油茶暹罗刺盘孢菌群体遗传结构分析
2017-06-05周国英蒋越西蒋仕强刘君昂
李 杨, 李 河, 周国英, 蒋越西, 蒋仕强, 刘君昂*
(1. 中南林业科技大学,森林有害生物防控湖南省重点实验室, 长沙 410004; 2. 中南林业科技大学,经济林培育与保护省部共建教育部重点实验室, 长沙 410004)
油茶暹罗刺盘孢菌群体遗传结构分析
李 杨1,2, 李 河1,2, 周国英1,2, 蒋越西1, 蒋仕强1, 刘君昂1,2*
(1. 中南林业科技大学,森林有害生物防控湖南省重点实验室, 长沙 410004; 2. 中南林业科技大学,经济林培育与保护省部共建教育部重点实验室, 长沙 410004)
暹罗刺盘孢菌是油茶炭疽病病原之一,在我国多个油茶产区均有分布。研究油茶暹罗刺盘孢菌群体遗传结构可为全面、有效防治油茶炭疽病害提供理论依据。本研究对分离自海南、江西、湖南、广西4省(自治区)6个地区暹罗刺盘孢菌菌株的ITS、CAL和GAPDH 3个基因的序列进行群体遗传结构分析。根据拼接的上述3个基因的序列,57个暹罗刺盘孢菌菌株可定义为13个单倍型,其中单倍型H7为主要单倍型,分布于本研究所涉及的所有地区。病菌不同地理种群间的遗传分化较大,AMOVA分析显示,遗传变异主要发生在种群内,病菌未经历过大规模的种群扩张。研究结果表明油茶炭疽病原暹罗刺盘孢菌种群具有丰富的遗传多样性。
暹罗刺盘孢菌; 地理种群; 多基因序列; 群体遗传结构
油茶炭疽病是油茶树的主要病害之一,在我国油茶主产区均有发生,尤其在湖南、江西、广西和海南等省(自治区),发生面积广,危害严重,造成了重大经济损失[1-2]。炭疽病主要造成油茶树落叶、落蕾及落果,导致茶籽产量降低,重病产区可减产50%及以上[3]。
本研究团队已鉴定报道的油茶炭疽病病原菌主要有果生刺盘孢菌Colletotrichumfructicola、暹罗刺盘孢菌C.siamense、胶孢炭疽菌C.gloeosporioides、博宁炭疽菌C.boninense、山茶刺盘孢菌C.camelliae以及尚未确定分类地位的炭疽属真菌1~3种,其中C.fructicola和C.siamense是优势致病菌[4-9]。李河等[4]对C.fructicola的群体遗传进行了深入研究,而C.siamense目前仅局限于种名的确定,尚未对其生物学特性、遗传多样性等方面进行研究,这在一定程度上增加了该病菌引起的油茶炭疽病的防治难度。
由于炭疽属真菌种间差异较小,仅根据ITS序列进行炭疽病菌的鉴定,错误率高达80%以上[10],而多基因序列的引入,大大提高了对炭疽属真菌的鉴定准确率,并得到了广泛的应用[11-13]。本研究利用ITS-CAL-GAPDH 3基因序列对我国6个油茶主产区暹罗刺盘孢菌的群体遗传结构进行研究,以期分析同一地区C.siamense种群的个体变异、阐明该病菌群体遗传分化与基因流等问题,为该病害流行机制的研究提供理论依据,这对于有效防治该病菌引起的油茶炭疽病具有重要意义。
1 材料与方法
1.1 材料
菌株采集:样品采自海南、湖南、江西和广西的6个地区,病原菌分离方法参考方中达[14]的组织分离法,取病健交界处组织进行分离,分离菌株用PDA培养基纯化培养后4℃冰箱保存备用。经回接试验、形态和多基因分子鉴定,共获得57株暹罗刺盘孢菌。
主要试剂:DNA快速提取试剂盒Fast DNA Kit,美国MPBIO公司;2×TaqPCR Master Mix,天根生化科技(北京)有限公司。
主要仪器:快速核酸提取仪,美国MPBIO公司;5145D离心机,德国Eppendorf公司;ABI 9700 PCR仪,美国应用生物系统公司。
1.2 方法
1.2.1 菌株DNA提取
