森林群落的控制作用。随着海拔升高,红松断面积组成起伏不定(主要是受红松年龄、材积均较大的影响),但红松重要值却稳步增长。结合针叶树株数、红松乔木株数对海拔变化的响应可知,红松将在该林班内,高海拔区占据更重要地位。
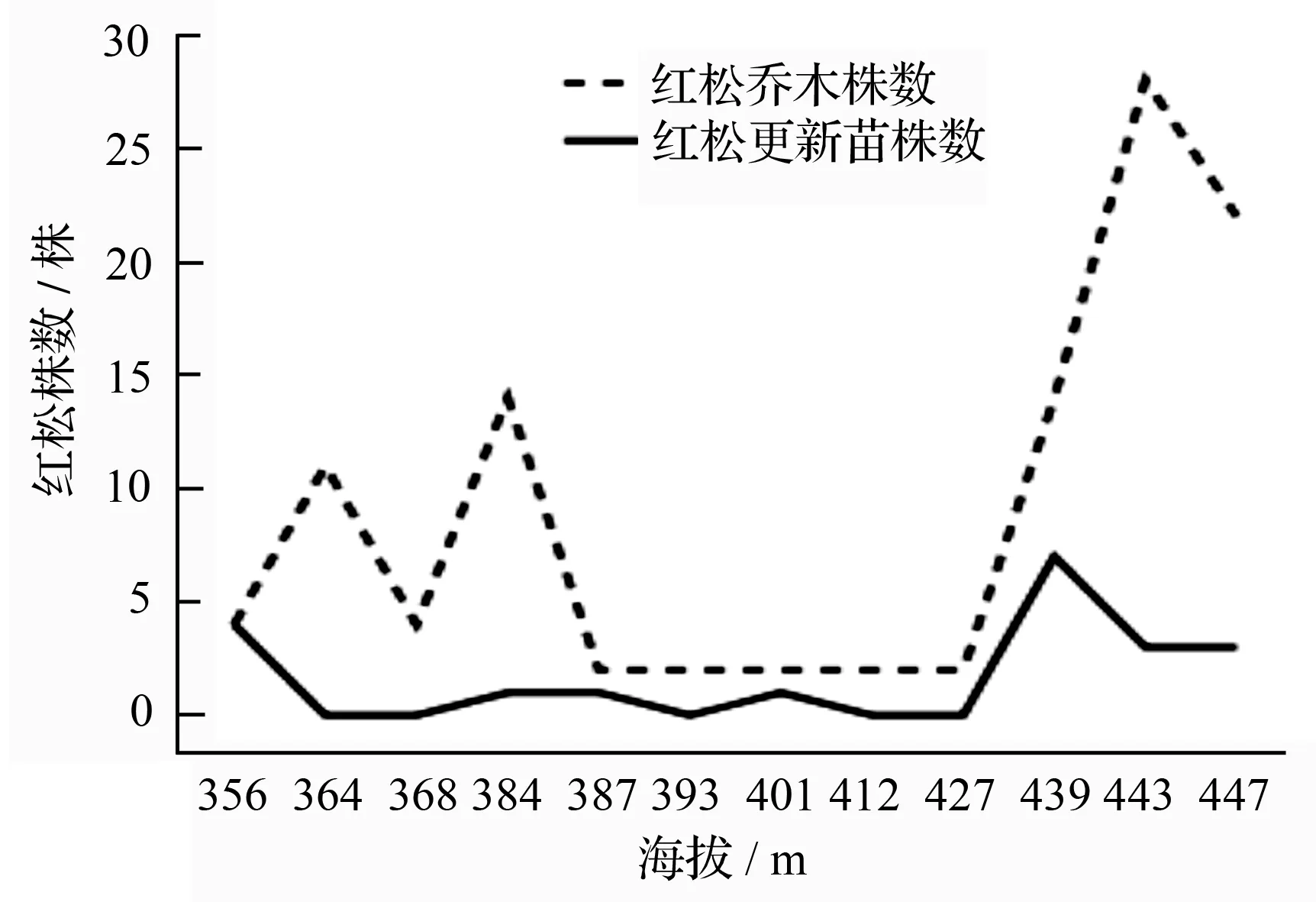
图3 红松乔木株数与更新苗株数
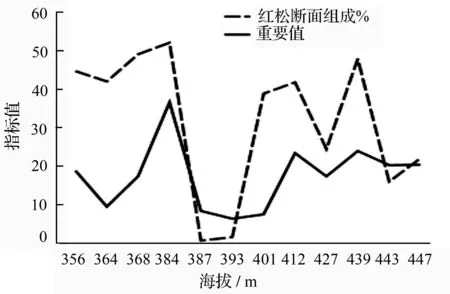
图4 红松断面积组成、重要值与连续小梯度海拔变化的关系
3.4 林内红松乔木与更新幼苗的分布格局对海拔变化的响应
选择山坡下部与山坡中部(海拔跨度约100 m,避免了人为干扰的样地)进行红松分布格局比较,能更好地说明该林班内红松分布格局随海拔变化的差异。对于立木层,山坡下部红松主要为随机分布,山坡中部也以随机分布为主。对冠下更新红松而言,山坡下部仅一个样方有红松更新且呈现随机分布,在山坡中部,3个样方皆有红松更新,且两个表现为聚集分布,另一个为随机分布。这也不难看出,在立木分布格局一致情况下,海拔较高地区红松更新良好,数量较多,其呈现聚集分布与红松母树较多有关外,很可能与动物或鸟类运输、储藏红松种子活动有关[22,39]。高海拔条件下,红松幼苗的这种分布将面临更多的空间与资源的竞争,经过剧烈演替,最终存活的红松幼苗(幼树)凭借其较强的耐荫性与生态学特性进入主林层,呈现出随机分布。这也与现实林分观察到的现象一致。
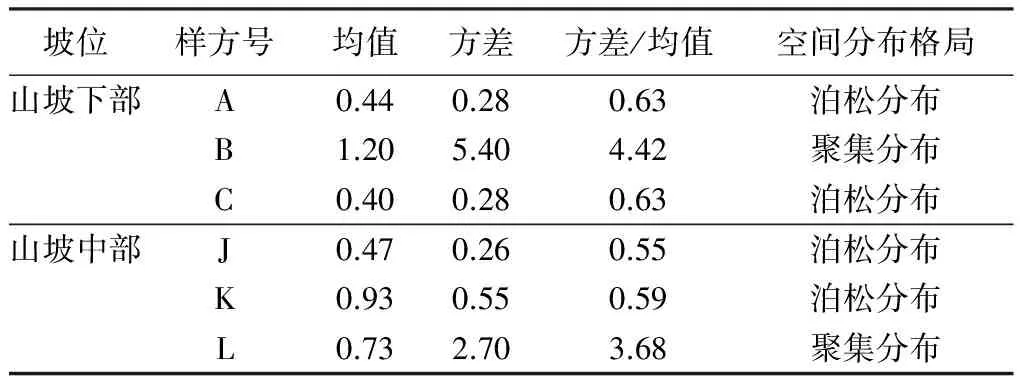
表4 山坡下部与中部红松乔木分布格局
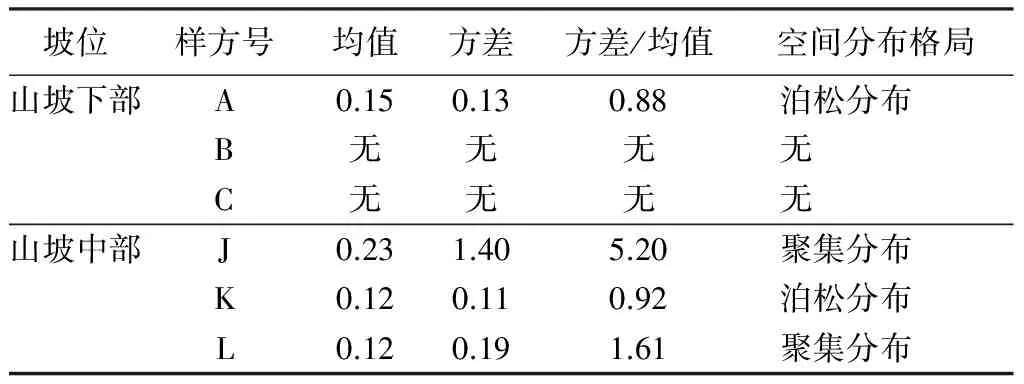
表5 山坡下部与中部红松更新苗分布格局
4 讨论
4.1 海拔与红松林天然更新的关系
海拔能够引起植被类型的变化,小兴安岭地区红松林多集中分布在海拔200~800 m范围内,研究表明海拔与坡度是影响该区红松林分布的主要因子[10]。刘晓东[14]等人通过对小兴安岭地区东折棱河林业经营所(6块样地,海拔跨度398~481 m)不同林型的红松林进行灰色关联分析表明:海拔是影响红松群落天然更新的重要因子,低海拔主要以红松、臭冷杉、红皮云杉和鱼鳞云杉等针叶树更新为主。随着海拔的升高,逐渐出现了蒙古栎、紫椴等树种的更新幼苗,且低海拔条件下,林木更新总株数较高海拔条件下多。