四种淡水缘毛类纤毛虫的形态学研究
2017-05-16姜传奇
姜传奇 缪 炜
(中国科学院水生生物研究所, 水生生物多样性与保护重点实验室, 武汉 430072)
四种淡水缘毛类纤毛虫的形态学研究
姜传奇 缪 炜
(中国科学院水生生物研究所, 水生生物多样性与保护重点实验室, 武汉 430072)
通过活体显微观察及银染法对采自武汉东湖的4种固着缘毛类纤毛虫: 粗茎睫纤虫Ophrydium crassicaule Penard, 1922, 念珠伪钟虫Pseudovorticella monilata (Tatem, 1870) Foissner & Schiffmann, 1974, 垂盖虫Opercularia nutans (Ehrenberg, 1831) Stein, 1854和一个鞘居虫未定种Vaginicola sp.的活体、纤毛图式、银线系以及细胞核的形态学特征进行详细的描述, 补充和完善了缘毛类纤毛虫的形态学资料, 为纤毛虫分类和系统发育学的研究提供重要信息。此外, 首次在伪钟虫中观察到有性生殖现象, 为纤毛虫生殖进化的研究提供了基础资料。
缘毛类纤毛虫; 形态学; 纤毛图式; 银线系
缘毛类纤毛虫隶属于纤毛门(Ciliophora)、寡膜纲(Oligohymenophorea)、缘毛亚纲(Peritrichia),由固着目(Sessilida)和游走目(Mobilida)组成, 广泛分布于淡水和海水环境[1,2]。17世纪, 荷兰博物学家列文虎克(Antonie van Leeuwenhoek, 1632—1723)在给英国皇家学会的那封著名的信件中描述到, “它由一根纤细的柄连接、固着在碎叶上, 虫体可以沿着螺旋形的轨迹收缩和伸展”, 经后人鉴定该单细胞生物就是缘毛类钟形虫(Vorticellid)。之后, Müller[3]、Ehrenberg[4,5]、Stokes[6—9]和Stein[10]等先后发表了大量的缘毛类纤毛虫的文章, 记录了一些缘毛类纤毛虫的活体形态图式。20世纪30年代, Kahl对当时已经发表的原生动物研究工作进行了总结, 发表了纤毛虫分类学的巨著《Urtiere oder Protozoa I: Wimpertiere oder Ciliata (Infusoria)》, 收录了缘毛类纤毛虫500余种[11]; 1992年, Foissner出版了纤毛虫分类学与生态学的著作, 其中记录了缘毛类纤毛虫51种[12]。在我国, 最早对缘毛类纤毛虫的研究见于1925年王家楫先生的《南京原生动物之研究》, 记录了10种缘毛类纤毛虫[13]; 1932年,王家楫和倪达书对4种缘毛类纤毛虫进行了详细的描述[14]; 1976年, 王家楫对我国32个废水处理厂展开调查, 发现了长期、稳定存在的一些优势缘毛类种群, 并提出将这些种群作为废水处理厂活性污泥的指示种[15]; 1977年, 王家楫[16]对西藏高原地区原生动物的调研过程中, 鉴定了缘毛类纤毛虫56种; 1980年以后, 沈蕴芬[17]、龚循矩[18—20]、徐振康[21,22]、余育和等[23]先后发表缘毛类新种/新亚种20多个; 宋微波对对虾体表的40多种病原性缘毛类的活体、表膜下纤毛图式及银线系进行了详细的描述[24—29]; 2009年, 宋微波等发表了《中国黄渤海的自由生纤毛虫》, 其中描述了64种缘毛类纤毛虫的活体特征、纤毛图式等重要的形态学资料[30]。2016年, 历时30多年完成的《中国动物志无脊椎动物第四十五卷纤毛门寡膜纲缘毛目》正式出版, 记录了我国38属507种缘毛类纤毛虫的分类检索、引证文献、形态特征和图片以及国内外地理分布情
况[31]。
当前, 缘毛类纤毛虫的研究主要包括传统形态学和分子系统发育学两个方面。但是, 随着分子系统发育学研究方法的日渐成熟和便捷, 关于缘毛类纤毛虫形态学的研究有被忽略的趋势, 尤其是大部分的缘毛类纤毛虫都不能或难于实验室内的稳定培养/纯培养, 导致很多种类缺少重要的活体、纤毛图式或银线系的形态学资料。然而分类学是研究生物的一种基本方法, 是生物学科的基础, 没有准确的鉴定, 其他领域的研究就难以深入展开, 因此缘毛类纤毛虫的形态学资料亟需详尽的补充和完善。
本文通过活体显微观察、荧光染料染色、蛋白银染色和干银法对3种缘毛类已知种进行了形态学重述, 补充了大量的通过现代染色技术获得的形态学显微照片及资料, 同时, 基于上述方法, 对鞘居虫科、鞘居虫属一未定种进行了形态学描述; 另外, 还观察到念珠伪钟虫的有性生殖现象。这些形态学研究为纤毛虫分类、系统发育学和有性生殖等深入研究提供重要基础。
1 材料与方法
4种淡水固着缘毛类纤毛虫: 粗茎睫纤虫(Ophrydium crassicaule)、念珠伪钟虫(Pseudovorticella monilata)、垂盖虫(Opercularia nutans)和鞘居虫未定种(Vaginicola sp.)均采自武汉东湖(N 30°32′, E 114°22′), 固着于水生植物金鱼藻(Ceratophyllum demersum)上。样品采集后, 在解剖镜(ZEISS Stemi 2000-C)下分离, 蛋白银染色见参考文献[32], 干银法见参考文献[33]。细胞核的形态以20%多聚甲醛固定细胞, 经DAPI或SYBR GreenⅠ核酸染料染色。显微观察及拍照利用尼康显微镜(Nikon ECLIPSE 80i)。名词术语和系统分类见参考文献[2]。
2 结果与讨论
2.1 粗茎睫纤虫Ophrydium crassicaule Penard, 1922(图版Ⅰ)
