养分添加对贝加尔针茅草原6种植物叶片性状的影响
2017-05-23刘红梅李洁于丽皇甫超河杨殿林
刘红梅,李洁,于丽,皇甫超河,杨殿林
(农业部环境保护科研监测所,天津300191)
养分添加对贝加尔针茅草原6种植物叶片性状的影响
刘红梅,李洁,于丽,皇甫超河,杨殿林*
(农业部环境保护科研监测所,天津300191)
养分添加是促进退化草地恢复常用的管理措施。以贝加尔针茅草原为研究对象,设置CK(对照,不施肥),N(单施氮肥,100 kg/hm2),P(单施磷肥,100 kg/hm2),K(单施钾肥,100 kg/hm2),NP(氮、磷肥混施,均为100 kg/hm2),NK(氮、钾肥混施,均为100 kg/hm2),PK(磷、钾肥混施,均为100 kg/hm2),NPK(氮、磷、钾肥混施,均为100 kg/hm2),研究贝加尔针茅草原包括建群种和优势种在内的6种植物的比叶面积、叶绿素含量和养分含量等叶片性状对养分添加的响应,旨在探讨草原生态系统中,不同物种叶片特性随养分供给条件发生的变异及其与土壤理化特性之间的关系。结果表明,6种植物的比叶面积(specific leaf area, SLA)、叶绿素含量、叶片养分在不同养分添加下都发生了变化,但变化的范围和方向都不尽相同。综合6种植物进行分析,SLA与叶绿素含量、叶片含N量呈极显著正相关(P<0.01),与叶片含P量呈显著正相关(P<0.05);叶绿素含量与叶片含N量、土壤铵态氮含量呈极显著正相关(P<0.01);叶片含N量与土壤pH呈显著正相关(P<0.05);叶片含P量与土壤P、土壤速效P含量和土壤pH值呈极显著正相关(P<0.01)。综合研究表明,贝加尔针茅草原植物叶片含N量较低,植物叶片性状受N素影响显著,但不同物种对N素添加的反应不同,土壤养分供给的差异是叶片结构特性和叶片养分组成发生变化的重要原因。
贝加尔针茅草原;养分添加;叶片性状;土壤理化性质
植物功能特性受遗传因素和外界环境共同影响[1],其变化既表征了生态系统基本的演替过程,也反映了群落对不同干扰所做出的综合响应,客观地表达了植物对环境条件的适应机制[2]。叶片是植物获取资源的重要器官,对环境变化敏感且可塑性大[3]。植物叶片的比叶面积、叶绿素含量、叶片含氮量、叶片含磷量等生理生态特征体现物种本身的生物学特性,与植物的生长对策及植物利用资源的能力紧密联系,能够反映植物适应环境变化所形成的生存对策,具有重要的生态学意义[4-5]。近年来,国内外研究者针对各种不同生活型、不同的功能型植物进行了大量的对比研究,获取了大量关于植物叶片性状变异的相关数据[6-10]。但目前,对于养分添加导致群落结构改变的机制没有取得一致的结论。一些研究者认为,物种间养分竞争力的差异是导致优势种发生转换的主要因素[11],另一些研究者则认为,养分添加使物种间的竞争转向光资源,光竞争能力的强弱是决定物种群落结构和物种替代的主要原因[6]。此外,一些研究表明,物种对养分添加的响应不仅与物种本身的功能特性密切相关,还受物种所在群落的环境条件的影响[3,12-13]。有关植物叶特性与土壤的相关研究主要集中在农田和森林生态系统方面[5,14-15],对草原生态系统植物功能特性与土壤养分供应关系的研究较少。当土壤营养状况发生改变后,植物叶片各功能性状组成将会发生怎样的改变?哪些性状的变化程度更大?何种因素起关键作用?这些问题还未得到充分的认知。
贝加尔针茅(Stipabaicalensis)草原是欧亚大陆草原的重要组成部分,是内蒙古草甸草原的代表类型之一,在我国畜牧业生产中占有重要的地位[16]。由于自20世纪80年代以来高强度的利用和不完善的保护机制,贝加尔针茅草原已出现不同程度的退化,总体表现为生物多样性降低,草地生产力下降,草畜矛盾日益加剧,草原的恢复重建迫在眉睫[17]。养分是草地生态系统生产力主要限制因素之一。科学的养分管理是提高草原生态系统功能,恢复退化草地植被的主要措施。氮素是草原生态系统中植物生长的最普遍、最主要的限制因子[18],磷有时也会成为主要限制的因素,其作用仅次于氮[19],钾也是植物生长的限制因素之一。关于氮添加对草原植物性状的影响已有很多研究,但关于P和K添加对草原植物性状的研究还很少。基于此,本试验比较研究了经过连续6年养分添加处理后,贝加尔针茅草原包括建群种和优势种在内的6种植物,在不同养分添加处理下比叶面积(specific leaf area, SLA)、叶绿素含量(leaf chlorophyll content)、叶片含氮量(leaf nitrogen content)、叶片含磷量(leaf phosphorus content)的变化,分析叶性状之间的关系及其与土壤理化性质的关系,以探讨典型草原生态系统中不同植物叶片性状的变异范围及其对养分供应改变的响应机制,为贝加尔针茅草原的植物恢复与重建提供理论依据。
1 材料与方法
1.1 研究区域概况
研究区位于大兴安岭西麓,内蒙古自治区鄂温克自治旗伊敏苏木境内,地理位置为北纬48°27′-48°35′,东经119°35′-119°41′,海拔为760~770 m,地势平坦,属于温带草甸草原区。试验地所在地区年均降水量328.7 mm,年均气温-1.6 ℃,≥0 ℃年积温2567.5 ℃,年均风速4 m/s,无霜期113 d,属半干旱大陆性季风气候,降水主要集中于7-8月。土壤类型为暗栗钙土[20]。植被类型为贝加尔针茅草甸草原,贝加尔针茅为建群种,羊草(Leymuschinensis)为优势种,羽茅(Achnatherumsibiricum)、扁蓿豆(Melissitusruthenica)、草地麻花头(Serratulaamatsutanna)、线叶菊(Filifoliumsibiricum)、日荫菅(Carexpediformis)、变蒿(Artemisiacommutata)、多茎野豌豆(Viciamulticaulis)、寸草苔(Carexduriuscula)、肾叶唐松草(Thalictrumpetaloideum)等为常见种或伴生种。共有植物66种,分属21科49属。
1.2 试验设计