而徐振邦等[38]以大尺度海拔变化(7块样地,海拔跨度640~1 270 m),考察了长白山地区红松更新与海拔变化的关系,表明排除采伐时,高海拔条件下,针、阔叶树更新均优于低海拔地区,且高海拔以红松、臭冷杉等针叶树更新为主,这主要与针阔叶树的生物学与生态学特性相关。以上两项研究,在考察海拔变化对更新影响或群落特征时,均在次生红松林中采取株数变化或更新频度、密度等做为林内天然更新的指标。但对红松群落在不同海拔梯度的天然更新及林木优势情况的观点并不一致。本文以小海拔梯度调查原始红松林天然更新情况,发现林木(针叶树、母树)株数是影响原始红松林天然更新的重要因子,这与刘晓东等[14]的研究结论一致。这种差异,可能与当地气候、幼苗、幼树划分有关[9]。红松天然更新需要种源,这就决定了其更新情况与红松母树有关。陶大力等[31]通过人为设置有无动物参与啃食松果的对比实验,更深入论证了红松天然更新对动物啃食后松果的依赖,得出有关动物是红松更新的关键,即红松天然更新与母树数量、动物传播密不可分,母树相对较多地区其获得地面种源较优,更新则获得优先权。
但对于立木株数与更新苗株数对海拔变化的响应,本文更倾向于徐振邦等[12]所述观点。即:在红松适生的中低海拔区,排除人为干扰,随着海拔梯度升高,林内针、阔叶总株数增多,针叶树将逐渐取代阔叶树成为高海拔条件下的主要树种,林内更新渐优,且针叶树更新数量将逐步上升。李景文等[38]也发现在红松适宜生长海拔区,随着海拔梯度升高易行成山坡侧方红松纯林,这除与坡度有关外,也说明高海拔条件下红松较阔叶树生长占优势。
4.2 人为干扰与红松林天然更新的关系
干扰通过影响植物种群结构与群落微生境进而影响林下更新[40-42],小兴安岭地区红松林存在不同程度的人为干扰[42-44]。本文研究表明:排除人为干扰,随着海拔梯度升高,在以红松为建群种的原始红松林内,红松的优势度将更加明显。这也与上一结论(高海拔条件下红松或针叶树优势更加明显)相符。而该结论与吴晓蒲等[17]综合长白山(3块样地,海拔跨度670~890 m)、小兴安岭(4块样地,海拔跨度400~446 m)与大秃子山(张广才岭1块样地1 080 m)等东北地区阔叶红松林群落结构进行分析,表明乔木层红松在低海拔区(小兴安岭优于长白山优于张广才岭)更加占据优势的结论不一致。吴晓蒲等[17]对东北三地红松林考察结果,所得低海拔区红松群落优势更为明显,但三地立地条件各不相同;此外,物种的优势度以重要值进行衡量[2],而胸高断面积仅可作为木本植物计算盖度的指标,还需综合物种出现频度与物种出现密度[3]进行其优势度考虑。胸高断面积并不能完全代替重要值,其两者趋势线有一定差距,仅凭胸高断面积只可大概衡量群落内某物种优势度。