形态学描述 虫体(35—45) μm×(165—205) μm。呈细长花瓶形, 颈呈长圆柱形, 收缩性较强, 虫体后部膨大, 呈纺锤形(图版Ⅰ-1、2、6), 虫体收缩时呈球形或椭球形(图版Ⅰ-7)。游泳体长筒形, 反口纤毛环处膨大最宽(图版Ⅰ-8)。伸缩泡一个, 位于虫体中部(图版Ⅰ-2、6), 胞内有时有绿藻分布(图版Ⅰ-6、7)。大核细长绳状, 纵位, 小核一个, 椭球形,位于细胞反口端、大核的末端(图版Ⅰ-2、9)。柄纤细线状, 不能收缩, 二叉式分枝(图版Ⅰ-1、6)。虫体由胶质囊包裹, 群体一般呈不规则半球形, 较大群体呈球形(图版Ⅰ-5)。
口器结构如图(图版Ⅰ-3、4、13—16), 口区单动基列(HK)、复动基列(PK)围绕口围盘旋转约 1.5圈后进入口前庭, 在口前庭内继续旋转约一圈。PK继续延伸形成三片口区小膜(P1、P2、P3), 均由三列动基列组成。P1和P2内侧靠近P1一列动基列从PK延伸出来, P1下端三列动基列等长, 和P3的外侧两列动基列在口前庭末端汇合并同时终止。P2的上端由内而外依次略短, 下端的三列动基列几乎等长, 终止于P1与P3的汇合处。P3上端接近P2中下部, 几乎等长, 下端内侧靠近P2一列比P2略长, 外侧两列终止于P1 的末端。芽动基列(GK)位于口前庭上部, 与HK平行。
反口纤毛环(A)由围绕在虫体反口端约五分之一处的一圈双动基系构成(图版Ⅰ-11、12)。纵向纤维粗壮(图版Ⅰ-11、12), 口端起始于复动基列下方, 形成纤维束网络向反口端延伸至帚胚终止, 使得虫体具较强的伸缩性。银线系呈横纹型, 排列均匀(图版Ⅰ-10), 从虫体口端到反口纤毛环银线数110—120条, 反口纤毛环到帚胚银线数为22—28条。
比较与讨论 本种由Penard于1922年首次报道, Kahl[11]、Nusch[34]、Guhl[35]及沈蕴芬和顾曼如[31]也对该种个体大小、大核、伸缩泡及游泳体等活体形态进行了补充和重述, 本文记述的O. crassicaule的形态特征与前人描述基本吻合。本工作补充小核的形态和位置, 并通过蛋白银法获得了表膜下纤毛图式, 给出了三片口区小膜的排布模式, 以及干银法获得了银线系特征。
2.2 念珠伪钟虫Pseudovorticella monilata (Tatem, 1870) Foissner & Schiffmann, 1974(图版Ⅱ)
形态学描述 虫体(50—65) μm×(55—75) μm,倒钟形, 伸展时体长比体宽略大, 长宽比约为1.1:1,口围向外扩张, 为虫体最宽处(图版Ⅱ-1、6), 收缩时呈球形(图版Ⅱ-2)。胞咽延伸至虫体的中部, 伸缩泡2个, 位于口前庭入口和口前庭与胞咽之间(图版Ⅱ-1、8)。大核“J”型, 纵位, 两端稍弯(图版Ⅱ-1、12、13)。小核一个, 椭球形, 位于虫体中上部,靠近大核(图版Ⅱ-1、12)。游泳体圆筒形(图版Ⅱ-3、7), 收缩时球形(图版Ⅱ-8)。表膜有明显的呈砖纹状排列的泡状结构(图版Ⅱ-1、2、6—8)。
口器结构如图(图版Ⅱ-4、5、18—22), 口区单(HK)、复动基列(PK)围绕口围盘旋转约1.25圈后进入口前庭, 三片口区小膜(P1、P2、P3)均由三排动基列组成, P1最长, P3最短, 只有P2的三分之一左右。P1和P2内侧靠近P1的一列动基列由PK延伸出来, P1的下端三列动基列等长。P2的上端外侧两列动基列几乎等长, 比内侧一列短, 下端的三列动基列几乎等长, 终止于P1和P3汇合处。P3三列动基列上端几乎等长, 下端内侧靠近P2一列比P2的下端稍长, 外侧两列比内侧一列略长、与P1汇聚终止于口前庭末端。微丝网结构(FR)发达与口区小膜平行。芽动基列(GK)位于口前庭上部, 与HK平行。
反口纤毛环(A)由围绕在虫体反口端约四分之一处的一圈双动基系构成(图版Ⅱ-16、17)。纵向纤维纤细, 由口区至帚胚贯穿整个细胞(图版Ⅱ-14、15)。银线系呈砖纹型(图版Ⅱ-17)。从虫体口端到反口纤毛环处的银线数20—24条, 反口纤毛环到帚胚的银线数14—17条。
本文还观察到念珠伪钟虫的有性生殖现象(图版Ⅱ-9—11)。该种大配子有柄固着, 与营养时期细胞的形态和大小类似; 小配子自由游泳, 球形, 约为营养细胞体积的四分之一。在接合过程中, 小配子的反口端与大配子接合, 并将其细胞核和细胞质注入大配子胞内, 小配子的细胞膜干瘪, 最终脱落。大、小配子永久性的融合成一个细胞, 进行合子细胞的发育, 再形成子代细胞。
比较与讨论 本种分布广泛, 是伪钟虫属较为常见的种类。由Tatem于1870年首次报道, 1974年, Foissner和Schiffmann对其修订, 将该种从钟虫属移入伪钟虫属[36]。Kahl[11]、Song和Wilbert[37]、Foissner等[12]多位学者对该种活体形态、银线系以及简单的纤毛图式进行了描述, 本文所述P. monilata的活体形态和银线系与前人描述基本吻合。本工作补充了小核的形态和位置, 以及三片口区小膜详细的排布模式, 并首次记述了该种的有性生殖现象。
2.3 垂盖虫Opercularia nutans (Ehrenberg, 1831) Stein, 1854(图版Ⅲ)
