3种木棉光合特性及碳汇能力研究
2017-06-01吴金山王亚沉徐诗涛赵庆杰李英英于旭东
吴金山, 王亚沉, 徐诗涛, 赵庆杰, 李英英, 于旭东
(海南大学 热带农林学院, 海南 儋州 571737)
3种木棉光合特性及碳汇能力研究
吴金山, 王亚沉, 徐诗涛, 赵庆杰, 李英英, 于旭东*
(海南大学 热带农林学院, 海南 儋州 571737)
以美丽异木棉、爪哇木棉、海南木棉叶片为测试材料,利用3051D光合作用测定仪测定3种木棉光合作用日变化规律,分析其相关性,并通过光合速率计算固碳量,从而得出三者碳汇能力.结果表明:3种木棉光合速率(Pn)日变化基本都呈“M”型双峰曲线,存在明显的“午休”现象;3种木棉蒸腾速率(Tr)均呈单峰曲线变化,12∶00~14∶00时达到最高,其中爪哇木棉 Tr 在出现的时间和峰值上都早和高于另外两个品种;3种木棉单位日固碳量在 0.59~0.68 g·m-2·.d-1之间,其中美丽异木棉和海南木棉均为0.68 g·m-2·d-1,爪哇木棉为0.59 g·m-2·d-1.3种木棉年固碳量在214.49~246.11 g之间,其中爪哇木棉比另两品种碳汇能力弱,约低12.8%.通过聚类分析表明,美丽异木棉和海南木棉具有极高的相似性.本研究可为以后园林绿化优良树种选育工作提供参考.
木棉; 光合特性; 碳汇; 测定
碳汇,一般是指从空气中清除 CO2的过程、活动、机制.自 1860 年工业革命以来,大气中 CO2浓度已从原来的 280 μmol/mol 增加到370 μmol/mol,目前每年仍增加约 1.5 μmol/mol,预计到本世纪中期, 大气的 CO2浓度会增加到目前的两倍[1].CO2浓度增加带来的温室效应也逐渐引起世界各国的重视,碳汇也随之作为一个较新的课题逐渐兴起,它是主要国家通过对陆地生态系统的有效管理尤其是加强碳汇林的建设来提高固碳潜力,利用所取得的成效抵消本国或者其他国家的碳减排份额,从而获得一定经济收益的途径自国家林业局成立碳汇管理办公室后,我国造林与再造林碳汇项目的实施逐渐在清洁发展机制下合理有序地发展.碳汇一般主要指森林吸收并储存的 CO2量,或者是森林吸收并储存 CO2的能力.历次森林资源清查表明,我国森林资源蓄积一直处于增长状态,我国已在西南、华北等一些地区陆续开展了碳汇项目.园林绿化树种不仅起到美化城市环境作用,其碳汇能力对 CO2带来的温室效应也起到一定的缓解作用[2-4],近年来也逐步被人们所重视.但我国目前在城市绿地生态方面仍研究较少.
植物对 CO2吸收利用主要是通过光合作用来完成的,其与植物固碳能力的强弱有着直接的关系.因此提高植物叶片光合效率是提高植物碳汇能力的重要途径.近年来,随着遗传育种学科的发展,高光效育种已成为育种工作者关注的主要问题.同一植物不同品种之间由于遗传因子的影响,光合潜能差异也很大.
木棉在我国有2 000多年的栽培历史[5],是一种难得的生态林、景观林、经济林树种.其主要分布在热带及亚热带地区,目前是广州市、攀枝花市市花,我国海南昌江还有中国“木棉之乡”之称[6].近年来,随着种植数量和栽培面积的不断增加,木棉树种在建设低碳及生态城市中的作用越来越重要[7].木棉树体高大,树型各异,不同品种各不一样,其光合生理特性也存在较大的差异.目前国内外有关木棉植物不同品种间高光效研究还未见报到.本文主要利用 3051D 光合速率仪对木棉科3个树种(海南木棉、爪哇木棉、美丽异木棉)进行了的光合特性研究,揭示3个树种的基本光合生理特征和规律,为高光效育种、选育优良园林绿化品种,加快良种的繁育与推广提供参考.同时分析了3个单绿化树种的碳汇能力,从而得知目标树种在建设低碳、生态城市中的作用,为城市绿地的建设规划与碳汇交易市场建设提供理论参考.
1材料与方法
1.1试验材料
供测试美丽异木棉(Chorisiaspeaiosa)、爪哇木棉(Ceibapertandra)、海南木棉(Bombaxcelibate)来自海南省儋州市热科院及热带农林学院园区.选择生长良好无病虫害,以长势基本一致,色泽正常成年植株上部3片叶为材料,美丽异木棉树高约 20 m,直径 1.4 m、爪哇木棉高约 18 m,直径 1.2 m、红花木棉高约 16 m,直径 1.1 m.