在有人为干扰条件下,乔木层林木减少,林内更新不良,林内的更新苗以幼树居多,这与林窗及林隙的形成改变林内光照环境有关。林内郁闭度过小,植物需光不足,容易受病虫害影响[32,39];林内郁闭度过大,下层灌草旺盛,对幼苗形成不利竞争[2],只有适宜的林内郁闭度才能促进幼苗生长[28]。通过与次生(杨桦,枫桦,椴树)红松林天然更新的研究结论相比,本文调查的原始红松林群落天然更新不良,这一点与张海军等[45]的研究结论一致。
4.3 红松林天然更新的空间格局
幼苗幼树的空间分布格局决定着未来林分的结构和生长动态,不同研究尺度植物的空间分布格局差异较大[9,15]。本文就立木层红松格局与更新层红松格局对海拔变化的响应分析表明:在红松适宜海拔区,小梯度连续海拔变化对红松立木分布格局影响不大,立木多呈随机分布,少有聚集分布,这可能与红松做为该区顶级群落及本文研究尺度有关;低海拔条件下红松较少获得更新或呈随机分布,高海拔条件下,红松更新呈现聚集分布,经过激烈竞争后,在演替过程中以随机分布进入主林层,形成对森林群落的控制。原始红松林冠下更新苗这一分布格局,与王树力等[33]对小兴安岭杨桦林等次生林地红松分布格局呈显著差异,杨桦林下红松更新呈聚集分布是与林分光照环境有关,杨桦林中以喜光阔叶树为主,林内红松是以年龄跨度较大的(5~65 a)且红松幼树居多组成的聚集分布;但与刘庆红[32]对小兴安岭红松种群天然更新研究结果一致,原始红松林有充足种源,其林下更新幼苗多成聚集分布,但遮荫作用强烈,年龄结构简单,多为(2~30 a)。
在借鉴前人研究成果的基础上,本文用更新株数、更新频度、更新密度、更新组成等指标描述群落天然更新情况,对红松天然更新的重要影响因子随海拔变化加以分析,并引进红松断面积组成与其重要值,及红松立木与对应样方下红松更新苗空间格局随海拔变化的差异。结合林下植被盖度、林班干扰情况、动物对松果的储藏、运输及日益严重的人为松果采摘等,解释了不同海拔梯度的红松群落及其更新现状,对指导林班尺度内精细的森林经营与植被恢复提供了科学依据。马建章等[6]研究了松鼠越冬生存策略对红松天然更新的影响,认为在种群尺度上,母树林林隙和距离母树林100~300 m范围内的其它针叶林中,是松鼠贮食密度最高的范围,在逃过其他动物取食后,便成为红松天然更新的种子库,这一结论进一步揭示了红松更新与母树在数量与距离的关系。但,本文并未对红松更新苗与母树的距离、林下种源分布及发育情况,加以观察、记录、分析,在考察及解释林内红松更新时,本试验还有待进一步完善。