形态学描述 虫体(25—45) μm×(75—90) μm,虫体伸展时呈纺锤形, 中部最宽, 口端与反口端变细(图版Ⅲ-1、3、5、6)。口围缘不膨大为口围唇,口区纤毛长而密。口围盘略小于口围, 高耸于口围之上(图版Ⅲ-1、3、5、6)。口前庭、胞咽发达, 达虫体中部(图版Ⅲ-1、3、6)。伸缩泡一个, 靠前庭末端和胞咽之间(图版Ⅲ-1、3、5), 大核短C形, 横位于虫体上半部, 小核球形, 贴近大核中部(图版Ⅲ-3、9)。虫体收缩时口端有突起, 反口端有3—4个褶皱(图版Ⅲ-7)。柄二叉式分枝(图版Ⅲ-2), 具有微弱纵纹, 柄基部无横纹, 二级及以上的柄具有明显环状横纹(图版Ⅲ-1、8)。
口器结构如图(图版Ⅲ-4、15—17)。银染固定过程中, 虫体剧烈收缩, 口围盘及口纤毛缩入胞内至虫体中上部三分之一处。口区单(HK)、复动基列(PK)围绕口围盘旋转约1.25圈后进入口前庭, PK在口前庭的下半部分成三片口区小膜(P1、P2、P3), 由三列动基列组成。P1上端的三列动基列与P2内侧靠近P1的一列一同由PK延伸而出, P1三列动基列下端等长, 终止于口前庭末端。P2上端外侧的两列动基列由内而外依次稍短于内侧一列, 三列动基列的下端靠近P1的一列最短, 外侧两列等长,短于P1。P3最短, 上端靠近P2的一列略短, 外侧的两列等长, 下端由内而外依次略长, 不与P1汇合。芽动基列(GK)位于口前庭上方。
反口纤毛环(A)由围绕在虫体后五分之一到六分之一处的一圈双动基系构成(图版Ⅲ-12)。纵向纤维不连续, 口端的纵向纤维起始于PK的下方, 多数由2—3组“爪”状纤维组成, 每个“爪”状纤维末端有5—7个分枝(图版Ⅲ-10、11)。反口端纵向纤维自帚胚穿过反口纤毛环向虫体中部延伸(图版Ⅲ-11、13)。银线系呈横纹型(图版Ⅲ-14), 虫体口端到反口纤毛环之间的银线数70—76条, 反口纤毛环到帚胚之间的银线数为20—22条。
比较与讨论 本种由Ehrenberg于1831年首次报道, 1854年Stein将其修订, 由累枝虫属移入盖虫属[38]。Kahl[11]、Nusch[34]、Foissner和Schiffmann[36],及沈蕴芬和顾曼如[31]等学者对该种形态学特征进行了补充和重述, 本工作补充了小核的形态及位置,并给出了详细的表膜下纤毛图式。除银线系特征与Foissner和Schiffmann[36]的描述有一定差异(口端到反口纤毛环的银线数105—122 vs. 70—76条, 反口纤毛环到帚胚的银线数25—35 vs. 20—22条)外,本文记述的O. nutans的形态特征与前人的描述基本一致。
2.4 鞘居虫未定种(Vaginicola sp.)(图版Ⅳ)
形态学描述 鞘(70—75) μm×(210—250) μm,无色透明, 呈长圆柱形, 鞘口微张, 为鞘的最宽处,鞘后端三分之一处开始呈圆锥形缩细, 鞘末端缩成小圆柱形, 直接以末端固着于基质上(图版Ⅳ-1、2、5—7)。鞘内1—2个个体, 虫体(40—60) μm× (280—360) μm, 透明, 细长喇叭形(图版Ⅳ-1、5),表膜具微弱横纹, 口围唇(PL)处最宽, 虫体末端纤细, 有一短柄, 以短柄固着于鞘底, 柄不伸出鞘外(图版Ⅳ-1、2、5—7)。虫体收缩时缩入鞘内, 约为鞘长的四分之三(图版Ⅳ-2、6)。伸缩泡一个, 顶位,靠近口围唇(图版Ⅳ-1、5)。大核细长绳状, 纵位(图版Ⅳ-1、7)。
口器结构如图(图版Ⅳ-3、4、10—12)。口端单(HK)、复动基列(PK)绕口围盘旋转约1.5圈, 三片口区小膜(P1、P2、P3)均由三排动基列组成。P1和P2内侧靠近P1的一列起始于PK, P1的下端等长, 与P3外侧两列汇合终止。P2的上端外侧两列由内而外依次略短, 下端靠近P1一列较短, 外侧的两列等长, 终止于P1和P3汇合处。P3上端靠近P2一列较长, 约为P2的二分之一, 外侧两列几乎等长, 约为内侧一列的二分之一, 下端内侧一列比P2的外侧两列稍长, 外侧两列几乎等长, 与P1汇合终止于口前庭末端。芽动基列(GK)位于口前庭上部, 与HK平行。
反口纤毛环(A)位于反口端约三分之一处(图版Ⅳ-8)。口端纵向纤维短但较为粗壮, 反口端纵向纤维束的排布较细密, 自帚胚延伸穿过反口纤毛环至虫体中上部渐细(图版Ⅳ-8、9)。银线系呈横纹型(图版Ⅳ-14), 从虫体口端到反口纤毛环处的银线数110—120条, 反口纤毛环到帚胚之间的银线数为80—90条。
比较与讨论 鞘居虫属(Vaginicola), Lamarck于1816年建立, 该属鞘外无柄, 直接以鞘的后端固着于基质上, 虫体直接或以内柄固着于鞘底, 已记录有50余种。在鞘居虫中, 有内柄且大核绳状的淡水种有2个, 纵长鞘居虫(V. elongata)和小口鞘居虫(V. leptosoma)[11,31]。本文记述的鞘居虫未定种区别于以上2个相似种的主要特征在于其鞘形态的特殊性, 本种鞘呈长圆柱形, 鞘口微张, 后端呈圆锥形缩细, 末端缩成小圆柱形, 以鞘末端固着于基质上, 该种鞘的形态在鞘居虫中是罕见的。V. elongate鞘呈粗钝瓶形, 鞘后端三分之一处最宽, 且鞘长约为60 μm(vs. 210—250 μm); V. leptosoma鞘形态也比较特殊, 是该种的主要分类特征, 鞘呈纺锤形, 中部为最宽处, 两端缩细, 以纤细的鞘底固着于基质的表面。在鞘居虫科扉门虫属(Thuricola)中, 虽然有些种鞘的外形与本种类似, 如圆锥扉门虫(T. obconica)、盖氏扉门虫(T. kellicottiana)[11,31], 但最明显的差别在于扉门虫鞘的内壁有膜盖, 而本文所述的鞘居虫的鞘内无膜盖。
3 总结
