O型口蹄疫病毒不同宿主适应毒P1基因分析
2017-05-18姚怀兵刘梦丽
姚怀兵,赵 毅,朱 妍,刘梦丽,刘 宏,任 方,黄 炯
(1.新疆农业大学动物医学学院,新疆乌鲁木齐 830052;2.天康生物股份有限公司,新疆乌鲁木齐 830000;3.新疆畜牧科学院兽医研究所,新疆乌鲁木齐 830000)
O型口蹄疫病毒不同宿主适应毒P1基因分析
姚怀兵1,3,赵 毅2,朱 妍1,刘梦丽1,刘 宏2,任 方2,黄 炯3*
(1.新疆农业大学动物医学学院,新疆乌鲁木齐 830052;2.天康生物股份有限公司,新疆乌鲁木齐 830000;3.新疆畜牧科学院兽医研究所,新疆乌鲁木齐 830000)
为了明确O型口蹄疫病毒不同宿主适应毒P1基因变异趋势,将口蹄疫病毒分别接种于牛、乳鼠、猪及BHK-21细胞,获得相应宿主的适应毒,以各宿主适应毒RNA为模板,扩增出P1(1A、1B、1C、1D)基因,将各适应毒株扩增的P1基因序列相互比对。结果表明,牛、乳鼠、猪、BHK-21细胞宿主适应毒株P1基因的核苷酸及其编码的氨基酸序列均未发生缺失;部分关键性氨基酸发生了变异,变异发生在VP1主要抗原区域的141-160位和200-213位、VP2抗原表位的156-166位和217-220位;VP3抗原蛋白发生较少氨基酸变异;VP4均未发生变异。试验结果为进一步分析不同宿主适应毒基因组的变异关系奠定了基础。
O型口蹄疫病毒;不同宿主适应毒;P1基因
口蹄疫病毒(Foot-and-mouth disease virus,FMDV)属小RNA病毒科(Picornaviridae)口蹄疫病毒属,是无囊膜病毒,其基因组为单股正链RNA,全长约有8 500 bp左右[1-2]。基因组的中部是一个大的开放阅读框(ORF),编码病毒的多聚蛋白,该聚蛋白经一级裂解后形成L前导蛋白、P1区结构蛋白、P2区和P3区非结构蛋白。FMDV P1区结构蛋白最终成熟裂解为4种结构蛋白VP1、VP2、VP3和VP4,这4种结构蛋白构成了FMDV衣壳[3-4]。其中,结构蛋白VP1、VP2、VP3是构成病毒衣壳的主要蛋白,也是主要抗原蛋白[5-6]。
已有研究表明,在FMDV抗原结构中VP1~VP4都有其重要作用。衣壳蛋白中VPl能诱导动物产生中和抗体,VP2具有抗原性,VP3具有无规则卷曲的柔性,在物理形态上表现出可动性和生化上的多功能性,在很大程度上增加了FMDV的抗原性,其还具有多段抗原决定簇,是机体免疫攻击的对象,在影响病毒衣壳组装和分解中起重要作用[7-11]。VP4蛋白在不同血清型中高度保守,具有1个主要的T细胞表位,在免疫应答中起决定性作用[12-13]。由此可见,P1的研究对掌握FMDV生物学特性和增殖机理,开展免疫学研究,诊断技术研究,新型疫苗开发都有着重要作用。
本研究通过缅甸98谱系FMDV在牛、乳鼠、猪、BHK-21细胞4种宿主进行攻毒、传代,获得以上宿主适应毒,运用分子生物学技术和生物分析软件,分析不同宿主适应毒P1基因内各结构蛋白变异情况,研究FMDV宿主间遗传变异趋势,以期对FMDV基因组的结构特征以及口蹄疫在宿主间传染关系有新的认识。
1 材料与方法
1.1 材料
1.1.1 主要试剂 MiniBEST viral RNA/DNA Extration Kit Ver 5、Primer ScriptTMRT Reagent kit(Perfect Real Time)、DNA Ligation Kit Ver 2.1、T-Vector pMD19 购自TakaRa公司;PCR Master MixTaq酶购自Thermo公司;DNA纯化回收试剂盒、DH5α感受态细胞购自TIANGEN公司;Gibco培养基、100 mL/L胎牛血清购自宝生物工程(大连)有限公司;口蹄疫O型抗体液相阻断ELLSA检测试剂盒购自中国农业科学院兰州兽医研究所。