试验地自2010年开始进行围封禁牧,并进行相应的施肥处理。试验设CK(对照,不施肥),N(单施氮肥,100 kg/hm2),P(单施磷肥,100 kg/hm2),K(单施钾肥,100 kg/hm2),NP(氮、磷肥混施,均为100 kg/hm2),NK(氮、钾肥混施,均为100 kg/hm2),PK(磷、钾肥混施,均为100 kg/hm2),NPK(氮、磷、钾肥混施,均为100 kg/hm2)8个处理,6次重复。小区面积8 m×8 m=64 m2,处理之间设2 m隔离带,重复间设5 m隔离带。养分添加试验于2010年开始进行,每年分两次进行添加,分别在牧草生长季 6 月 15 日、7 月 15 日进行,每次施入全年添加总量的50%。N(尿素)、P(重过磷酸钙)、K(硫酸钾)施肥时均匀手撒。
1.3 样品采集与测定
于2015年8月10日开展实验测定和采集样品。此时6种草地常见植物(贝加尔针茅、羊草、羽茅、扁蓄豆、线叶菊和草地麻花头)已经完成叶片形态建成,生育时期为营养生长盛期至初花期。在每个小区,每种物种剪取完整健康植株30株。其中15株用于测定叶面积,另15株低温保存用于测定叶绿素。同时用土壤采样器在各个小区内按照S型取样法选取10个点,去除表面植被,取0~10 cm土壤混匀,去除根系和土壤入侵物,采用“四分法”选取1 kg土壤,迅速装入无菌封口袋,将其分成两部分,一部分于-20 ℃超低温冰箱中保存,用于土壤速效养分分析,一部分土样于室内自然风干后研磨过筛,用于土壤理化性质分析。
比叶面积测定:采用Li-3100A叶面积仪 (Li-Cor, Lincoln, Nebraska, USA) 测定叶面积。将测定完叶面积的叶片单独装在自封袋中,带回实验室烘干至恒重,测定叶片干重。按照以下公式计算比叶面积:
SLA=ULA/DY
(1)
式中:SLA为叶片比叶面积,单位为m2/kg;ULA为单位叶片面积,单位为m2;DY为叶片干重,单位为kg。
叶绿素含量的测定:把采集的6种植物鲜叶用去离子水清洗干净,称取叶片0.30 g,用丙酮法提取色素,采用分光光度计(Unic 2800 UV/Vis Spectrophotometer)测定提取液在663和645 nm处吸光度值,按照以下公式[21]计算:
ρ(Chl a)=12.7OD663-2.69OD645
(2)
ρ(Chl b)=22.9OD645-4.68OD663
(3)
ρ(Chl)=ρ(Chl a)+ρ(Chl b)
(4)
式中:OD663、OD645分别为叶绿体色素提取液在波长663和645 nm下的吸光度值;ρ(Chla)、ρ(Chlb)分别为叶绿素a、叶绿素b的质量浓度,g/kg;ρ(Chl)为叶绿素总量,g/kg。
叶片元素的测定:植物样品105 ℃杀青30 min,然后65 ℃烘干至恒重,叶片粉碎过0.15 mm筛,混匀后保存在塑封袋中以备分析。叶片N、P含量测定采用浓硫酸-双氧水比色法[15]。
土壤理化性质测定[22]:土壤样品自然风干后,去除根、石头等杂物,研磨,分别过0.85和0.15 mm筛并保存。土壤有机质测定采用水合热重铬酸钾-比色法测定,土壤NH4+-N和NO3--N含量采用流动注射分析仪(QC8000,Lachat,USA)测定,土壤全N采用开氏消煮法,土壤全P采用钼锑抗比色法,土壤pH按土水比1∶2.5测定。经过6年连续养分添加后土壤理化性质见表1。
1.4 数据分析
应用SPSS 16.0进行统计分析,应用Excel 2010对分析结果作图。统计数据以平均值和标准误差表示,采用单因素方差分析(one-way ANOVA)和最小显著差数法(LSD法)进行不同养分处理植物均值的方差分析和差异显著性比较(P=0.05)。叶片SLA、叶绿素含量与养分含量、土壤理化性质之间的关系采用Pearson相关分析,并进行双尾显著性检验。
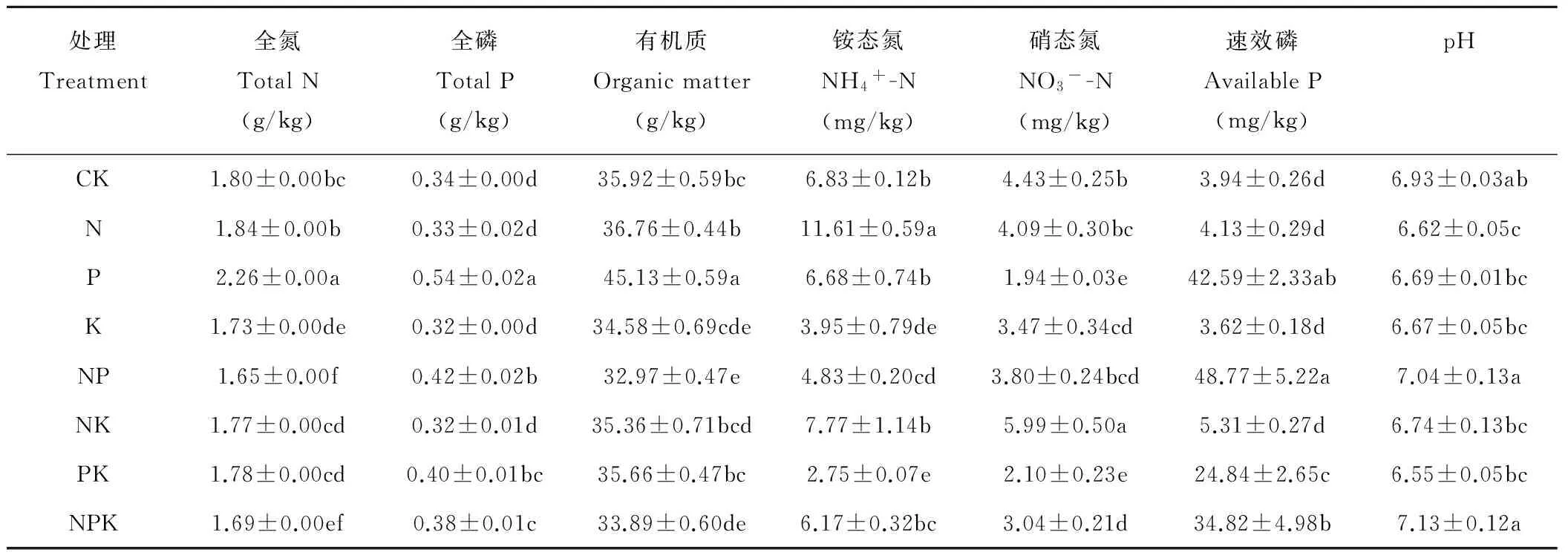
表1 不同养分添加处理下土壤理化性质Table 1 Soil physical and chemical characteristics under different nutrient treatments
注:同列不同字母表示差异显著(P<0.05)。
Note: Different letters in the same column mean significant difference atP<0.05 level.
2 结果与分析
2.1 养分添加对植物比叶面积和叶绿素含量的影响
2.1.1 比叶面积 6种植物在连续6年不同养分添加下,SLA都发生了显著变化(图1),变化幅度从大到小依次为羽茅、扁蓿豆、羊草、草地麻花头、线叶菊、贝加尔针茅。与对照CK相比,含N素添加处理(N、NK、NP、NPK)显著提高了贝加尔针茅、羊草、羽茅、草地麻花头和扁蓿豆的SLA。线叶菊除在NK添加处理SLA与其对照CK无显著差异以外,N、NP、NPK添加处理均显著提高了SLA。含P素添加处理(P、NP、PK、NPK)显著提高了羽茅、扁蓿豆和线叶菊的SLA。贝加尔针茅在P处理添加下,SLA与对照CK相比无显著差异,羊草和麻花头在P处理添加下,SLA显著低于对照CK,在NP、PK、NPK处理下,3种植物的SLA均显著高于对照CK。含K素添加处理下,6种植物的SLA无一致的变化。6种植物,N素处理SLA均高于或显著高于P素处理和K素处理,说明N素添加处理对SLA影响均高于P素添加处理和K素添加处理的影响。
2.1.2 叶绿素含量 连续6年不同养分添加下,6种植物的叶绿素含量均呈现出对氮添加的敏感性,且禾本科牧草高于豆科和杂类草;磷素的添加,亦可增加禾本科牧草叶绿素含量,但激发效应显著小于氮的添加,对于豆科与杂类草而言,磷的添加,降低了其叶绿素含量(图2)。扁蓿豆在NP养分添加下叶绿素含量最高,其余5种植物均在N养分添加下,叶绿素含量最高。6种植物在N养分添加下的叶绿素含量均显著高于P养分和K养分添加下的叶绿素含量。在N养分添加下,贝加尔针茅、羊草、羽茅、扁蓿豆、草地麻花头、线叶菊的叶绿素含量与对照CK相比分别增加了62.88%,139.42%,84.78%,6.01%,62.05%和22.02%。其中羊草增加最多,扁蓿豆叶绿素含量增加最少。在P养分添加下,贝加尔针茅、羊草、羽茅、草地麻花头与对照CK相比分别增加了17.40%,49.52%,17.67%,18.99%;扁蓿豆、线叶菊叶绿素含量与对照CK相比降低了8.61%,39.77%。在K养分添加下,羊草、羽茅、草地麻花头的叶绿素含量与对照CK相比增加了33.69%,20.38%,11.48%;贝加尔针茅、扁蓿豆、线叶菊叶绿素含量与对照CK相比,降低了5.11%,7.28%,28.76%。除扁蓿豆和线叶菊在P养分、K养分处理对叶绿素含量的变化高于N养分处理外,其余4种植物在N养分处理叶绿素含量变化均显著高于P养分和K养分添加处理。