通过显微观察和现代染色技术对3科3种固着缘毛类的形态学研究, 在活体形态学重述的基础上,补充了小核的形态和位置, 并获得了详细的表膜下纤毛图式和银线系等重要鉴别性状, 有力的补充和完善了形态学资料。同时, 还记述了鞘居虫科、鞘居虫属一未定种, 本文对该种的活体形态、纤毛图式和银线系进行了详细的描述, 为缘毛类纤毛虫形态分类和系统发育学研究提供重要的基础资料。另外, 本文首次记录了伪钟虫的有性生殖现象, 为深入研究缘毛类纤毛虫有性生殖过程提供了基础资料, 而且这种大、小配子融合成一个细胞再发育成子代细胞与高等动物精卵细胞的融合是相似的,也为研讨生殖进化提供线索。
[1]Corliss J O. The Ciliated Protozoa: Characterization, Classification and Guide to the Literature [M]. Oxford: Pergamon Press. 1979, 1—455
[2]Lynn D H. The Ciliated Protozoa. Characterization, Classification, and Guide to the Literature (3rd edition) [M]. Springer: Dordrecht. 2008, 1—605
[3]Müller O F. Vermium terrestrium et fluviatilium, seu Amimalium Infusorium, Helminthicorum et Testaceorum, non Marinorum, Succincta Historia (Vol.1) [M]. Havniae et Lipsiae, Heineck & Faber. 1773, 1—135
[4]Ehrenberg C G. Beiträge zur Kenntniss der Organisation der Infusorien und ihrer geographischen Verbreitung, besonders in Sibirien. Abh. Akad. Wiss. Berlin. 1830, 1—88
[5]Ehrenberg C G. Über die Entwickelung und Lebensdauer der Infusionthiere; nebst ferneren Beiträgen zu einer Vergleichung ihier Organischen Systeme. Abh. Akad. Wiss. Berlin. 1831, 1—154
[6]Stokes A C. A new Thuricola [J]. American Monthly Microscopical Journal, 1882, 3: 182—183
[7]Stokes A C. A new Vorticella [J]. American Monthly Microscopical Journal, 1883, 4: 208
[8]Stokes A C. A new infusorien belonging to the Genus Vorticella [J]. American Naturalist, 1884, 18: 829—830
[9]Stokes A C. Some new infusoria from freshwater [J]. American Naturalist, 1885, 19: 18—27
[10]Stein F. Neue beiträge zue kennteiss der entwicklungsgeschichte und des feineren baues der infusionsthiere [J]. Zeitschrift für Wissenschaftliche Zoologie, 1851, 3: 475—509
[11]Kahl A. Urtiere oder Protozoa I: Wimpertiere oder Ciliata (Infusoria) 4. Peritricha und Chonotricha [M]. Tierwelt Dtl. 1935, 651—886
[12]Foissner W, Berger H, Kohmann F. Taxonomische und ölogische Revision der Ciliaten des Saprobiensystems—Band II: Peritrichia, Heterotrichida, Odontostomatida [M]. Informationsberichte des Bayerischen Landesamtes für Wasserwirtschaft. 1992, 1—502
[13]Wang C C. Study of the protozoa of Nanking. Contributions from the Biological Laboratory of the Science Society of China. 1925, 1(3): 1—60
[14]Wang C C, Nie D S. A survey of the marine protozoa of Amoy. Contributions from the Biological Laboratory of the Science Society of China, Zoological Series. 1932, 8(9): 285—385