1.1.2 毒株与细胞 O型口蹄疫缅甸98谱系病毒(O/XJ/10-11)由兰州兽医研究所国家参考实验室分离鉴定,由天康生物股份有限公司保存,作为原始毒种;不同宿主适应毒、幼仓鼠肾细胞(BHK-21)均由天康生物股份有限公司提供和保存。
1.1.3 试验用动物 3日龄~7日龄Balb/c乳鼠购自新疆医科大学;2月龄犊牛、40日龄仔猪由天康生物股份有限公司动物房饲养。
1.2 方法
1.2.1 引物的设计与合成 利用生物学软件Vector NTI 11.5,对O/XJ/10-11毒株的基因序列按P1(1A、1B、1C、1D)基因的顺序分段设计引物(表1),委托北京博迈德科技发展有限公司合成。

表1 引物序列信息
1.2.2 获取不同宿主适应毒 参照《口蹄疫检疫技术规范》(SN/T 1181—2010),将O/XJ/10-11株病毒离心后吸取上清,攻毒乳鼠,收集运动不灵活、四肢麻痹、呼吸迫促、30 h左右出现典型口蹄疫症状最终死亡的乳鼠,处理尸体,采样研磨,4 ℃过夜,离心取其上清,保存备用。牛、猪攻毒前采血,通过ELLSA试剂盒检测为阴性,挑取健康牛、猪、3日龄~7日龄乳鼠,O型口蹄疫病毒抗体进行攻毒,收取足量的不同宿主适应毒。选择细胞形态正常,形成良好单层的BHK-21细胞,倾去旧的Gibco培养基,用不含血清的培养基洗细胞面两次,然后接种第1代乳鼠毒,传代收取BHK-21适应毒。猪毒传至第5代,牛毒传至第4代,乳鼠传至第5代,BHK-21传至第25代,收毒,置-80℃冰箱保存。
1.2.3 病毒RNA的提取 取牛、猪、乳鼠、BHK-21细胞适应毒各200 μL原液,按照RNA抽提试剂盒提供的方法提取RNA,所有器皿和试剂均经DEPC(焦碳酸二乙酯)去RNA酶处理。
1.2.4 cDNA的制备 按照TaKaRa Primer ScriptTMRT Reagent kit反转录试剂盒提供的方法进行RT-PCR。反转录50 μL体系如下:5×buffer prime script 10 μL,Prime Script®RT Enzyme Mix 2.5 μL,Oligo dT Primer 2.5 μL,Random 6 mers 2.5 μL,模板为牛、猪、乳鼠、BHK-21细胞适应毒的RNA各32.5 μL;反转录条件:离心混匀后,在42 ℃水浴40 min~60 min,70 ℃水浴10 min。
1.2.5 阳性DNA克隆、连接和转化 用制备的cDNA为模板进行PCR扩增,其50 μL反应体系如下:TaqDNA聚合酶(0.05 U/μLTaqDNA polymerase,reaction buffer,4 mmol/L MgCl2,0.4 mmol/L dNTPs 25 μL,模板5 μL,上游引物(10 pmol/L)2.5 μL,下游引物(10 pmol/L)2.5 μL,无核酸酶纯水15 μL。离心混匀后置于PCR自动扩增仪中,扩增条件为:94 ℃ 2 min;95 ℃30 s,60 ℃30 s,72 ℃ 1 min 10 s,30次循环;72 ℃延伸10 min。反应完毕后,经过10 g/L的琼脂糖电泳检测。将扩增出的目的片段用DNA胶纯化回收试剂盒纯化,将回收的目的DNA连接到pMD19-T载体,连接体系为:纯化回收的目的基因4.5 μL,pMD19-T载体0.5 μL,SolutionI 5 μL,混匀后放入连接仪中,连接2 h~3 h。连接反应结束后,将连接产物转化到大肠埃希菌DH5α感受态细胞(冰上操作)。37℃、200 r/min摇床孵育,预培养45 min。均匀涂板于含氨苄西林的LB平板,37 ℃过夜培养,挑取单克隆菌落接种于含有氨苄的LB培养液中培养。