1.2试验设计与方法
利用 3051D 光合作用测定仪(来自浙江托普仪器有限公司)对选取的美丽异木棉、爪哇木棉、海南木棉3类目标树种同一树上部不同叶片进行瞬时光合速率测定.于 2015 年 9 月植物叶片全面展时,选择连续3个无风、阳光充足的全天进行,从8∶00~18∶00,每1 h监测1次,每叶片一次记录5个点,每个时间点重复3次(树叶监测叶面积为11 cm2,系统容积0.06 L),然后求平均值得出光合速率日变化.在测定前首先对仪器进行预热30~60 min,待探头器室 CO2稳定后再进行测定,得到不同树种的净光合速率(Pn)、蒸腾速率(Tr)、胞间 CO2浓度(Ci)、光合有效辐射(PAR)、大气 CO2浓度(Ca)、空气相对湿度(RH)、大气温度(Ta)、气孔导读(Gs)及其他环境因子参数,结合实验结果确定不同树种在不同气候条件下的 CO2固定效率.
1.3碳固定量
净日同化量是昼浄光合量与夜间暗呼吸量的差,浄日同化量是植物对生态的实际贡献量.植物的光合速率和呼吸速率由于同时受到生理和生态环境因子的影响,全天各个时段速率值都在变化,采用植物浄日同化量作为评价的基础数据较为准确.因此根据净光合速率可得出植物的净同化量[6].
同化量计算公式为:
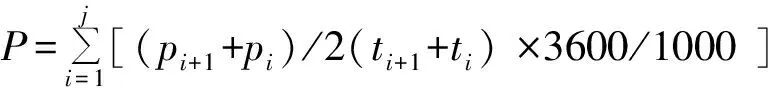
(1)
式中,P为同化总量, 单位为 mmol,pi为初测点瞬时光合速率,pi+1为下一测点瞬时光合速率,单位为μmol·m-1·s-1;ti为初测点时间,ti+1为下一测点时间,单位 h,j为测试次数,3 600 指每小时为 3 600 s,1 000 指 1 mmol 为 1 000 μmol.用测定日的同化总量换算为测定日固定量为:
WCO2=P×44/1000,
(2)
式中,44 为CO2的摩尔质量,单位为 g·mol-1,WCO2为单位面积的叶片固定CO2的摩尔质量, 单位为g·m-2·d-1.
植物单位绿地面积上的固碳能力按公式(3)计算:
QCO2=WCO2×LAI,
(3)
式中,QCO2为单位绿地面积固碳量,单位 g·mol-2;LAI为叶面积指数.
1.4数据处理
所有数据主要利用Excel2003进行整理分析并绘制图表,方差及相关性分析主要利用SPSS11.0软件进行分析处理.
2结果与分析
2.1环境因子日变化规律
木棉光合蒸腾特性主要测定PAR、RH、Ta和Ca共4个主要环境因子.如图1,PAR日变化呈单峰曲线,在中午11∶00~12∶00时达到最大值,12∶00时为全天最高值1 812.6 μmol·m-1·s-1),12∶00时后开始呈下滑趋势.而全天空气中CO2呈“U”字型变化,在中午12∶00~13∶00时达到最低,为329.31 μmol/mol,8∶00时和18∶00时均为两端最高,分别为466.29 μmol/mol和467.98 μmol/mol.大气温度和湿度全天变化均呈单峰曲线(如图2),但温度变化呈上升再下降的趋势,在中午12∶00~14∶00时达到最高值,为35.5 ℃.而在全天中大气湿度与温度变化呈相反变化趋势,在11∶00~14∶00时处于低谷,其中13∶00时达到最低为62.49% RH,在8∶00时和18∶00时则均较高,都在80% RH以上.上述现象可能与海南四周环海,离赤道比较近,充足的降水量及高湿环境有关.