[15]Wang J J, Shen Y F, Gong X J. Illustrated Microzoologial Fauna in Biological Wastewater [M]. Beijing: Architechture and Building Press. 1976, 99—129 [王家楫,沈韫芬, 龚循矩. 废水生物处理微型动物图志. 北京: 中国建筑工业出版社. 1976, 99—129]
[16]Wang J J. Prototoa from some districts of Tibetan Plauteau [J]. Acta Zoologica Sinica, 1977, 23(2): 131—160 [王家楫. 西藏高原部分地区的原生动物. 动物学报, 1977, 23(2): 131—160]
[17]Shen Y F. Descriptions of six new species of periphytic protozoa in Lake Donghu of Wuhan [J]. Acta Hydrobiologia Sinica, 1980, 7(2): 245—251 [沈韫芬. 武汉东湖周丛原生动物六新种. 水生生物学集刊, 1980, 7(2): 245—251]
[18]Gong X J. Descriptions of six new species of Preitrich from Xinjiang, China [J]. Acta Zootaxonomica Sinica, 1989, 14(4): 396—403 [龚循矩. 新疆缘毛目六新种记述. 动物分类学报, 1989, 14(4): 396—403]
[19]Gong X J. Descriptions of three new species of Preitrich from Qinghai Province, China [J]. Acta Zootaxonomica Sinica, 1989, 14(3): 257—261 [龚循矩. 青海缘毛目三新种. 动物分类学报, 1989, 14(3): 257—261]
[20]Gong X J, Xiao H Z, Shen Y F. Studies on the protozoa fauna of Three Gorge area in Changjiang (the Yangtze) River [J]. Acta Hydrobiologica Sinica, 1990, 14(4): 289— 297 [龚循矩, 肖化忠, 沈韫芬. 长江三峡地区的原生动物区系研究. 水生生物学报, 1990, 14(4): 289—297]
[21]Xu Z K. Two new species of the Family Vaginicolidae from Tianjin [J]. Acta Zootaxonomica Sinica, 1987, 12(1): 5—9 [徐振康. 天津市鞘居科纤毛虫二新种. 动物分类学报, 1987, 12(1): 5—9]
[22]Xu Z K. Notes on two new species of genus Thuricola from Tianjin [J]. Acta Zootaxonomica Sinica, 1990, 15(1): 6—9 [徐振康. 天津扉门虫属二新种记述缘毛目:鞘居科. 动物分类学报, 1990, 15(1): 6—9]
[23]Yu Y H, Feng W S, Shen Y F. Description of Campanella hanchuanensis sp. nov. (Ciliophora: Peritrichida): general morphology and isozyme pattern [J]. Report of Suwa Hydrobiology, 1995, 9: 87—94
[24]Song W B. Descriptions of seven new speices of peritrichs on Penaeus orientalis (Peritricha: Zoothamnidae: Epistylidae) [J]. Acta Zootaxonomica Sinica, 1986, 11(3): 225—235 [宋微波. 中国对虾体表共栖纤毛虫七新种(缘毛目: 聚缩虫科, 累枝虫科). 动物分类学报, 1986, 11(3): 225—235]
[25]Song W B. A new commensal ciliate, Zoothamnium paraentzii (Ciliophora: Peritrichida) [J]. Zoological Research, 1991, 12(4): 355—359 [宋微波. 对虾体表共栖纤毛虫一新种—拟恩茨聚缩虫(纤毛动物门: 缘毛目).动物学研究, 1991, 12(4): 355—359]
[26]Song W B. Contribution to the commensal ciliates on Penaeus orientalis Ⅰ. (Ciliophora: Peritrichida) [J]. Journal of Ocean University of Qingdao, 1991, 21(3): 119—128 [宋微波. 共栖于对虾体表的致病性纤毛虫Ⅰ.(纤毛动物门: 缘毛目). 青岛海洋大学学报, 1991, 21(3): 119—128]