1.2.6 目的基因的核苷酸序列的测定及分析 经PCR鉴定为阳性菌后,将阳性菌液保存于150 mL/L的甘油中,送至北京博迈德科技发展有限公司测序。用通用引物对P1基因阳性质粒测序,得到克隆基因片段的核苷酸序列,用Vector NTI 11.5软件进行拼接、翻译,得到4种宿主适应毒的P1基因序列。用DNA Star软件中的Megalign对所得序列与原始序列进行比对分析。
2 结果
2.1 动物攻毒试验结果
将制备的第1代原始毒接种乳鼠,待其发病后采集病料,连续传至第5代。从第4代起,发病时间稳定,毒力变强且比较稳定,每0.2 mL病毒含量LD50在108.0~108.5之间。各代次对乳鼠的毒力及LD50测定结果见表2。将制备的第1 代原始毒接种牛,待其发病后采集病料,继续对牛传代,传至第4 代。测定第1 代至第4 代毒对牛的ID50,结果说明从第 3代起,对牛的毒力变强且比较稳定,毒价ID50在106.1/0.2 mL~106.3/0.2 mL之间(表3)。
将制备的第1 代原始毒接种猪,待其发病后采集病料,继续对猪传代,传至第5 代。测定第1 代至第5 代毒对猪的ID50,结果说明从第 3代起,对猪的毒力变强且比较稳定,毒价ID50在107.1/0.2 mL~107.2/0.2 mL之间(表4)。
2.2 RT-PCR产物的电泳结果
从牛、猪、乳鼠及BHK-21细胞毒的传代性FMDV中提取RNA,并用设计引物对基因1A(VP4)、1B(VP2)、1C(VP3)、1D(VP1)进行PT-PCR扩增,经电泳检测,牛、猪、乳鼠及BHK-21细胞的适应毒分别在399、683、780、672 bp左右均出现扩增条带(图1),均与预期目的片段大小相符,且特异性好。

表2 乳鼠的适应性传代及毒力测试
表3 牛的适应性传代及半数感染量(ID50)的测定
表4 猪的适应性传代及半数感染量(ID50)的测定
1.阴性对照;2~5.BHK-21细胞毒, 2.1D,672 bp;3.1C,780 bp;4.1B,683 bp;5.1A 339 bp;M.DNA 标准 DL 2 000;6~9.猪毒, 6.1D,672 bp;7.1C,780 bp;8.1B,683 bp;9.1A,339 bp;10~13.乳鼠毒,10.1D,672 bp,11.1C,780 bp,12.1B,683 bp,13.1A 339 bp;14~17.牛毒,14.1D,672 bp,15.1C,780 bp,16.1B,683 bp,17.1A,339 bp
2.3 菌液的PCR鉴定结果
连接转化后获得的白色菌落,挑取单菌落进行培养,用1A(VP4)、1B(VP2)、1C(VP3)、1D(VP1)特异性的引物分别对挑取的菌液进行PCR扩增,经电泳检测,牛、猪、乳鼠及BHK-21细胞适应毒的菌液均获得与预期条带大小为399 bp、683 bp、780 bp、672 bp左右的片段(图2)。不同适应毒的1A(VP4)、1B(VP2)、1C(VP3)、1D(VP1)基因已插入pMD19-T载体。
2.4 不同宿主间的P1基因的核苷酸序列及推导的氨基酸序列分析