图1 PAR与CO2的日变化Fig.1 Diurnal change of PAR、CO2
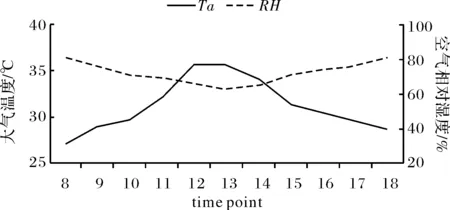
图2 RH与Ta的日变化Fig.2 Diurnal change of RH、Ta
2.2光合速率日变化特征
植物光合作用随外界环境因子的变化而变化,而光合速率是对光合作用强弱的一种反应,也会因环境因素的变化而发生波动[8-9].从图3可看出3种木棉树种Pn日变化规律基本呈相同“M”型双峰曲线变化趋势,存在明显的“午休”现象,因此也说明3树种都受到不同程度的光抑制作用,但3种木棉Pn日变化差异不明显.8∶00~10∶00时3种木棉净光合速率呈上升趋势,10∶00时美丽异木棉达到最大值为15.1 μmol·m-2·s-1,而在下午13∶00时海南木棉和爪哇木棉达到最大值分别为13.4 μmol·m-2·s-1与12.1 μmol·m-2·s-1,随之下降;14∶00时美丽异木棉再次出现小高峰为13.3 μmol·m-2·s-1,之后开始下降.由Pn全天变化趋势可知,3种木棉在15∶00~18∶00时光合速率呈下降趋势.而到下午17∶00时3木棉Pn急剧下降,18∶00时净光合速率基本停止.美丽木棉“M”型曲线变化表现最为明显,其他两种较弱.

图3 3种木棉光合速率日变化趋势Fig.3 Three kinds of kapok Pn change trend
2.3蒸腾速率日变化特征
Tr的大小在一定程度上反映植物调节水分及适应逆境能力,同时也与植物光合速率密切相关[10],从图4可知,3种木棉Tr均呈单峰曲线变化,12∶00~14∶00时达到最高,美丽异木棉、爪哇木棉、海南木棉分别为1.66 μmol·m-2·s-1、1.78 μmol·m-2·s-1和1.62 μmol·m-2·s-1,而爪哇木棉 Tr 峰值在出现的时间上早于另外两个品种,峰值也高于另两品种,表明爪哇品种在高温、强光的情况下能更好适应环境,进行光合作用,降低高温对植株的生理性伤害.