[27]Song W B. Contribution to the commensal ciliates on Penaeus orientalis Ⅱ. (Ciliophora: Peritrichida) [J]. Journal of Ocean University of Qingdao, 1991, 21(4): 45—55 [宋微波. 共栖于对虾体表的致病性纤毛虫Ⅱ.(纤毛动物门: 缘毛目). 青岛海洋大学学报, 1991, 21(4): 45—55]
[28]Song W B. A new marine ciliates, Zoothamnium pnaei sp. nov. (Ciliophora: Peritrichida) [J]. Oceanologia et Limnologia Sinica, 1992, 23(1): 90—94 [宋微波. 海洋纤毛虫一新种—对虾聚缩虫. 海洋与湖沼, 1992, 23(1): 90—94]
[29]Song W B. Contribution to the commensal cliates on Penaeus orientalis Ⅲ. (Ciliophora: Peritrichida) [J]. Journal of Ocean University of Qingdao, 1992, 22(2): 107—116 [宋微波. 共栖于对虾体表的致病性纤毛虫Ⅲ.(纤毛动物门: 缘毛目). 青岛海洋大学学报, 1992, 22(2): 107—117]
[30]Song W B, Warren A, Hu X Z. Free—living Ciliates in the Bohai and Yellow Seas, China [M]. Beijing: Academic Press. 2009, 217—286 [宋微波, A. 沃伦, 胡晓钟. 中国黄渤海的自由生纤毛虫. 北京: 科学出版社. 2009, 217—286]
[31]Shen Y F, Gu M R. Fauna Sinica: Invertebrata Ciliophora Oligohymenophorea Peritrichida [M]. Beijing: Science Press. 2016, 669 [沈蕴芬, 顾蔓茹. 中国动物志无脊椎动物第四十五卷纤毛门寡膜纲缘毛目. 北京: 科学出版社. 2016, 669]
[32]Shi X B, Frankel J. Morphology and development of mirror image doubles of Stylonychia mytilus [J]. Journal of Protozoology, 1990, 37: 1—13
[33]Klein B M. Weitere Beiträge zur Kenntnis des Silberliniensystems der Ciliaten [J]. Archiv für Protistenkunde, 1929, 65: 183—257
[34]Nusch E A. Ökologische und systematische Untersuchungen der Peritricha (Protozoa, Ciliata) im Aufwuchs von Talsperren und Flußstauen mit verschiedenem Saprobitätsgrad (mit Modellversuchen) [J]. Archiv. Fur Hydrobiologie, Supplementband, 1970, 37: 243—386
[35]Guhl W. Revision der Peritrichen familie Ophrydiidae [J]. Archiv für Protistenkunde, 1985, 129: 127—143
[36]Foissner W, Schiffmann H. Vergleichende Studien an argyrophilen Strukturen von vierzehn peritrichen Ciliaten [J]. Protistologica, 1974, 489—508
[37]Song W, Wilbert N. Taxonomische Untersuchungen an Aufwuchsciliaten (Protozoa, Ciliophora) im Poppelsdorfer Weiher, Bonn. Lauterbornia. 1989, 3: 1—222
[38]Stein F. Die Infusionsthiere auf ihre Entwickelungsgeschichte untersucht [M]. Leipzig: W. Engelmann. 1854, 1—265
MORPHOLOGY OF FOUR FRESHWATER PERITRICHOUS CILIATES
JIANG Chuan-Qi and MIAO Wei
(Key Laboratory of Aquatic Biodiversity and Conservation, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China)