对不同宿主分别连续传代后,获得不同宿主适应毒,通过与O/XJ/10-11原始毒比较,P1基因由2 208个核苷酸组成,编码736个氨基酸(图3),P1基因核苷酸序列同源性在99.6%~99.8%(图4),氨基酸序列同源性在99.3%~99.9%,核苷酸同源性高,均未出现碱基缺失现象(图5)。核苷酸序列分析有18处变异,其中9处突变发生在密码子第3位碱基上,为同义突变,不会造成所编码的氨基酸突变;第2位碱基上的有5处;第1位碱基上的有4处。推导出的氨基酸序列比对中发生变异的有8处,且均由核苷酸第1、2位碱基突变而造成,为非同义突变。可见,氨基酸变异区明显少于核苷酸变异区,表明病毒表型变化程度低于遗传性变异程度。1A(VP4)由255个核苷酸组成,编码1-85位氨基酸,均未发生变异;1B(VP2)由654个核苷酸组成,编码86-303位氨基酸,其中VP2的70-80位和131-134位氨基酸残基为抗原表位,发生3处变异且均在抗原表位上;1C(VP3)由660个核苷酸,编码304-523位氨基酸,有2处变异;1D(VP1)由639个核苷酸组成,编码524-736位氨基酸,有3处变异。VP1蛋白上氨基酸变异位点有3处,乳鼠适应毒1处发生变异,变异点位于核苷酸序列第472位,由C变为T,氨基酸序列第158位由脯氨酸(P)变为丝氨酸(S),变异发生在FMDV主要抗原区域141-213位氨基酸区域上;猪适应毒2处变异,变异点位于核苷酸序列第356位,由A变为G,氨基酸序列第119位,由酪氨酸(Y)变为半胱氨酸(C),核苷酸序列第532位,由T变为C,氨基酸第178位,由酪氨酸(Y)变为组氨酸(H);牛适应毒和BHK-21细胞适应毒在VP1蛋白上均没有变异。VP2蛋白上氨基酸变异位点有3处,牛适应毒2处变异,变异位点位于核苷酸序列第235位,由T变为C,氨基酸序列第79位,由酪氨酸(Y)变为组氨酸(H),核苷酸序列第544位,由A变为G,氨基酸序列第182位,由甲硫氨酸(M)变为缬氨酸(V);猪适应毒1处变异,变异位点位于核苷酸序列第398位,由A变为G,氨基酸第133位,由谷氨酸(E)变为甘氨酸(G);乳鼠适应毒和BHK-21细胞适应毒在VP2上没有变异。VP3蛋白上氨基酸变异位点有2处,均为BHK-21细胞适应毒变异,变异位点在核苷酸序列第167位,由A变为G,氨基酸序列第56位,由组氨酸(H)变为精氨酸(R),核苷酸序列第179位,由A变为G,氨基酸序列第60位,由天冬氨酸(D)变为甘氨酸(G);VP4基因的氨基酸位点上各宿主适应毒的均没有发生变异,最为最保守。
M.DNA 标准 DL 2 000;1~4.1A 339 bp,1.牛毒,2.猪毒,3.鼠毒,4.BHK-21细胞毒;5~8.1B 683 bp,5.牛毒,6.猪毒,7.鼠毒,8.BHK-21细胞毒;9~12.1C 780 bp,9.牛毒,10.猪毒,11.鼠毒,12.BHK-21细胞毒;13~16.1D 780 bp,13.牛毒,14.猪毒,15.鼠毒,16.BHK-21细胞毒; 17.阴性对照
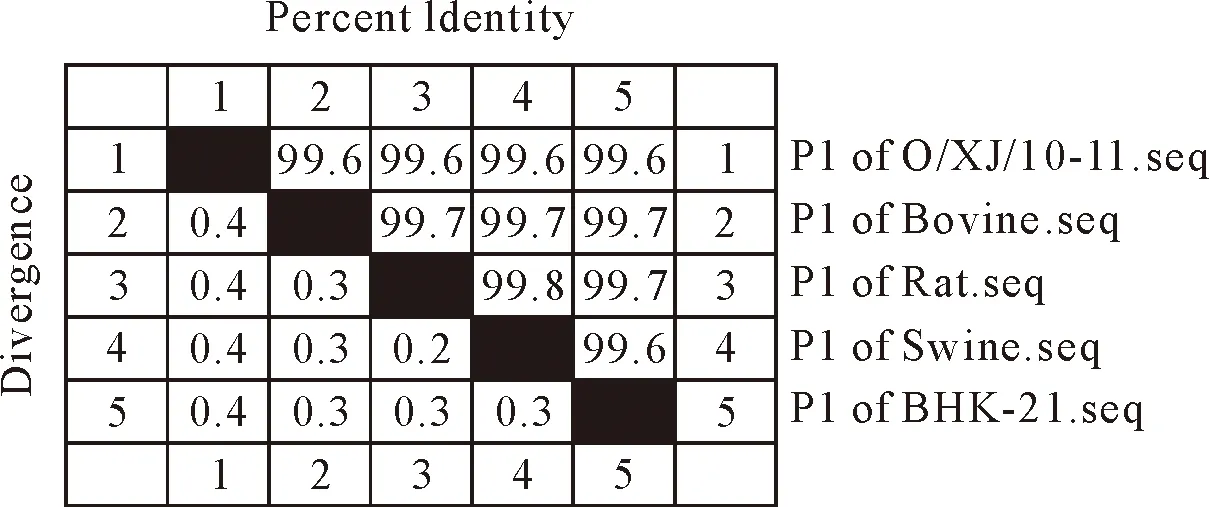
图3 P1基因核苷酸序列同源性
图4 P1基因氨基酸序列同源性

图5 P1基因氨基酸序列突变图
3 讨论
研究编码衣壳蛋白的P1基因,能够使我们进一步研究FMDV的相关基因功能,如P1基因在体外的功能特点及衣壳组装、非结构蛋白的功能等,而且P1区可能含有其他潜在的抗原表位,可能制备更好、更接近天然抗原的诊断试剂和基因工程疫苗,为今后建立安全、敏感、特异的诊断抗原以及新型疫苗提供新的思路和研究方向[14]。编码FMDV结构蛋白P1基因是构成病毒衣壳的基础,衣壳由于包含VP0、VP3和VP1,能诱导与感染性FMDV粒子相同的免疫反应。VP1、VP2、VP3基因位于衣壳表面,是楔形的,具有相似的结构,均由8个β-折叠桶和2个α-螺旋组成,每个β-折叠桶由2个4条链组成的β片层结构组成,是构成病毒衣壳的主要的抗原蛋白,与宿主的宿主范围和毒力有一定的联系。VP4基因位于FMDV粒子的内部[15]不易发生变异,与其保守性有关。