用灭菌牙签刮取0.5 g菌丝,使用DNA快速提取试剂盒提取DNA,提取方法参见说明书,-20℃保存。
1.2.2 基因选择、PCR扩增及测序
基因选择及目的片段扩增与测序参照 Weir 等[11]的方法、反应体系及条件。对所有分离筛选获得的菌株,选择核糖体转录间隔区(internal transcribed spacers,ITS)、3-磷酸甘油醛脱氢酶基因(glyceralde-hyde-3-phosphate dehydrogenase,GAPDH)和钙调蛋白基因(calmodulin,CAL)进行扩增。相应的PCR扩增引物和退火温度见表1。PCR反应产物测序由上海铂尚生物技术有限公司完成。

表1 本试验所用引物和退火温度
1.2.3 数据分析
DNA序列采用BioEdit软件[15]进行编辑加工并辅以手工调整。使用MEGA6.0对核苷酸组成、变异位点等信息进行分析。不同地区炭疽病菌群体的ITS-CAL-GAPDH单倍型分析利用软件GenALEx6完成,同时对不同群体的单倍型多样性进行分析。运用DnaSP5.0[16]计算暹罗刺盘孢菌的3个基因拼接序列的核苷酸多样性(nucleotide diversity,Pi)、不同地区群体间遗传分化指数(fixation index,Fst),并对所有病菌群体序列的单倍型错配进行分析;使用TCS1.21[17]构建暹罗刺盘孢菌单倍型的网络图。采用MEGA 6.0构建系统发育树:将依据ITS-CAL-GAPDH 3个基因推导出的单倍型序列与GenBank中炭疽属真菌菌株的ITS、CAL、GAPDH基因序列进行分析比对,构建近邻归群(neighbor-joining,NJ)系统发育树,利用自展法(bootstrap,1 000次重复)检验各分支的置信度[18]。
2 结果与分析
2.1 暹罗刺盘孢菌单倍型地理分布及其多样性
ITS-CAL-GAPDH 3个基因拼接序列共1 385个碱基,核苷酸组成分析表明:A+T含量46.0%略低于C+G含量54.0%;共发现22个变异位点,其中,简约信息位点20个,单突变位点2个;57条暹罗刺盘孢菌ITS-CAL-GAPDH 3个基因序列共定义了13种单倍型,各单倍型在不同种群中的分布见表2。在所有单倍型中,单倍型H7为主要单倍型,分布于所有地区,占总序列数量的56.1%,结果表明种群间存在基因交流。此外,江西分宜和湖南马家河含有单倍型种类最多,均含有5种单倍型,而海南屯昌含有单倍型的数量最少,仅有2种单倍型。除海南屯昌外,其他地区均含有自己独特的单倍型,如海南五指山含有H5和H10,江西分宜含有H1、H3及H6,这表明每个群体内存在独立进化,群体间存在一定程度的遗传分化。
油茶炭疽病菌不同地理种群的单倍型多样性在0.219~0.735之间,其中湖南马家河种群的单倍型多样性最高,为0.735;海南屯昌种群的单倍型多样性最低,为0.219。各地理种群具有较高的单倍型多样性,这可能与当地气候或地理环境有关。暹罗刺盘孢菌的ITS-CAL-GAPDH 3个基因拼接后核苷酸多样性(Pi)也处于较高水平(表2),除海南屯昌、湖南天际岭种群的Pi分别为0.000 54、0.000 55外,其他4个种群的核苷酸多样性在0.001 01~0.002 32之间。结果表明,暹罗刺盘孢菌在6个地区具有丰富的遗传多样性。

表2 暹罗刺盘孢菌样品的地理分布及单倍型多样性
2.2 单倍型网络分析及种群扩张分析
由病菌ITS-CAL-GAPDH 3个基因拼接序列单倍型网络结构图(图1)可知,暹罗刺盘孢菌各单倍型并未依采样地点形成各自的分支,而是以单倍型H7为辐射中心,呈现出一个非典型星状发散图,H7可能为原始单倍型。我们从图中还可以看出大部分单倍型通过一次或数次突变与原始单倍型H7相连。此外,由暹罗刺盘孢菌不同种群3基因拼接序列的核苷酸不配对分布图(图2)可知,分布曲线呈多峰型曲线,说明油茶炭疽病原暹罗刺盘孢菌可能未经历过大规模地种群扩张或“瓶颈效应”。