图4 3种木棉蒸腾速率日变化趋势Fig.4 Three kinds of kapok Tr change trend
2.4气孔导度与胞间CO2浓度日变化特征
气孔是水分和 CO2气体交换的重要通道,而Gs则表示气孔的张开程度,是决定植物光合作用和蒸腾作用的重要因素,是连接生态系统碳循环的结合点.在一定范围内,叶面CO2浓度增加光合速率增强[11],超过一定浓度,气孔导度则变小.由图5可知,3种木棉Gs日变化呈单峰趋势变化,在上午10∶00时均达到最大值,爪哇木棉在3种木棉中最高.
由图6可知,3种木棉Ci呈“U”型变化趋势,在早上8∶00和下午18∶00时数值都比较最高,在12∶00~13∶00时为最低,其中最低的美丽异木棉在13∶00时值为242.08 μmol·mol-1.由图3、图5可得,胞间 CO2浓度与光合速率呈相反的单峰曲线变化趋势.
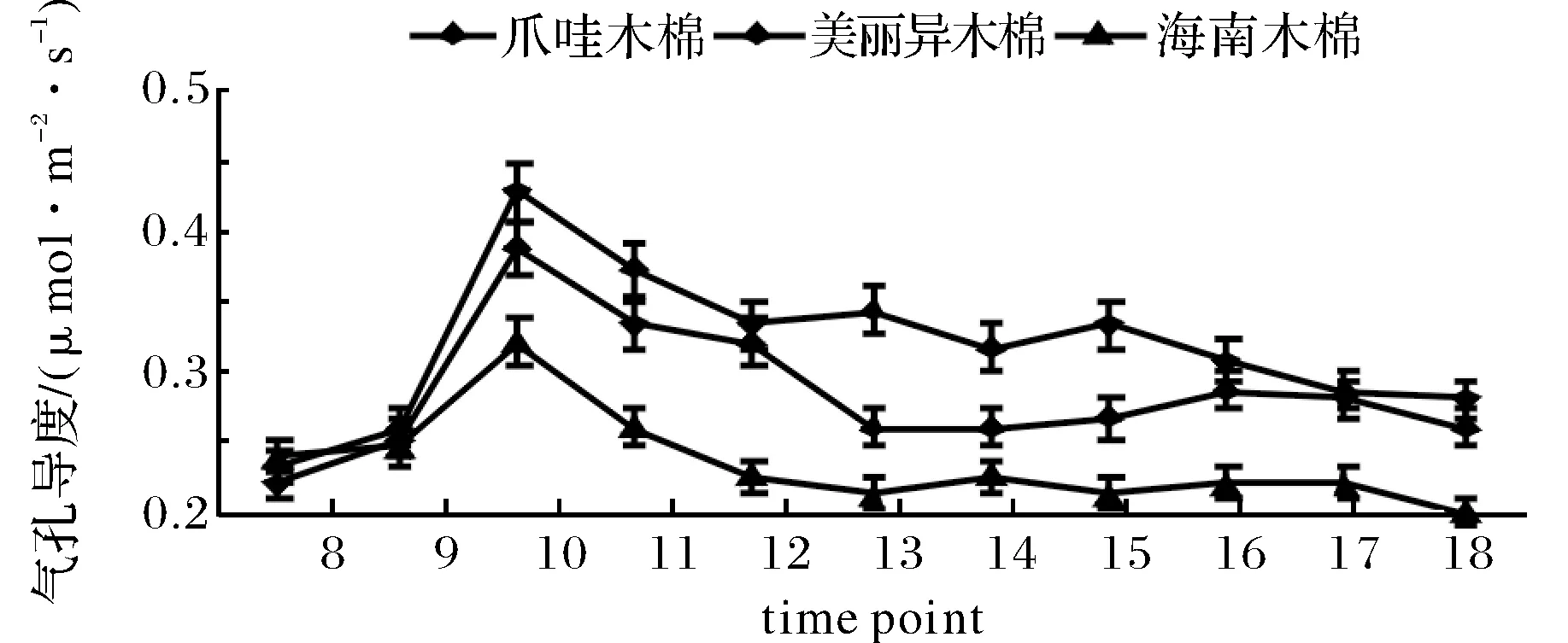
图5 Gs日变化Fig.5 Diurnal change of Gs

图6 Ci浓度日变化Fig.6 Diurnal change of Ci
2.5光合日变化与生理生态因子相关性分析
由表1、表2、表3可知,海南木棉的PAR与Ta、Tr、Pn均呈极显著正相关,同时与Ca、RH、Ci呈现为负相关,这与上文所述日变化趋于一致.Ca与Ta、Tr呈现为负相关,与Ci表现为极显著正相关,同时,进一步表明Ci随着不同时间段Ta增加或减少而表现出相应的变化.RH与Ta、Tr表现为显著负相关性,与Ci呈现为显著正相关.结果表明:Ci随着RH的变化而增加或减少.Ta与Ci表现为显著负相关,与Tr为显著正相关性,说明大气温度越高Ci越低,相反,Tr是随着Ta的升高而加快,这与木棉的气孔开张规律有关.Ci与Tr、Pn表现为负相关性,说明当Tr和Pn提高时,Ci的呈现下降趋势.Tr与Pn呈现为显著正相关性.
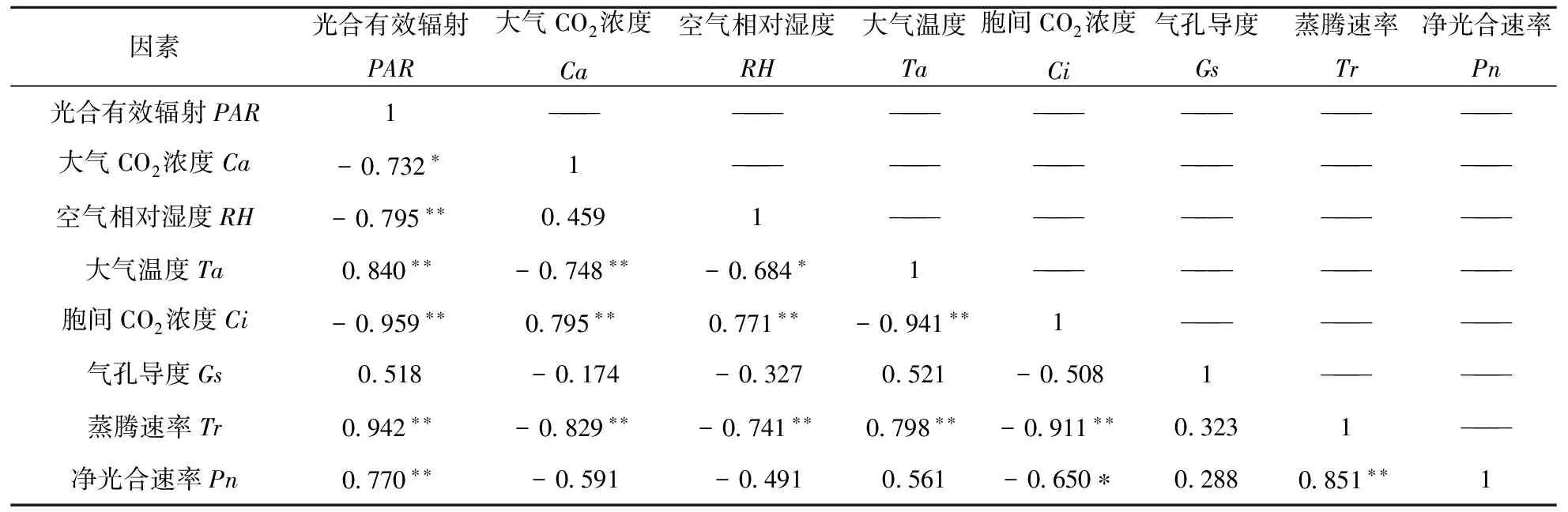
表1 海南木棉光合日变化与生理生态因子的相关分析
注:*为显著相关,p<0.05;**为极显著显著相关,p<0.01.