在上述讨论的基础上,建议对小兴安岭地区原始红松林在林班尺度内,采取不同海拔梯度段的精细经营。对无人为干扰地段的山坡下部与中部,可考虑采用适当的抚育措施,营造适宜的光照条件促进红松幼苗生长。在人为干扰严重的山坡下部与山坡中度,严禁松果采摘、伐木、运输等活动,适当培植红松或其他珍贵母树,有利于更新苗数量。此外,本文着重小尺度海拔对原始红松林天然更新进行了分析,具体而言,小尺度内不同的地形因子及其他影响因子各自对林分天然更新的定量分离即贡献率将会成为本实验下一阶段研究重点。
5 结论
红松天然更新情况受海拔变化的影响,其实质是由林内针、阔叶树总株数、针叶树株数,坡度,光照条件,动物活动情况等影响红松天然更新的重要因子在不同海拔梯度之间的差异所致。
在山坡下部,坡度较小,土壤潮湿,无人为干扰条件下,红松为群落优势种,乔木层以伴生阔叶树数量占优,此时林下以阔叶树更新为主;存在人为干扰时,林窗出现,林内乔木层针、阔叶树株数减少,此时群落更新不良,阔叶树较针叶树占优,更新苗以幼树为主。在山坡中部,无人为干扰条件下,红松为群落优势种,群落内针、阔叶树株数均增多,立木层针叶树株数首次超过阔叶树株数,更新层针阔叶树更新均良好,针叶树更新株数首次超过阔叶树更新株数。有人为干扰条件下,林隙过大致使林下灌、草长势旺盛,林内幼苗所需光照条件不能满足,加之种源缺少,该样地更新不良。
此外,在以红松为建群种的中低海拔区,以小梯度海拔变化考察原始红松林天然更新情况。表明,随着海拔升高,影响红松天然更新的林木株数、针叶树株数增多,红松在群落内优势度逐步增强;林下针、阔叶更新苗株数增多,与母树株数随海拔变化趋势一致。
[1] 赵秀海,张春雨,郑景明.阔叶红松林林隙结构与树种多样性关系研究[J].应用生态学报.2005,16(12):2236-2240.
[2] 张健,郝占庆,宋波,等.长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性[J].应用生态学报,2007,18(8):1681-1687.
[3] 王旭高,郝占庆,叶吉,等长白山阔叶红松林物种多度和空间分布格局的关系[J].生态学杂志,2008,27(2):145-150.