2.3 暹罗刺盘孢菌的AMOVA分析
由表3可知暹罗刺盘孢菌种群变异主要发生在种群内,占总变异的90%,而种群间的变异仅占10%,这说明病菌不同地理种群间基因交流频繁。暹罗刺盘孢菌不同地理种群间和种群内的变异差异显著大于0,表明暹罗刺盘孢菌不同地理种群内的遗传变异显著(P<0.05)。
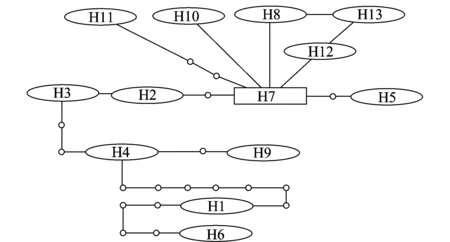
图1 暹罗刺盘孢菌单倍型网络图Fig.1 Parsimony network of the haplotypes of Colletotrichum siamense
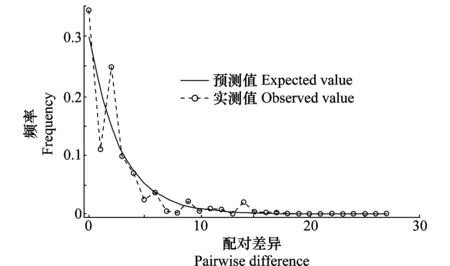
图2 暹罗刺盘孢菌群体核苷酸不配对分布Fig.2 Mismatch distribution of Colletotrichum siamense population
2.4 暹罗刺盘孢菌不同地理种群间遗传分化分析
Wright[19]于1978年提出遗传分化指数(Fst)概念:当遗传分化指数Fst≤0.05时,群体间遗传分化很弱;当0.05

表3 暹罗刺盘孢菌种群内和种群间的分子变异分析
海南五指山与海南屯昌、湖南天际岭、广西南宁之间的Fst在0.05~0.15之间;广西南宁与江西分宜、湖南天际岭、湖南马家河之间的Fst也在0.05~0.15之间,表明这些种群间存在一定程度的遗传分化。江西分宜与海南五指山、海南屯昌、湖南天际岭之间遗传分化较大,Fst在0.164 97~0.180 01之间;湖南马家河与海南五指山、海南屯昌、湖南天际岭之间遗传分化较大,Fst在0.220 47~0.240 60之间。种群间存在较大的遗传分化,这可能是地理环境、气候等因素造成的。Mantel测试显示,暹罗刺盘孢菌地理距离与种群间遗传分化之间无显著的线性关系(R2=0.059,P=0.20)。

表4 暹罗刺盘孢菌不同地理种群间遗传分化值
2.5 基于ITS-CAL-GAPDH 3基因序列的系统发育树分析
暹罗刺盘孢菌ITS-CAL-GAPDH 3基因序列单倍型的系统发育树见图3,所有种群单倍型散乱地分布在系统树中,同一种群的单倍型序列并未聚在一起;不同种群的独特单倍型并未完全形成自己特有的进化支。此外,13种单倍型与GenBank中的模式菌株(表5)C.siamense(ICMP18578、ICMP18642)以很高的支持率聚为一支,也验证了从6个地区分离到的57个菌株为暹罗刺盘孢菌。
3 讨论
遗传多样性是生物多样性的基础,研究病原菌遗传结构可为明确病菌对不同环境的适应能力提供理论基础。一个物种的遗传多样性越高,其对不同生存环境的适应力也就越强[20]。李河等[4]分析了我国油茶果生刺盘孢菌种群的遗传多样性,发现该病菌在我国油茶产区分布十分广泛,单倍型多样性较丰富,存在一定的遗传分化现象。本研究利用ITS-CAL-GAPDH 3基因序列对油茶暹罗刺盘孢菌群体遗传结构进行研究,发现该病菌广泛分布在我国油茶产区,单倍型多样性也较为丰富,存在一定的遗传分化现象,这揭示了油茶炭疽病在我国发生面积广和危害程度重的遗传基础。

表5 构建N-J系统树所用的模式菌株及其GenBank登录号
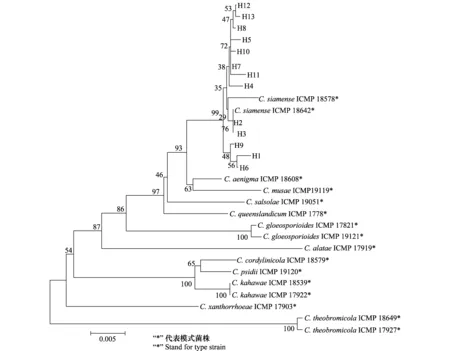
图3 采用邻接法构建的ITS-CAL-GAPDH基因序列系统树Fig.3 Phylogenetic tree of ITS-CAL-GAPDH genes sequences based on N-J method