表2 爪哇木棉光合日变化与生理生态因子的相关分析
注:*为显著相关,p<0.05;**为极显著显著相关,p<0.01.

表3 美丽异木棉光合日变化与生理生态因子的相关分析
注:*为显著相关,p<0.05;**为极显著显著相关,p<0.01.
爪哇木棉、美丽异木棉在PAR、Ta及其另外几个生理因子相互之间的相关性与海南木棉趋于一致.主要表现为PAR与Ta、Tr、Pn显著相关性上没有海南木棉表现的程度高.说明海南木棉对光合作用的利用率高于另外两木棉.爪哇木棉Ca与Ci相关性比海南木棉高,美丽异木棉的表现则不明显,有可能与植物品种组织结构有关,需要进一步探究.RH、Ta、Ci、Tr与其他因子相关性与海南木棉的结果相同.
2.63种木棉单位叶面积固碳量分析与碳汇能力的聚类
植物固碳能力大小对整这个生态环境的影响起着重要的作用[12].而植物年固碳能力通常用一年中的光合作用、固碳量表示较为准确.根据植物净同化量计算公式(见上实验方法1.3),由木棉净光合速率可得出固碳量.由表4可知,3木棉单位日固碳量在0.59~0.68 g·m-2·d-1之间,其中爪哇木棉为0.59 g·m-2·d-1,美丽异木棉和海南木棉均为0.68 g·m-2·d-1,爪哇木棉日固碳量低于美丽异木棉和海南木棉,因此说明爪哇木棉固碳能力低于美丽异木棉、海南木棉;3种木棉年固碳量在214.49~246.11 g之间(见图7),其中爪哇木棉比另两品种碳汇量低,约为12.8%.

表4 3种木棉的固碳量
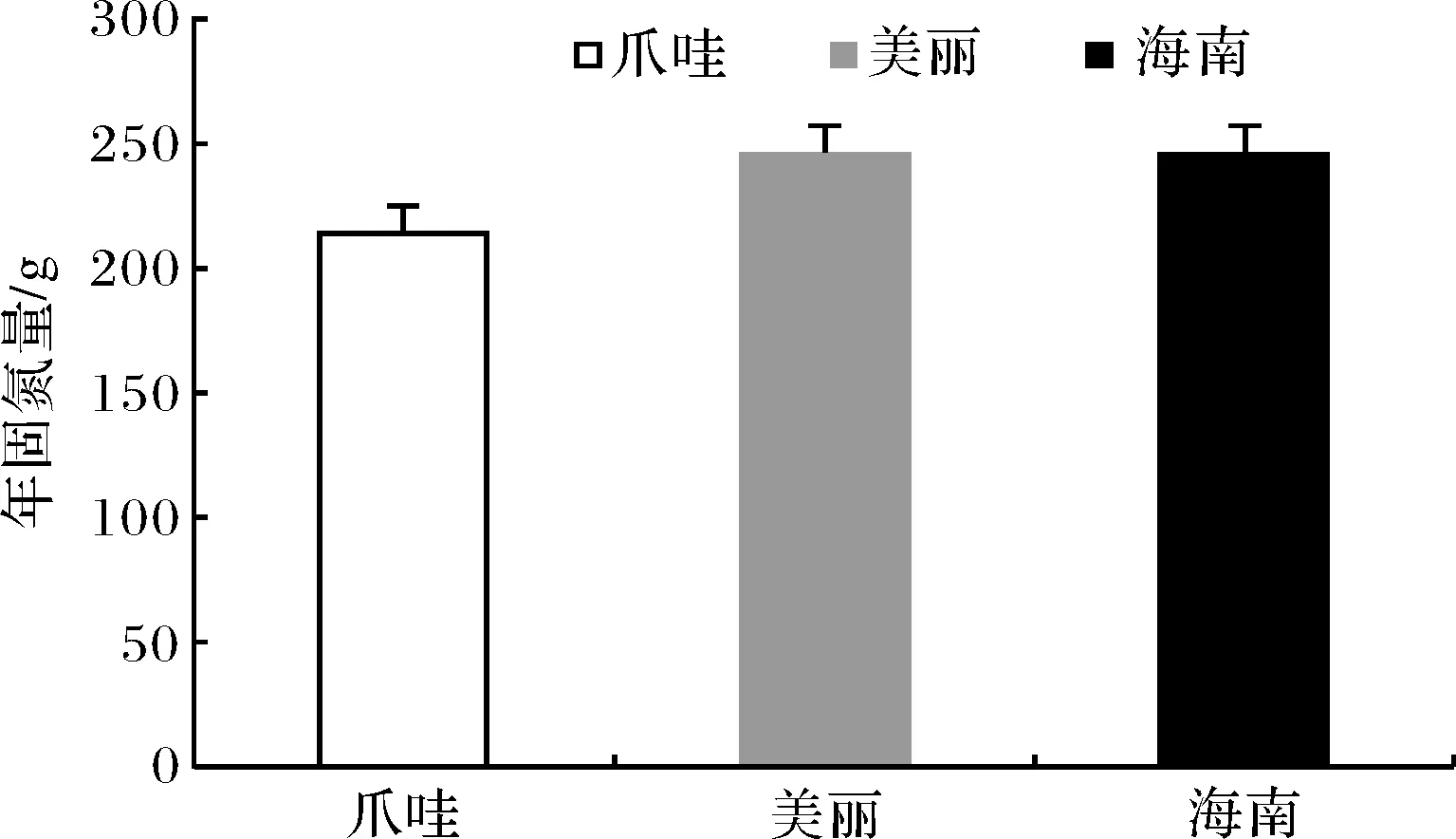
图7 3种木棉的年固碳量Fig.7 Annual carbon sequestration of three kinds of kapok
本实验中3种木棉都为园林绿化良好树种,树型、花形、花色各有特色,受到人们的喜爱,但在种植过程中既要考虑到美化作用又要考虑到该树种对大气环境的影响.特别是近年来,随着我国城市化不断发展,木棉的种植面积、数量也不断增多,而木棉在美化环境净化空气中作用也相应的增大.因此,在种植选择过程中,碳汇能力的大小和对环境美化的作用可作为树种选择的依据,为其提供参考.