2.2 养分添加对植物叶片养分含量的影响
2.2.1 叶片含氮量 在不同养分添加下,叶片含氮量变化范围各不相同(图3)。6种植物叶片单位质量含氮量变化从大到小依次为线叶菊、羽茅、草地麻花头、羊草、贝加尔针茅、扁蓿豆。在不同养分添加的处理间,线叶菊的叶片含氮量变化最大,达13.96 g/kg,扁蓿豆的叶片含氮量变化最小,为3.94 g/kg。除扁蓿豆以外,其余5种植物在含N素添加的处理(N、NK、NP、NPK)中叶片含N量均高于各自对照CK。贝加尔针茅、羊草、羽茅、草地麻花头和线叶菊在含P添加的处理(P、NP、PK、NPK)中叶片含N量均高于各自对照CK;扁蓿豆在N、P和NPK添加处理下叶片含N量低于对照CK,在K、NP、NK、PK下高于对照CK。除扁蓿豆在K养分添加下,叶片N含量变化高于N养分添加外,其余5种植物在N养分处理中叶片N含量变化均显著高于P养分和K养分添加处理。

图1 养分添加对6种植物比叶面积的影响Fig.1 Effects of nutrient addition on the SLA of six plant species

图2 养分添加对6种植物叶绿素含量的影响Fig.2 Effects of nutrient addition on the leaf chlorophyll content of six plant species

图3 养分添加对6种植物叶片含氮量的影响Fig.3 Effects of nutrient addition on the leaf nitrogen content of six plant species
2.2.2 叶片含磷量 在不同养分添加下,6种植物叶片含磷量变化范围各不相同(图4)。叶片含磷量变化从大到小依次为扁蓿豆、贝加尔针茅、羽茅、羊草、线叶菊、草地麻花头。扁蓿豆的叶片含磷量变化最大,达7.23 g/kg,草地麻花头的叶片含磷量变化最小,为2.54 g/kg。贝加尔针茅、羊草、羽茅和线叶菊在含P素添加处理(P、NP、PK、NPK)叶片含P量均高于对照CK。扁蓿豆在PK添加处理叶片含P量低于对照CK,在NP、PK和NPK添加处理显著高于对照CK。草地麻花头在P、PK添加处理叶片含P量显著低于对照CK,NP添加处理与对照CK无显著差异,NPK添加处理高于对照CK。对于同一种植物叶片含P量与P素添加相关,P素添加处理的6种植物的叶片含磷量高于N素和K素添加处理。在连续6年不同养分添加处理下,6种植物叶片P含量差异很大,这可能与植物系统发育变化有关。
2.3 植物叶性状之间的关系
对所有不同养分添加处理6种植物的SLA与叶片养分含量的相关分析表明,植物叶片SLA与叶绿素含量、叶片含N量呈极显著正相关(图5A,图5B,P<0.01),与叶片含P量呈显著正相关(图5C,P<0.05)。综合看来,较大的SLA具有较高的叶绿素含量和较高的养分含量,叶片含N量和叶片含P量对植物SLA的影响大于叶片N∶P的影响。

图4 养分添加对6种植物叶片含磷量的影响Fig.4 Effects of nutrient addition on the leaf phosphorus content of six plant species

图5 植物比叶面积与叶绿素含量(A)、叶片含N量(B)、叶片含P量(C)、叶片N∶P(D)的相关性Fig.5 Correlations between SLA and leaf chlorophyll content, leaf nitrogen content, leaf phosphorus content, leaf N∶P
养分添加处理6种植物的叶绿素含量与叶片养分含量相关分析表明,叶绿素含量与叶片含N量呈极显著正相关(图6A,P<0.01),与叶片含P量无显著相关性(图6B,P>0.05),随着叶片N∶P增大,叶绿素含量增加,但无显著相关性(图6C)。随着叶片含P量增加,叶片含N量也增加,但无显著相关性(图6D)。

图6 叶绿素含量与叶片含N量(A),含P量(B),N∶P(C)及叶片含N量与含P量(D)的相关性Fig.6 Correlations between leaf chlorophyll content and leaf nitrogen content, leaf phosphorus content, leaf N∶P and correlation between leaf nitrogen content and leaf phosphorus content
2.4 植物叶性状与土壤特性的关系
植物叶性状与土壤特性的相关分析表明,6种植物SLA与土壤有机质呈极显著负相关(P<0.01),与土壤N呈显著负相关(P<0.05)(表2)。叶绿素含量与土壤铵态氮含量呈极显著正相关(P<0.01)。叶片含N量与土壤pH呈显著正相关(P<0.05)。叶片含P量与土壤P、土壤速效P含量和土壤pH值呈极显著正相关(P<0.01),与土壤有机质、土壤硝态氮呈显著负相关(P<0.05),与土壤N∶P呈极显著负相关(P<0.01)。叶片N∶P与土壤硝态氮、土壤N∶P呈极显著正相关(P<0.01),与土壤P含量、速效P含量呈极显著负相关(P<0.01)。
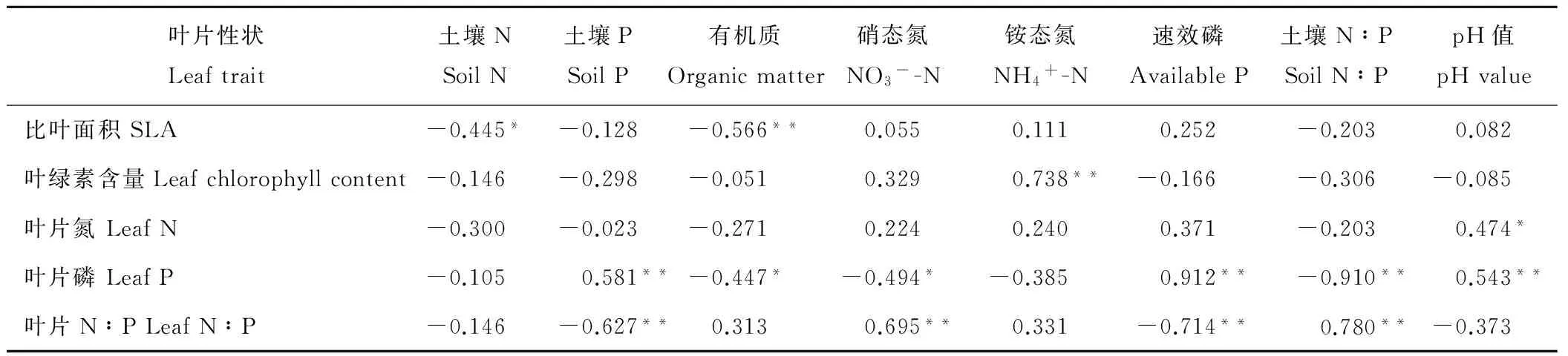
表2 植物叶性状与土壤理化性状之间的关系Table 2 Correlation between leaf traits and soil physical and chemical properties of plants
注:*表示显著相关(P<0.05),**表示极显著相关(P<0.01)。
Note: *indicated significant correlation (P<0.05), **indicated highly significant correlation (P<0.01).
3 讨论
不同植物叶片的化学组分、形态结构和生理活性是相互联系的,这种内在的关系被称为叶片经济谱[23-24]。植物对环境的适应策略可通过各性状之间功能的权衡和协变来实现。当两个或多个重要植物性状在不同物种间具有一致的相关性时,这种性状关系可以认为是形成了一个策略维度。在长期进化的过程中,植物逐渐形成了较强的生理生化调节能力,来适应环境因子的变化[25]。土壤养分供给状况的改变,会显著影响植物的光合作用和矿质代谢等过程。通过养分添加试验研究植物群落结构、功能的变化与植物功能特性的变化之间的联系,有助于反映出植物对环境因子变化的适应性和生态对策[13,26]。