本研究将FMDV不同宿主适应毒进行比对分析,P1结构蛋白核苷酸序列同源性比较高(99.6%以上),氨基酸同源性在99.3%以上,未出现碱基缺失或增加现象。不同宿主适应毒的变异,均发生在P1衣壳表面蛋白上,主要集中于VP1的主要抗原区域的141-160位和200-213位,VP2抗原表位的156-166位和217-220位,关键氨基酸发生了明显的变化;VP3抗原蛋白也发生了氨基酸变异。对FMDV抗原表位的研究已经非常多,主要的研究集中于其结构蛋白VP1、VP2和VP3。但是由于这3个构蛋白的高变异性,因此其上的抗原表位也都有很大的变异性,基本都是特异性的抗原表位[16-17]。而结构蛋白VP4不仅在不同血清型之间体现出其保守性,而且在本试验中,在不同宿主适应毒之间也均未变异,也体现出VP4蛋白是极其保守的蛋白。与毒力相关基因的改变将引起病毒宿主范围和毒力方面的变化。对FMDV而言,柳纪省等[18]研究表明,VP1与FMDV的抗原性有关,而与宿主嗜性和持续性感染相关的基因可能存在于VP1之外。
本研究将缅甸98谱系FMDV在牛、乳鼠、猪、BHK-21细胞宿主进行攻毒试验并连续传代,分别获得各宿主适应毒,运用分子生物学技术及生物分析软件,分析不同宿主P1(1A、1B、1C、1D)内各段基因的变异情况,对FMDV宿主间遗传变异趋势、P1基因的结构特征与宿主关系有了新的认识,为今后研究口蹄疫在宿主间传染关系提供了基础信息。
[1] 刘运超,冯丽丽,赵宝磊,等.O型口蹄疫病毒VP3蛋白的可溶性表达与反应原性分析[J].河南农业科学,2015,44(11):124-128.[2] Jamal S M,Belsham G J.Foot-and-mouth disease:past,present and future[J].Vet Res,2013,44(44):1-14.
[3] Chitray M,Beer T A P D,Vosloo W,et al.Genetic heterogeneity in the leader and P1-coding regions of foot-and-mouth disease virus serotypes A and O in Africa[J].Arch Virol,2014,159(5):947-961.
[4] Vanlierop M J,Wagenaar J P,Vannoort J M,et al.Sequences derived from the highly antigenic VP1 region 140 to 160 of foot-and-mouth disease virus do not prime for a bovine T-cell response against intact virus[J].J Virol,1995,69(7):4511-4514.
[5] Mason P W,Grubman M J,Baxt B.Molecular basis of pathogenesis of FMDV[J].Virus Res,2003,91(1):9-32.
[6] Zheng C,Wang H,Zhou G,et al.Adenovirus-vectored type Asia1 foot-and-mouth disease virus (FMDV) capsid proteins as a vehicle to display a conserved,neutralising epitope of type O FMDV[J].J Virol Meth,2013,188(1-2):175-182.
[7] 陈启伟,王永录,张永光,等.口蹄疫病毒株AF72 VP2的结构模拟与分析[J].华北农学报,2009,24(4):1716-1722.