采用 SPSS 11.0 软件对3种木棉的碳汇值采用欧式最短距离法进行聚类分析,从图8可知,三品种木棉可分为两个等级,一级为爪哇木棉,二级为美丽异木棉、海南木棉,说明美丽异木棉、海南木棉两个品种的碳汇能力相近.

图8 3种木棉碳汇能力聚类Fig.8 Clustering of carbon sequestration capacity of three kinds of kapok
3结论与讨论
已有研究表明,因物种和环境因素的差异,光合作用净光合速率日变化呈现不同的曲线类型[13-15],有些植物呈单峰曲线,有些呈双峰曲线.本文对木棉科3个重要园林绿化树种(美丽异木棉、爪哇木棉、海南木棉)进行光合速率测定得知,3种木棉Pn日变化规律基本都呈不同程度的“M”型双峰曲线,存在明显的“午休”现象.这与吴吉林等[16]研究结果一致.但植物发生生理性“午休”现象可能由于中午高温低湿,气孔导度变小,CO2含量变低等环境条件所引起植物机体发生的一种保护机制.因此在育苗的过程中,夏天中午尽量对苗圃进行遮阴,降低大气温度,增加空气湿度,在冬季低温时对苗圃采取增温保护措施.
Tr的大小在一定程度上反映植物调节水分及适应逆境能力,与植物光合速率密切相关[17].一般认为Pn变化与Tr呈相同趋势变化.但通过对3种木棉Tr结果分析得知,3种木棉品种Tr日变化均呈单峰曲线变化,在中午12∶00~13∶00时温度为33℃,接近一天中最高温度,空气湿度为60 HR/%,为一天中最低空气湿度,并且在Pn发生“午休”现象时呈上升趋势,较高的大气温度和相对较干燥的空气湿度使蒸腾速率在中午13∶00时出现小高峰.美丽异木棉、爪哇木棉、海南木棉分别为1.66 μmol·m-2·s-1、1.78 μmol·m-2·s-1和1.62 μmol·m-2·s-1,爪哇木棉Tr出现的时间和峰值都早于、高于另两个品种.而Tr的大小在一定程度上反映了植物调节水分损失及适应逆境的能力[13,18-20].由此表明爪哇品种在高温、强光的情况下能更好适应环境,进行光合作用,降低高温对植株的生理性伤害.