很多学者研究发现,植物病原菌在进化过程中存在基因流,同时种群间存在遗传变异。王玲等[21]对浙皖鄂地区水稻纹枯病菌5个种群的遗传结构进行了研究,发现水稻纹枯病菌种群变异主要发生在种群内,病菌群体间基因交流频繁。这与本研究中暹罗刺盘孢菌种群变异也主要发生在种群内,种群间变异仅占10%,且病菌在不同地区间存在基因传递的结论相同。朱丹雪等[22]对我国油茶果生刺盘孢菌10个种群的遗传结构进行研究,发现单倍型H5分布于每一个种群中,为优势单倍型;Pascual等[23]也发现在不同玉米纹枯病菌种群间存在相同单倍型。本研究发现优势单倍型H7在供试的每个地理种群中均有分布,57个样本中有32个含有单倍型H7,占总样品数量的56.1%,这说明暹罗刺盘孢菌种群间的基因流未受地理差异的影响,其优势单倍型可在不同地区间进行传播,导致毒性较强的基因型在种群内维持下去;另外,研究发现除海南屯昌外,其他5个地理种群均含有至少一个独特的单倍型,说明该病菌在长时间进化过程中为了适应环境变化或寄主选择,形成了自己独特的单倍型,丰富了种群的遗传多样性,增加了对该病害的防治难度。
易克贤等[24]对43株中国柱花草炭疽菌菌株的基因组DNA进行了分析,并和国外菌株进行综合聚类分析,发现同一地区的菌株聚成一类,表明不同地区的柱花草炭疽菌具有各自独立的进化历程。在本研究中,同一地理种群的病菌单倍型并未聚在一起,而是散乱地分布在系统树中,表明没有地理特异性,且病菌地理距离与种群遗传分化之间无显著线性关系。
本研究中样品来自我国6个油茶种植区,不同地区样品数存在一定差异,但我们的结果已能够清晰地显示我国油茶主产区中暹罗刺盘孢菌具有丰富的遗传多样性,揭示了该病害在油茶主产区广泛发生和危害严重的理论基础。另外,研究发现不同地理种群间基因交流频繁,同时病菌种群间存在一定遗传分化现象,这可能是病菌为了适应环境变化而不断进化的原因。下一步将扩大采样范围,增加病菌数量,同时应用其他分子标记,以获取暹罗刺盘孢菌更多的遗传信息,为阐明该病菌引起的病害流行机制提供理论依据。
[1] 靳爱仙, 周国英, 李河. 油茶炭疽病的研究现状、问题与方向[J]. 中国森林病虫, 2009, 28(2): 27-31.
[2] 陈守常, 田泽君, 郭隆锡. 油茶炭疽病的发生与蔓延[J]. 植物保护学报, 1965, 4(3): 207-218.
[3] 罗万周, 罗万业. 油茶炭疽病及其防治方法[J]. 农技服务, 2007,24(6): 70.
[4] Li He, Zhou Guoying, Liu Junang, et al. Population genetic analyses of the fungal pathogenColletotrichumfructicolaon tea-oil trees in China [J].PLoS ONE, 2016,11(6): 1-24.
[5] 李河, 李杨, 蒋仕强, 等. 海南省不同寄主植物炭疽病菌侵染油茶研究[J]. 植物保护, 2016,42(4):64-70.
[6] 李河, 周国英, 徐建平.一种新油茶炭疽病原多基因序列鉴定[J].植物保护, 2015, 41(2): 92-96.
[7] 李河, 周国英, 徐建平, 等. 一种油茶新炭疽病原的多基因系统发育分析鉴定[J]. 植物保护学报, 2014, 41(5): 602-607.
[8] 汤铱泠, 周国英, 李河, 等. 多基因序列鉴定油茶炭疽病原Colletotrichumboninense新种[J]. 热带作物学报, 2015, 36(5): 972-977.
[9] 李杨,李河,周国英,等. 油茶新炭疽病原Colletotrichumcamelliae鉴定及致病性测定[J]. 生物技术通报,2016,32(6):96-102.