[8] Borca M V,Pacheco J M,Holinka L G,et al.Role of arginine-56 within the structural protein VP3 of foot-and-mouth disease virus (FMDV) O1 Campos in virus virulence[J].Virology,2012,422(1):37-45.
[9] 刘亚丽,丁耀忠,马小元,等.南非2型口蹄疫病毒VP1基因的核苷酸及其氨基酸序列分析[J].黑龙江畜牧兽医,2016(9):35-37.
[10] Borca M V,Pacheco J M,Holinka L G,et al.Role of arginine-56 within the structural protein VP3 of foot-and-mouth disease virus (FMDV) O1 Campos in virus virulence[J].Virology,2012,422(1):37-45.
[11] Das B,Mohapatra J K,Pande V,et al.Evolution of foot-and-mouth disease virus serotype A capsid coding (P1) region on a timescale of three decades in an endemic context.[J].Inf Gen Evol,2016,41:36-46.
[12] Pacheco J M,Gladue D P,Holinka L G,et al.A partial deletion in non-structural protein 3A can attenuate foot-and-mouth disease virus in cattle.[J].Virology,2013,446(1-2):260-267.
[13] Ahmeda H,Harbi H M,Habtoor A M.Compositional variations and tectonic settings of podiform chromitites and associated ultramafic rocks of the Neoproterozoic ophiolite at Wadi Al Hwanet,northwestern Saudi Arabia[J].J Asian Earth Sci,2012,56(3):118-134.
[14] 廖德芳,李华春.口蹄疫病毒结构蛋白P1研究进展[J].动物医学进展,2007,28(8):23-26.
[15] 吕 律,杨 帆,何继军,等.口蹄疫病毒O型Mya98谱系猪源毒株和牛源毒株全基因组的比较[J].中国兽医科学,2014(2):111-118.
[16] 栾喜梅,左小彤,黄梦菡,等.O型口蹄疫病毒VP4蛋白一个保守性表位的精细鉴定[J].复旦学报(自然科学版),2014,53(6):710-715.
[17] 李夏莹,郑鹭飞,张秀杰,等.口蹄疫病毒研究进展[J].中国畜牧兽医,2015,42(7):1910-1916.
[18] 柳纪省,梁沛勤,常惠芸,等.口蹄疫病毒在小鼠体内传代过程中基因突变的研究[C].家畜口蹄疫学术研讨会(第8次),2000 ,11 :58-59.
Study on Variations of P1 Genes of Different Host-Adaptive Type O Foot-and-mouth Disease Virus
YAO Huai-bing1,3,ZHAO Yi2,ZHU Yan1,LIU Meng-li1,LIU Hong2,REN Fang2,HUANG Jiong3
(1.VeterinaryMedicineCollegeofXinjiangAgriculturalUniversity,Urumqi,Xinjiang,830052China; 2.TeconBio-technologyCo,Urumqi,Xinjiang,830000,China;3.InstituteofVeterinaryMedicine,XinjiangAcademyofAnimalSciene,Urumqi,Xinjiang,830000,China)
In order to identify P1 gene sequence differences of different host-adaptive type O foot-and-mouth disease virus(FMDV) and study the trend of P1 genetic variation,FMDV was inoculated into cattle,neonatal mice,swine and BHK-21 cell line to obtain corresponding adapted virus.The RNA of FMDV in each adapted virus was taken as template for reverse transcription and amplification of P1(1A,1B,1C,1D) gene ,the sequences of each adapted virus amplified P1 gene were compared with each other.The results were as follows:different host-adapted virus nucleotide and amino acid sequences were no deletions;the mutation occurred in some of the key amino acids,the variation occurs in 141-160 and 213-200 in the main antigen region of VP1,156-166 and 217-220 in the antigen epitope region of VP2; less variation occurs in antigen protein region of VP3;no mutation occurs in region of VP4.The results provided important information for genome analysis of different host-adapted FMDV.
type O Foot-and-mouth disease virus; different host-adapted virus; P1 gene
2016-11-01
新疆维吾尔自治区科研机构创新发展专项资金(2016D04008);新疆维吾尔自治区产学研联合培养研究生示范基地项目(xjaucxy-yjs-20152008)
姚怀兵(1990-),男,新疆五家渠人,硕士研究生,主要从事动物口蹄疫病毒研究。*通讯作者
S852.659.6
A
1007-5038(2017)05-0011-06