3个木棉品种Gs都呈单峰曲线变化,从早上8∶00开始上升,上午10∶00时达到最大,之后呈下降趋势,早上和晚上Gs较小,蒸腾作用最弱,Tr较低,随着温度和光强的增加,Gs和叶内和叶外水蒸气压力也随之增大,蒸腾速率也呈不断上升的趋势,中午前后达到峰值,午后植物为了维持体内水分平衡,较强蒸腾耗水使气孔导度减小,且随着光照减弱,蒸腾速率也随之减弱.气孔导度美丽异木棉>爪哇木棉>海南木棉.Ci与Pn基本呈相反的单峰曲线变化,与马志波研究六种阔叶树种结果相一致[21],其可能与植物白天光合作用利用 CO2量大而引起胞间碳浓度下降有关.
光合速率日变化与生理生态因子相关性分析表明:海南木棉、爪哇木棉Pn与PAR、Tr呈极显著相关,与Ci呈负相关,与高雪等试验结论表达相同[13];爪哇木棉与Ca、RH呈负相关;美丽异木棉Pn与PAR、Tr、Ta呈显著相关,与Ca、Ci呈负相关;3种木棉Pn随PAR、Tr增加而加快,同时随着Ci、Ca、RH增加而减弱,与其他生理生态因子无关,其结果与上述Pn与PAR、Tr等环境因子关系结果相一致.
植物固碳量是通过净日同化量根据净光合速率转换而来[22],而植物生理生态因子变化直接影响着固碳量的多少.3种木棉单位日固碳量在0.59~0.68 g·m-2·d-1之间,其中爪哇木棉为0.59 g·m-2·d-1,美丽异木棉和海南木棉均为0.68 g·m-2·d-1,爪哇木棉日固碳量明显低于美丽异木棉和海南木棉,因此说明爪哇木棉固碳能力明显低于美丽异木棉、海南木棉;3种木棉年固碳量在214.49~246.11 g之间,其中爪哇木棉比另两品种碳汇量低,约为12.8%.3种木棉碳汇能力大小表现为美丽异木棉、海南木棉两品种高于爪哇木棉.聚类分析结果表明:美丽异木棉和海南木棉品种具有极高的相似性,可能在物种进化过程中亲缘关系更近.因此,在园林绿化树种栽植过程中应适当增加美丽异木棉、海南木棉种植面积,可为美化环境净化空气中的CO2做出贡献,为树种的选择提供参考.
植物生长也有特定的生长周期,一般为一年,各个季节气候变化对净光合速率及固碳能力也有一定的影响,以净日光合速率和日固碳量来衡量一个树种的固碳能力不一定完全准确,如果以年固碳量来作为衡量指标会更确切些,更能反映植物动态固碳的能力.但目前有关园林绿化植物年净光合速率方面计算方法还不是太多,还比较难进行准确、全面的估算、评价园林植物年固碳量.因此,在这方面也可以作为研究深入的一个方面,结合物理、数学及植物生理生化等方面的知识、技术,跨领域的解决问题.为更好的探究林园绿化植物碳汇能力,选择种植园林绿化树种提供参考.
[1] HOUGHTON J T, FILHO L M, CALLANDER B A, et al. Climate Change 1995: the Science of Climate Change: Contribution of Working Group I to the Second Assessment Report of the Inter-governmental Panel on Climate Change[M]. Cambridge:Cambridge University Press, 1996.
[2] 刘 宁, 吴左宾. 城市道路绿地景观设计[J].西安建筑科技大学学报, 2005, 35(3):250-253.
[3] 冯建军. 城市道路绿化模式环境效益分析[J].城市环境建设与城市生态, 2006, 200(4):5-8.
[4] 丁铭绩. 浅谈城市道路绿化设计[J].科技情报开发与经济, 2003, 21(3):25-28.
[5] ZELITCH I.The close relationship between net photosynthesis and crop yield[J].BioScience, 1982, 32(10):796-802.
[6] 郭 丽, 刘 坤, 张 栋, 等. 三个红叶石楠品种光合特性的比较研究[J].安徽师范大学学报, 2010, 33(4):371-374.
[7] 袁嘉祖, 范晓明. 中国森林碳汇功能的成本效益分析[J].河北林果研究, 1997, 12(1):20-24.
[8] ISERANDS J C, CEULEMANS R, WIARD B. Genetic variation in photosynthetic traits among populaces clones in relation to yield[J].Plant Physiology Biochem, 1998, 26:427-437.
[9] 许大全. 光合作用效率[M].上海:上海科技出版社, 2000, 125-130.
[10] 张文标, 金则新, 柯世省, 等. 木荷光合特性日变化及其与环境因子相关性分析[J].广西植物, 2006, 26(5):492-498.
[11] AND G D F, SHARKEY T D. Stomatal conductance and photosynthesis[J].Plant Biol, 1982, 33(33):317-345.