Morphology of four sessile peritrichous ciliates from Donghu Lake, including Ophrydium crassicaule Penard, 1922, Pseudovorticella monilata (Tatem, 1870) Foissner & Schiffmann, 1974, Opercularia nutants (Ehrenberg,1831) Stein, 1854 and an unidentified species Vaginicola sp., were investigated through living observation, protargol staining and silver impregnation. The detailed descriptions of living characters, infraciliature, silverline systems and dimorphic nuclei morphology were presented. These results reinforce the morphology data of peritrichous ciliates, and provide important information to the taxonomy and phylogeny of ciliates. Furthermore, the first report of conjugation process of Pseudovorticella provided the basic knowledge for studying the sexual reproduction of ciliates.
Peritrichous ciliates; Morphology; Infraciliature; Silverline system

图版Ⅰ 粗茎睫纤虫的形态PlateⅠ Morphology of Ophrydium crassicaule1. 群体形态; 2. 个体形态; 3. 口区纤毛图式; 4. 口区小膜的排布; 5. 群体形态; 6. 伸展个员, 箭头指示伸缩泡; 7. 收缩个员; 8. 游泳体,箭头指示反口纤毛环; 9. 大核和小核; 10. 银线系, 箭头指示反口纤毛环; 11. 纵向纤维; 12. 反口端纵向纤维及帚胚; 13. 单动基列和复动基列; 14. 小膜1和小膜2; 15. 小膜1—3的排布; 16. 小膜1和小膜3, 箭头指示小膜1下端的三列动基列。A, 反口纤毛环; CV, 伸缩泡; GK, 芽动基列; HK, 单动基列; MA, 大核; MI, 小核; PK, 复动基列; P1–3, 小膜 1—3; SC, 帚胚; ST, 柄。标尺: 50 μm1. Colony form; 2. Zooid form; 3. Oral infraciliature; 4. The detailed arrangement of infundibular polykineties; 5. Colony in vivo; 6. Expanded zooid in vivo, arrow marks contractile vacuole; 7. Contracted zooids in vivo; 8. Swimmer in vivo, arrow marks aboral ciliary wreath; 9. Macronucleus and micronucleus; 10. Silverline pattern, arrows mark aboral ciliary wreath; 11. Longitudinal fibers; 12. Longitudinal fibers at distal end and scopula; 13. Haplokinety and polykinety; 14. Infundibular polykineties1 and 2; 15. Arrangement of infundibular polykineties 1—3; 16. Infundibular polykineties 1 and 3, arrows mark the end of 3 rows of infundibular polykinety 1. A, aboral ciliary wreath; CV, contractile vacuole; GK, germinal kinety; HK, haplokinety; MA, macronucleus; MI, micronucleus; PK, polykinety; P1–3, infundibular polykineties 1—3; SC, scopula; ST, stalk. Scale bars: 50 μm
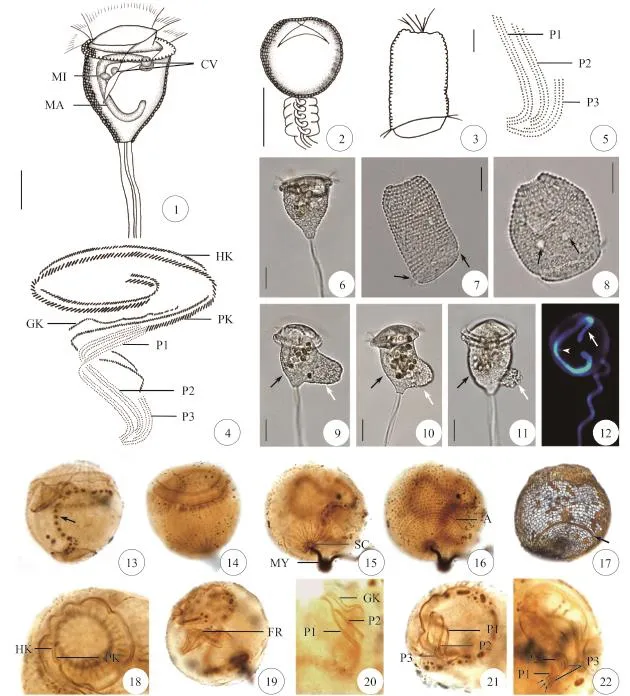