植物叶性状为适应不同环境而发生的改变可以反映它对生境条件变化的敏感程度[27]。N素参与植物体内多种物质的合成代谢,直接间接参与植物的光合作用[28]。随着土壤N有效性提高,可供植物获取的N数量增多,因此植物增大叶片面积,提高叶片的光合能力。P素在细胞膜结构、物质代谢以及植物光合作用中起着极其重要的作用[29-30]。本研究中6种植物的SLA与叶绿素含量、叶片含N量呈极显著正相关(图5A,图5B,P<0.01),与叶片含P量呈显著正相关(图5C,P<0.05),这与于鸿莹等[31]研究结果一致。本研究6种植物叶片的叶绿素含量与植物叶片含氮量极显著正相关(图6A,P<0.01),在6种被观测的植物中,羊草的叶绿素含量增加的最多(图2),说明氮元素的添加使羊草占据更有利的生态位,增强了其在群落中优势地位。羊草作为根茎型禾草,对氮素的响应高于贝加尔针茅、羽茅等丛生型禾草,这与张云海等[32]、万宏伟等[33]研究结果一致。SLA、叶绿素含量、叶片含N量、叶片含P量间的权衡和协变关系反映了植物可通过叶片形态与营养物质的调节分配,适应土壤养分环境变化的策略。上述相关关系证实了植物叶片功能性状间关系的趋同效应。
不同功能群植物在生活史、形态、生理等多个方面均不相同,因此其对生态系统功能的影响也存在差异。本研究所选6个物种分属3个功能群,禾草类的贝加尔针茅、羊草、羽茅,菊科类的草地麻花头、线叶菊,豆科类的扁蓿豆。本研究中6个物种叶片含N量和含P量最高的是豆科的扁蓿豆,叶片含N量最低的是连续K养分添加处理禾草类的羽茅,对于不同的功能群,叶片含N量都表现为豆科大于非豆科。叶片含P量最低的是连续NK养分添加下的禾草类的贝加尔针茅,除此之外,含N和P素添加处理(N、P、NP、NK、PK、NPK)叶片含P量表现为禾草类大于菊科,说明非豆科植物比豆科植物、禾草比非禾草具有更高的养分利用效率,这与于丽等[34]养分添加提升了贝加尔针茅和羊草为主的禾本科地位研究结果互为印证。该研究结果与宾振钧等[35]的研究结果一致。
绿叶养分浓度反映了土壤的养分供应能力[36],叶片摄取养分的多少受土壤养分供应能力的影响。植物叶片的N∶P值能够反映植物在种群、群落、生态系统尺度上生长受养分限制的情况。我国草地植物叶片N浓度的平均值为28.6 g/kg,叶片P平均值为1.9 g/kg[37]。我国草地豆科植物叶片N浓度30.6 g/kg,叶片P浓度的平均值为2.0 g/kg[38]。本研究中6种植物的叶片含N量均低于我国平均值。在8种养分添加处理下,6种植物叶片含P量大致表现为高于我国草地植物叶片 P 浓度的平均值。本研究中6种植物叶片养分对N素和P素添加较为敏感,对K素添加没有一致的结果。除扁蓿豆以外,含N素和P素添加处理显著提高了贝加尔针茅、羊草、羽茅、草地麻花头和线叶菊叶片含N量(图3),贝加尔针茅、羊草、羽茅和线叶菊在含P素添加处理叶片含P量均高于对照CK(图4)。大多数研究认为,豆科植物可以通过生物固能满足自身对N的需求。本研究中N处理降低了扁蓿豆的叶片含氮量,可能是由于N的过量施用抑制了根瘤的形成,降低了生物固氮效率。Koerselman等[39]研究认为,当N∶P<14 时,N 为限制性因子;N∶P>16 时,P 为限制性因子;14 植物叶片性状与其生存环境条件密切相关[40]。叶片特性与土壤理化性质相关分析表明,6种植物叶片SLA、叶绿素含量和叶片养分与土壤理化性质密切相关。本研究中叶片N∶P与土壤N∶P呈极显著正相关(P<0.01),叶绿素含量与铵态氮含量显著正相关,这与顾大形等[15]研究结果一致。叶片N∶P与土壤全N含量显著负相关,这与韦兰英等[3]研究结果一致。在养分添加处理中,6种植物的叶片性状因子SLA、叶绿素含量、叶片养分含量均发生了显著变化,但变化的范围和方向都不尽相同,说明贝加尔针茅草原共存的6种植物其叶片性状特征对环境异质性具有不同的响应。6种植物叶片性状的变异机制以及同一养分添加处理下不同植物叶片性状的差异原因还需进一步深入研究。 综合以上分析,养分添加改变了6种植物的叶片功能特性,N素添加处理对于叶片SLA和叶绿素含量的影响高于P素和K素添加处理。未添加养分处理,6个物种N∶P在3.11~9.01之间,与我国其他地区草地植物N∶P相比较,植物叶片N∶P都比较低,说明贝加尔针茅草原群落主要受N素限制。叶片性状与土壤理化性质密切相关,说明土壤养分供给条件是叶片结构特性和叶片养分组成发生变化的重要原因。该研究结果对退化草地生态系统的恢复与管理具有重要的指导意义。在采用养分添加促进退化草地恢复的实践中,可以考虑通过养分添加的方法改善土壤条件,并应考虑草地生态系统自身原有的养分状况,设置适当的N、P肥施用量。对于植物生长受氮素限制的草地,施用氮肥是提高草地生产力的首选措施;对于氮素不是限制因素的草地,适当添加P肥有助于生产力的提高。 References: [1] Donovan L A, Maherali H, Caruso C M,etal. The evolution of the worldwide leaf economics spectrum. Trends in Ecology and Evolution, 2011, 26(2): 88-95. [2] Mclntyre S, Lavorel S, Landsberg J,etal. Disturbance response in vegetation-towards a global prespective on functional traits. Journal of Vegetation Science, 1999, 10(5): 621-630. [3] Wei L Y, Shangguan Z P. Relation between specific leaf areas and leaf nutrient contents of plants growing on slope lands with different farming-abandoned periods in the Loess Plateau. Acta Ecologica Sinica, 2008, 28(6): 2526-2535. 韦兰英, 上官周平. 