[10]Crouch J A, Clarke B B, Hillman B I. What is the value of ITS sequence data inColletotrichumsystematics and species diagnosis? A case study using the falcate-sporedgraminicolousColletotrichumgroup [J].Mycologia, 2009, 101(5): 648-656.
[11]Weir B S, Johnston P R, Damm U. TheColletotrichumgloeosporioidesspecies complex [J]. Studies in Mycology, 2012, 73(1): 115-180.
[12]杨友联. 中国云南、贵州、广西炭疽菌属真菌多基因分子系统学研究[D].武汉: 华中农业大学, 2010.
[13]朱英芝, 廖旺姣, 邹东霞, 等. 广西油茶炭疽病病原菌鉴定及生物学特性[J]. 植物保护学报, 2015, 42(3): 382-389.
[14]方中达.植病研究方法[M]. 第3版.北京: 中国农业出版社, 1998: 122-145.
[15]Hall T A.BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT [J]. Nucleic Acids Symposium Series, 1999(41): 95-98.
[16]Librado P, Rozas J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data [J]. Bioinformatics, 2009, 25(11): 1451-1452.
[17]Clement M, Posada D, Crandall K A.TCS: a computer program to estimate gene genealogies[J]. Molecular Ecology, 2000, 9(10): 1657-1660.
[18]Tamura K, Nei M, Kumar S. Prospects for inferring very large phylogenies by using the neighbor-joining method[J]. Proceedings of the National Academy of Sciences of United States of America, 2004, 101(30): 11030-11035.
[19]Wright S. Evolution and the genetics of populations. Volume 4: Variability within and among natural populations [M]. Chicago: The University of Press, 1978: 325-327.
[20]袁娟, 张其中, 李飞, 等. 铜鱼线粒体控制区的序列变异和遗传多样性[J]. 水生生物学报, 2010, 34(1): 9-19.
[21]王玲, 黄雯雯, 黄世文, 等. 浙皖鄂地区水稻纹枯病菌5个种群的遗传结构分析[J]. 生态学报, 2010, 30(20): 5439-5447.
[22]朱丹雪,周国英,徐建平,等.果生刺盘孢菌Colletotrichumfructicola群体遗传结构研究[J].菌物学报, 2015, 34(3): 366-374.
[23]Pascual C B, Toda T, Raymondo A D, et al. Characterization by conventional techniques and PCR ofRhizoctoniasolaniisolates causing banded leaf sheath blight in maize [J]. Plant Pathology, 2000, 49(1): 108-118.
[24]易克贤, 黄俊生, 刘国道, 等.中国柱花草炭疽病原菌遗传多态性的RAPD分析[J]. 微生物学报, 2003, 43(3): 379-387.
(责任编辑:田 喆)
Population genetic structure ofColletotrichumsiamenseon tea-oil trees
Li Yang1,2, Li He1,2, Zhou Guoying1,2, Jiang Yuexi1, Jiang Shiqiang1, Liu Jun’ang1,2
(1.Hunan Provincial Key Laboratory for Control of Forest Diseases and Pests, Central South University of Forestry andTechnology, Changsha 410004, China; 2.Key Laboratory of Cultivation and Protection for Non-wood Forest Trees ofMinistry of Education,Central South University of Forestry and Technology, Changsha 410004, China)
Colletotrichumsiamenseis one of the primary anthracnose pathogen ofCamelliaoleiferafrom most producing areas in China. To clarify the population genetic structure will provide theoretical basis for the effective control of plants anthracnose. The population genetic structure of 57C.siamensestrains collected from Hainan, Jiangxi, Hunan and Guangxi of China were analyzed in this paper. According to the assembled sequences of ITS, CAL and GAPDH genes, the 57C.siamensestrains were defined into 13 haplotypes, and the haplotype H7 was found dominant and distributed in all collection sites. There is large genetic differentiation between different geographical populations. AMOVA analysis showed that the genetic variation mainly took place within population, and the pathogen doesn’t experience large-scale population expansion. The result showed that the natural population ofC.siamensehad rich genetic diversity.
Colletotrichumsiamense; geographical population; multiple gene sequences; population genetic structure
2016-03-21
2016-08-25
国家林业公益性行业科研专项(201304403);国家自然科学基金(31570641)
S 435.65
A
10.3969/j.issn.0529-1542.2017.03.008
* 通信作者 E-mail:csuftfp@163.com