[12] 冷平生, 杨晓红. 5种园林树木的光合蒸腾特性的研究[J].北京农学院学报, 2000, 15(4):13-18.
[13] 高 雪, 莫宝盈, 奚如春, 等. 盆栽条件下三个油茶栽培种的光合蒸腾特性比较[J].中南林业科技大学学报, 2012, 32(4):89-94.
[14] 张启昌, 杜凤国, 夏富才, 等. 美国椴光合蒸腾的生理生态[J].华北大学学报(自然科学版), 2000, 5(1):436-438.
[15] 孙 磊, 章 铁, 李宏开, 等. 柿树光合速率日变化及其影响因子的研究[J].北方果树, 2006(2):4-6.
[16] 吴吉林, 李永华, 叶庆生, 等. 美丽异木棉光合特性的研究[J].园艺学报, 2005, 32(6):1061-1064.
[17] 傅瑞树, 叶 青. 油茶某些生理活动的昼夜变化[J].林业科技通讯, 1984(8):11-14.
[18] 吴吉林, 李永华, 叶庆生, 等. 美丽异木棉光合特性的研究[J].园艺学报, 2005, 32(6):1061-1064.
[19] 刘奉觉, 郑世楷, 巨关升, 等. 树木蒸腾耗水测算技术的比较研究[J].林业科学, 1997, 33(5):117-125.
[20] 陈 硕, 王 琪, 任志刚, 等. 城市绿地碳汇能力研究[J].山东林业科技, 2011(6):21-23.
[21] 马志波, 马庆彦, 韩海荣, 等. 北京地区6种落叶阔叶树光合特性的研究[J].北京林业大学学报, 2004, 3(26):13-18.
[22] 冷平生, 杨晓红. 5种园林树木的光合蒸腾特性的研究[J].北京农学院学报, 2000, 15(4):13-18.
The photosynthetic characteristics andcarbon sink capacity of three kinds of Kapok
WU Jinshan, WANG Yachen, XU Shitao, ZHAO Qingjie, LI Yingying, YU Xudong
(College of Tropical Agriculture and Forestry, Hainan University, Danzhou, Hainan 571737)
The blades of three kinds of Kapok, includingChorisiaspeciosa,CeibapentandraandBombaxcelibate, were used as experimental materials. The daily change in photosynthesis were measured by 3051D photosynthetic apparatus with their correlation analyzed. Then the amount of carbon sequestration were calculated by their photosynthetic rate, and thus the carbon sequestration capacity of each was obtained. Results are as follows: the daily change in photosynthetic rate (Pn) of the three kinds of kapok follow an pattern of “M” bimodal curve, having an obvious “noon nap” phenomenon. Each kind of kapok’s transpiration rate (Tr) varies as a single-peak curve with a peak during 12∶00~14∶00 and the Tr ofCeibapentandrapossess earlier apperand higher than the other two varieties; the daily solid carbon of the three Kapoks is 0.59~0.68 g·m-2·d-1,of which that of theChorisiaspeciosaandBombaxcelibateare both 0.68 g·m-2·d-1, while kapok 0.59 g·m-2·d-1. The annual solid carbon of the three kapoks is 214.49~246.11 g, with the carbon sequestration capacity of theChorisiaspeciosaweaker than the other two varieties, about 12.8% lower. Cluster analysis showed that there is an extremely high similarity betweenChorisiaspeciosaandBombaxcelibate. This study provides reference for the breeding of optimized variety of kapok trees in gardening.
Kapok; photosynthetic features; carbon sequestration; measurement
2016-09-03.
海南省自然科学基金项目(314057);海南省产学研一体化专项项目(CXY20140027);海南大学中西部高校提升综合实力工作资金项目(201417);海南大学应用科技学院(儋州校区)基金项目(HYK1524);国家级大学生创新创业训练计划项目(201421).
1000-1190(2017)01-0072-07
Q945.11
A
*通讯联系人. E-mail: doeast@163.com.
杂志排行

华中师范大学学报(自然科学版)的其它文章
- 基于高阶相关聚类的脱机手写文本行分割
- 基于改进ICA算法对云南地区重力固体潮中地震前兆信息的提取与识别
- 2,3-二芳基-1,3-苯并噁嗪的合成及对利什曼原虫CYP51活性的初步研究
- W/Sn/Mg复合氧化物催化合成过氧乙酸的工艺研究
- Fabrication of PZT PCs with nanoporearrays by sol-gel process
- Mechanism exploration of xylitol selenite-inducedapoptosis in a human hepatoma cell lineSMMC-7221 cells using real-time quantitative PCR