黄土高原不同退耕年限坡地植物比叶面积与养分含量的关系. 生态学报, 2008, 28(6): 2526-2535. [4] Casadebaig P, Debaeke P, Lecoeur J. Thresholds for leaf expansion and transpiration response to soil water deficit in arrange of sunflower genotypes. European Journal of Agronomy, 2008, 28(4): 646-654. [5] Wei L Y, Huang Y Q, Li X K,etal. Effect soil water on photosynthetic characteristics and leaf traits ofCyclobalanopsisglaucaseedlings growing under nutrient-rich and-poor soil. Acta Ecologica Sinica, 2009, 29(3): 160-165. [6] Knops J M H, Reinhart K. Specific leaf area along a nitrogen fertilization gradient. The American Midland Naturalist, 2000, 144(2): 265-272. [7] Zheng Y, Wen Z M, Song G,etal. Adaptation strategies of different plant functional types and their composition along a vegetation restoration gradient in a forest-steppe zone in the Yanhe River catchment, Shaanxi, China. Acta Ecologica Sinica, 2015, 35(7): 5834-5845. 郑颖, 温仲明, 宋光, 等.延河流域森林草原区不同植物功能型适应策略及功能型物种数量随退耕年限的变化. 生态学报, 2015, 35(17): 5834-5845. [8] Diaz S, Lavorel S, McIntyre S U E,etal. Plant trait responses to grazing: a global synthesis. Global Change Biology, 2007, 13: 313-341. [9] Meng T T, Ni J, Harrison S P. Plant morphometric traits and climate gradients in northern China: a meta-analysis using quadrat and flora data. Annals of Botany, 2009, 104(6): 1217-1229. [10] Swenson N G, Enquist B J, Pither J,etal. The biogeography and filtering of woody plant functional diversity in North and South America. Global Ecology and Biogeography, 2012, 21(8): 798-808. [11] Aerts R. Interspecific competition in natural plant communities: mechanisms, trade-offs and plant-soil feedbacks. Journal of Experimental Botany, 1999, 50: 29-37. [12] Knops J M, Reinhart K. Specific leaf area along a nitrogen fertilization gradient. American Midland Naturalist, 2000, 144(2): 265-272. [13] Pennings S C, Clark C M, Cleland E E,etal. Do individual plant species show predictable responses to nitrogen addition across multiple experiments. Oikos, 2005, 110(3): 547-555. [14] McIntyre S. The role of plant leaf attributes in linking land use to ecosystem function in temperate grassy vegetation. Agriculture, Ecosystems&Environment, 2008, 128(4): 251-258. [15] Gu D X, Chen S L, Huang Y Q. Effects of soil nitrogen and phosphonium on leaf nitrogen and phosphonium stoichiometric characteristics and chlorophyll content ofOligostachyumlubricum. Chinese Journal of Plant Ecology, 2011, 35(12): 1219-1225. 顾大形, 陈双林, 黄玉清.土壤氮磷对四季竹叶片氮磷化学计量特征和叶绿素含量的影响. 植物生态学报, 2011, 35(12): 1219-1225. [16] Zhao S, Zhang J N, Lai X,etal. Analysis of microbial biomass C, N and soil microbial community structure ofStipasteppes using PLFA at grazing and fenced in Inner Mongolia, China. Journal of Agro-Environment Science, 2011, 30(6): 1126-1134. 