[4] 陈高,代力民,周莉.受干扰长白山阔叶红松林林分组成及冠层结构特征[J].生态学杂志,2004,23(5):116-120.
[5] 孙越,夏富才,赵秀海,等.张广才岭温带次生针阔混交林物种组成和群落结构特征[J].生态学报,2017,(4):485-488.
[6] 马建章,宗诚,吴庆明,等.凉水自然保护区松鼠贮食生境选择[J].生态学报,2006,26(11):3542-3548.
[7] KERR G. The use of silvicultural systems to enhance the biological diversity of plantation forest in Britain [J]. Forestry, 1999,72(3):191-205.
[8] 刘足根,姬兰柱,郝占庆,等.松果采摘对长白山自然保护区红松天然更新的影响[J].应用生态学报,2004,15(6):958-962.
[9] 刘妍妍.典型阔叶红松林木本植物的空间分布格局及其与生境的关联性[D].哈尔滨:东北林业大学,2014.
[10] 郑景明,罗菊春,长白山阔叶红松林结构多样性的初步研究[J].生物多样性.2003,11(4):295-302.
[11] BORDELON M A, MCALLISTER D C, HOLLOWAY R. Sustainable forestry oregon style[J]. Journal of Forestry,2000,98(1):26-32.
[12] 徐振邦,代力民,陈吉泉,等.长白山红松阔叶混交林森林天然更新条件的研究[J].生态学报,2001,21(9):1413-1420.
[13] 杨华,李艳丽,沈林,等.长白山云冷杉林幼苗幼树空间分布格局及其更新特征[J].生态学报,2014,34(24):7311-7319.
[14] 刘晓东,张彦雷.东折棱河森林经营所红松混交林的天然更新[J].东北林业大学学报,2009,37(9):8-11.
[15] 聂志文,布仁仓,李冰伦.红松存在/不存在数据的多尺度空间分布格局[J].生态学报,2015,37(3):810-818.
[16] 杨惠滨,国庆喜.地形与竞争因子对红松胸径与年龄关系的影响[J].生态学报,2016,36(20):6487-6495.
[17] 吴晓莆,朱彪,赵淑清,等.东北地区阔叶红松林的群落结构及其物种多样性比较[J].生物多样性,2004,12(1):174-181.
[18] 郭海燕,葛建平,李景文.中国红松林生态学研究文献概述[J].东北林业大学学报,1995,23(3):57-62.
[19] 国庆喜,葛剑平,马承慧,等.长白山红松混交林林隙状况与更新研究[J].东北林业大学学报,1998,26(l):4-7.
[20] 戎可,马建章.从灰松鼠竞争排斥红松鼠看外来物种入侵[J].生物学通报,2009,44(2):9-10.
[21] 宗诚,陈涛,马建章,等.凉水自然保护区松鼠和星鸦贮食生境选择差异[J].兽类学报,2007,27(2):105-109.
[22] 鲁长虎.动物与红松天然更新关系的研究综述[J].生态学杂志,2003.22(1):49-53.
[23] LI BING, FAN JINSHUAN, CHE XIAOQIANG. A review of studies on structural features, regeneration reatures and management of natural spruce-fir mixed stand of coniferous and broadleaved trees in China[J]. World Forestry Research,2012,25(3):43-49.
[24] 宋新章,张智婷,张慧玲,等.长白山森林不同演替阶段采伐林隙幼苗更新特征[J].江西农业大学学报,2010,32(3):504-509.
[25] 陈列,赵秀海,张赟.长白山北坡椴树红松林空间分布及其空间关联[J].北京林业大学学报,2009,31(3):6-10.
[26] 邹春静,韩士杰,张军辉.阔叶红松林树种间竞争关系及其营林意义[J].生态学杂志,2001,20(4):35-38.
[27] 何文利,李小凡,安慧君.阔叶红松林更新研究[J].内蒙古科技与经济,2004(23):31-32.
[28] 刘长海,倪淑清.天然红松林下的更新和林分类型的关系[J].林业科技情报,2007,39(2):26-27.
[29] 李东亮,王本俊.内蒙古大兴安岭栽培红松的研究[J].木本植物研究,2000,20(3):294-299.
[30] 巫志龙,陈金太,周新年,等.择伐强度对天然次生林乔木层6种优势种群生态位的影响[J].热带亚热带植物学报,2013,21(2):161-167.
[31] 陶大力,赵大昌,赵士洞,等。红松天然更新对动物的依赖性:一个排除动物影响的求过发芽实验[J]生物多样性,1995,3(3):131-133.