赵帅, 张静妮, 赖欣, 等. 放牧与围栏内蒙古针茅草原土壤微生物生物量碳、氮变化及微生物群落结构PLFA分析. 农业环境科学学报, 2011, 30(6):1126-1134. [17] Zhao X Z, Liu H M, Huangfu C H,etal. Responses of functional diversity of soil microbial communities to nutrient additions inStipabaicalensissteppe in Inner Mongolia, China. Journal of Agro-environment Science, 2014, 33(10): 1933-1939. 赵晓琛, 刘红梅, 皇甫超河, 等. 贝加尔针茅草原土壤微生物功能多样性对养分添加的响应. 农业环境科学学报, 2014, 33(10): 1933-1939. [18] LeBauer D S, Treseder K K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology, 2008, 89(2): 371-379. [19] Vance C P, Uhde-Stone C, Allan D L. Phosphorus acquisition and use: critical adaptations by plants for securing a nonrenewable resource. New Phytologist, 2003, 157(3): 423-447. [20] Zhang N L, Wan S Q, Li L H. Impacts of urea N addition on soil microbial community in a semiarid temperate steppe in northern China. Plant and Soil, 2008, 311(1/2): 19-28. [21] Shu Z, Zhang X S, Chen J,etal. The simplification of chlorophyll content measurement. Plant Physiology Communications, 2010, 46(4): 399-402. 舒展, 张晓素, 陈娟, 等.叶绿素含量测定的简化. 植物生理学通讯, 2010, 46(4): 399-402. [22] Bao S D. Soil and Agricultural Chemistry Analysis[M]. Third Edition. Beijing: China Agricultural Press, 2000. 鲍士旦. 土壤农化分析[M]. 第三版. 北京: 中国农业出版社, 2000. [23] Osnas J L D, Lichstein J W, Reich P B,etal. Global leaf trait relationships: mass, area, and the leaf economics spectrum. Science, 2013, 340: 741-744. [24] Read Q D, Moorhead L C, Swenson N G,etal. Convergent effects of elevation on functional leaf traits within and among species. Functional Ecology, 2014, 28(1): 37-45. [25] Weng E S, Zhou G S. Defining plant functional types in China for global change studies. Journal of Plant Ecology, 2005, 29(1): 81-97. 翁恩生, 周广胜. 用于全球变化研究的中国植物功能型划分. 植物生态学报, 2005, 29(1): 81-97. [26] Brewer J S. Nitrogen addition does not reduce belowground competition in a salt marsh clonal plant community in Mississippi (USA). Plant Ecology, 2003, 168(1): 93-106. [27] Li Y L, Cui J Y, Su Y Z. Specific leaf area and leaf dry matter content of some plants in different dune habitats. Acta Ecologica Sinica, 2005, 25(2): 304-311. 李玉霖, 崔建垣, 苏永中. 不同沙丘生境主要植物比叶面积和叶干物质含量的比较. 生态学报, 2005, 25(2): 304-311. [28] Chown S L, Labber S, Mcgeoch M A,etal. Phenotypic plasticity mediates climate change responses among invasive and indigenous arthropods. Proceedings of the Royal Society B: Biological Sciences, 2007, 274: 2531-2537. [29] Feng Y L, Cao K F, Feng Z L,etal. Acclimation of lamina mass per unit area, photosynthetic characteristics and dark respiration to growth light regimes in four tropical rainforest species. Acta Ecologica Sinica, 2002, 22(6): 901-910. 冯玉龙, 曹坤芳, 冯志立, 等.四种热带雨林树种幼苗比叶重,光合特性和暗呼吸对生长光环境的适应. 生态学报, 2002, 22(6): 901-910. [30] Wright I J, Reich P B, Westoby M. Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats. British Ecological Society, 2001, 15(4): 423-434. [31] Yu H Y, Chen Y T, Xu Z Z,etal. Analysis of relationships among leaf functional traits and economics spectrum of plant species in the desert steppe of Nei Mongol. Chinese Journal of Plant Ecology, 2014, 38(10): 1029-1040. 于鸿莹, 陈莹婷, 许振柱, 等. 内蒙古荒漠草原植物叶片功能性状关系及其经济谱分析.植物生态学报, 2014, 38 (10): 1029-1040. [32] Zhang Y H, He N P, Zhang G M,etal. Nitrogen deposition andLeymuschinensisleaf chlorophyll content in Inner Mongolian grassland. Acta Ecologica Sinica, 2013, 33(21): 6786-6794. 张云海, 何念鹏, 张光明, 等. 氮沉降强度和频率对羊草叶绿素含量的影响. 生态学报, 2013, 33(21): 6786-6794. [33] Wan H W, Yang Y, Bai S Q,etal. Variations in leaf function traits of six species along a nitrogen addition gradient inLeymuschinensissteppe in Inner Mongolia. Journal of Plant Ecology, 2008, 32(3): 611-621. 万宏伟, 杨阳, 白世勤, 等. 羊草草原群落6种植物叶片功能特性对氮素添加的响应. 植物生态学报, 2008, 32(3): 611-621. [34] Yu L, Zhao J N, Wang H,etal. Effects of nutrient addition on plant diversity and productivity in aStipabaicalensisgrassland in Inner Mongolia, China. Acta Ecologica Sinica, 2015, 35(24): 8165-8173. 于丽, 赵建宁, 王慧, 等. 养分添加对内蒙古贝加尔针茅草原植物多样性与生产力的影响. 生态学报, 2015, 35(24): 8165-8173. [35] Bin Z J, Wang J J, Zhang W P,etal. Effects of N addition on ecological stoichiometric characteristics in six dominant plant species of alpine meadow on the Qinghai-Xizang Plateau, China. Chinese Journal of Plant Ecology, 2014, 38(3): 231-237. 宾振钧, 王静静, 张文鹏, 等. 氮肥添加对青藏高原高寒草甸6个群落优势种生态化学计量学特征的影响. 植物生态学报, 2014, 38(3): 231-237. [36] Aerts R, Chapin III F S. The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns. Advances in Ecological Research, 1999, 30(8): 1-67. [37] Han W X, Fang J Y, Guo D L,etal. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist, 2005, 168(2): 377-385. [38] He J S, Wang L, Flynn,etal. Leaf nitrogen: phosphorus stoichiometry across Chinese grassland biomes. Oecologia, 2008, 155(2):301-310. [39] Koerselman W, Meuleman A F M. The vegetation N∶P ratio: a new tool to detect the nature of nutrient limitation. Journal of Applied Ecology, 1996, 33(6): 1441-1450. [40] Tsialtas J T, Pritsa T S, Veresoglou D S. Leaf physiological traits and their importance for species success in a Mediterranean grassland. Photosynthetica, 2004, 42(3): 371-376. Effects of nutrient addition on leaf traits of six plant species in aStipabaicalensisgrassland in Inner Mongolia, China LIU Hong-Mei, LI Jie, YU Li, HUANGFU Chao-He, YANG Dian-Lin* Agro-EnvironmentalProtectionInstitute,MinistryofAgriculture,Tianjin300191,China Nutrient addition is an effective management measure for recovery of degraded grassland.Stipabaicalensisgrassland is a type of meadow steppe that plays an important role in livestock husbandry in China. We conducted a field experiment to explore the mechanisms underlying variation in plant leaf traits in a mature typical steppe ecosystem in Inner Mongolia where nutrients have been added since 2010. The experiment had a randomized block design with six blocks of eight treatments: CK (control, no nutrient addition), N (N addition only, 100 kg/ha), P (P addition only, 100 kg/ha), K (K addition only, 100 kg/ha), NP (mixed N and P addition, 100 kg/ha each), NK (mixed N and K addition, 100 kg/ha each), PK (mixed P and K addition, 100 kg/ha each), and NPK (mixed N, P, and K addition, 100 kg/ha each). We selected six dominant and sub-dominant plant species for analyses:S.baicalensis,Leymuschinensis,Achnatherumsibiricum,Melissitusruthenica,Serratulayamatsutanna, andFilifoliumsibicum. Together, these species account for >90% of the total community aboveground biomass. We investigated variations in specific leaf area (SLA), leaf chlorophyll content, leaf N content, and leaf P content of the six plants under different nutrient treatments, to determine their responses to variations in soil physical and chemical properties. In all six plant species, the SLA, chlorophyll content, and leaf nutrient contents varied among different nutrient treatments, but the scope and direction of changes differed among plant species. A correlation analysis demonstrated that leaf SLA was positively correlated with leaf chlorophyll content and leaf N content (P<0.01), and with leaf P content (P<0.05). Leaf chlorophyll content was positively correlated with leaf N content and soil NH4+-N (P<0.01). Leaf N content was positively correlated with soil pH (P<0.01). Leaf P content was positively correlated with soil P, soil available P, and soil pH (P<0.01). Our results indicate that there is a very low leaf N content, and that plant growth is N-limited in theS.baicalensisgrassland in Inner Mongolia, China. Different species had different responses to N addition. Differences in soil nutrients may be one important reason for the observed variations in leaf structure and leaf nutrient contents. Stipabaicalensisgrassland; nutrient addition; leaf trait; soil physic-chemical properties 10.11686/cyxb2016242 http://cyxb.lzu.edu.cn 2016-06-13;改回日期:2016-08-05 国家自然科学基金项目(31170435)资助。 刘红梅(1976-),女,河北沧州人,副研究员。E-mail:liuhongmei@caas.cn*通信作者Corresponding author. E-mail:yangdianlin@caas.cn 刘红梅, 李洁, 于丽, 皇甫超河, 杨殿林. 养分添加对贝加尔针茅草原6种植物叶片性状的影响. 草业学报, 2017, 26(5): 81-91. LIU Hong-Mei, LI Jie, YU Li, HUANGFU Chao-He, YANG Dian-Lin. Effects of nutrient addition on leaf traits of six plant species in aStipabaicalensisgrassland in Inner Mongolia, China. Acta Prataculturae Sinica, 2017, 26(5): 81-91.