[32] 刘庆洪.红松阔叶林中红松种子的分布及更新[J].植物生态学报,1988,12(2):134-141.
[33] 王树力,葛剑平,徐继成,等.小兴安岭杨桦林下红松种群天然更新的格局与过程[J].东北林业大学学报,1993,21(5):7-12.
[34] 张国春.长白山阔叶红松林红松更新与生长规律[D].北京:北京林业大学,2011.
[35] 何学凯,张俊.黑龙江省东折棱河森林经营所次生林多样性研究[J].内蒙古林业调查设计,2008,31(3):122-124.
[36] 付士磊,刘兴双,宋晓东,等.长白山白然保护区阔叶红松林林隙更新状况[J].辽宁林业科技,2003(6):1-3.
[37] 刘长海,李玉玺,石艳丽.天然红松林结实与林分因子的关系[J].林业科技情报,2007,39(1):18-19.
[38] 李景文,刘传照,徐贵林,等.林冠下红松直播试验研究初报[J].东北林业大学学报,1984,12(S1):30-37.
[39] 聂佳旭,刘蓓蓓,杨慧,等.阔叶红松林中花鼠的集中贮藏洞穴特征[J].野生动物学报,2015,36(4):386-390.
[40] 张悦,易雪梅,王远遐,等.采伐对红松种群结构与动态的影响[J].生态学报,2015,35(1):38-45.
[41] 朱良军,金光泽,王晓春.典型阔叶红松林干扰历史重建及干扰形成机制[J].植物生态学报,2015,39(2):125-139.
[42] 包也,于大炮,周莉,等.择伐对阔叶红松林资源可持续利用的影响[J].生态学报,2015,35(1):31-37.
[43] 汤景明,翟明普.影响天然林树种更新因素的研究进展[J].福建林学院学报,2005,25(4):379-383.
[44] 肖笃宁,陶大力,徐振邦.大兴安岭北坡特大火灾对森林资源及环境的影响[J].生态学杂志,1988,7(增刊):5-9.
[45] 张海军,张淑兰,王长宝.小兴安岭红松种群天然更新及影响因子的探讨[J].林业资源管理,2015(2):150-153.
Natural Regeneration of Primitive Korean Pine Forests in Response to the Variation in Continuous Small Elevation Gradient//
Zhang Fanbing, Long Ting
(Beijing Forestry University, Beijing 100083, P. R. China); Jin Wenbin(Training Offices of Armed Police in Beijing Forestry University); Li Jingwen(Beijing Forestry University)//
Journal of Northeast Forestry University,2017,45(5):1-10.
With natural regeneration of Korean pine seed stands in NO.56 Forest compartment of Dong Zhelenghe Nature Reserve belonging to Xiaoxing’an Mountains, three levels in 12 plots included the lower slopes in forest compartment (Plots A, B, C, D, E, F), the lower-middle slopes (Plots G, H, I) and the central slopes (Plots J, K, L) were sited based on small elevation gradient (changed from three to fifteen meters). Without human disturbance, as the elevation gradient was increased, conifers replaced companion broadleaves gradually in the Tachiki layer. Gap between conifers and companion broadleaves was reduced gradually. The number of update trees was increased including Korean pine. Under human disturbances, some plots of understory vegetation cover were more than 10% on average, and had a bad update. In this scenario, Heliousphytes Broadleaves such as Acertegmentosum Maxim, Acerukurunduense Trautv had a smaller advantage on regeneration than Coniferous, and Korean pine had a few update young trees. Korean pine is a dominant species in this forest community although it has a few Tachiki numbers and increased slowly. Korean pine has unanimous trends in importance value and percent area, both them showed a steady growth with altitude increasing. Elevation changing has no influence in spatial distribution pattern of Tachiki layer for Korean pine. Korean pine showed randomly distributed or no regeneration in lower altitudes, while aggregated distribution in medium altitude, and a randomly distribute in advantage layer at last by intense natural succession.
Xiaoxing’an Mountains; Primitive Korean pine forest; Continuous small gradient elevation changes; Natural regeneration; Spatial pattern
张凡兵,男,1992年10月生,北京林业大学林学院,硕士研究生。E-mail:beilinjinwb@163.com。
李景文,北京林业大学林学院,教授。E-mail:lijingwen@bjfu.edu.cn。
2016年10月29日。
S791.247.02
1)国家“十三五”重点研发计划课题(2016YFC0503106)。
责任编辑:王广建。