图版Ⅱ 念珠伪钟虫的形态PlateⅡ Morphology of Pseudovorticella monilata1. 伸展个员; 2. 收缩个员; 3. 游泳体; 4. 口区纤毛图式; 5. 口区小膜的排布; 6. 伸展个员; 7. 伸展的游泳体, 箭头指示反口纤毛环; 8. 收缩的游泳体, 箭头指示伸缩泡; 9—11. 接合生殖, 黑色箭头指示大配子, 白色箭头指示小配子; 12. 大核(箭头)和小核(无尾箭头); 13. 蛋白银染色大核(箭头); 14. 纵向纤维; 15. 反口端纵向纤维及帚胚; 16. 反口纤毛环; 17. 银线系, 箭头指示反口纤毛环; 18. 单动基列和复动基列; 19. 微丝网结构; 20. 小膜1–2和芽动基列; 21. 小膜1—3的排布; 22. 小膜1—3, 箭头指示小膜1下端的三列动基列。A, 反口纤毛环; CV, 伸缩泡; FR, 微丝网; GK, 芽动基列; HK, 单动基列; MA, 大核; MI, 小核; MY, 肌丝; PK, 复动基列; P1–3, 小膜 1—3; SC, 帚胚。标尺: 20 μm1. Expanded zooid; 2. Contracted zooid; 3. Swimmer; 4. Oral infraciliature; 5. The detailed arrangement of infundibular polykineties; 6. Expanded zooid in vivo; 7. Expanded swimmer in vivo, arrows mark aboral ciliary wreath; 8. Contracted swimmer in vivo, arrows mark contractile vacuoles; 9—11. Conjugation processes, black arrows mark macroconjugants and white arrows mark microconjugants; 12. Macronucleus (arrow) and micronucleus (arrow head); 13. Macronucleus after protargol impregnation (arrow); 14. Longitudinal fibers; 15. Longitudinal fibers at distal end and scopula; 16. Aboral ciliary wreath; 17. Silverline pattern, arrows mark aboral ciliary wreath; 18. Haplokinety and polykinety; 19. Filamentous reticulum; 20. Infundibular polykineties 1–2 and germinal kinety; 21. Arrangement of infundibular polykineties 1—3; 22. Infundibular polykineties 1—3, arrows mark the end of 3 rows of infundibular polykinety 1. A, aboral ciliary wreath; CV, contractile vacuole; FR, filamentous reticulum; GK, germinal kinety; HK, haplokinety; MA, macronucleus; MI, micronucleus; MY, myoneme; PK, polykinety; P1–3, infundibular polykineties 1—3; SC, scopula. Scale bars: 20 μm

图版Ⅲ 垂盖虫的形态PlateⅢ Morphology of Opercularia nutans1. 群体形态; 2. 二叉式分枝的柄; 3. 个体形态; 4. 口区纤毛图式; 5. 群体形态, 箭头指示伸缩泡; 6. 伸展个员和胞咽; 7. 收缩个员; 8. 柄的环纹和纵纹; 9. 大核(箭头)和小核(无尾箭头); 10—11. 纵向纤维, 箭头指示口端纵向纤维末端的指状分枝; 12. 反口纤毛环; 13. 纵向纤维和帚胚(箭头); 14. 银线系, 箭头指示反口纤毛环; 15. 单动基列和复动基列; 16. 芽动基列; 17. 小膜1—3的排布。A, 反口纤毛环; CV, 伸缩泡; GK, 芽动基列; HK, 单动基列; MA, 大核; MI, 小核; CPH, 胞咽; PK, 复动基列; P1—3, 小膜 1—3; SC, 帚胚。标尺: 50 μm1. Colony form; 2. Dichotomously branched stalk; 3. Zooid form; 4. Oral infraciliature; 5. Colony in vivo, arrow marks contractile vacuole; 6. Expanded zooids and cytopharynx; 7. Contracted zooids in vivo; 8. Longitudinal and transverse striations on the stalk; 9. Macronucleus (arrow) and micronucleus (arrow head); 10—11. Longitudinal fibers, arrows mark the end of palmately branches at anterior end; 12. Aboral ciliary wreath; 13. Longitudinal fibers and the scopula (arrow); 14. Silverline pattern, arrow marks aboral ciliary wreath; 15. Haplokinety and polykinety; 16. Germinal kinety; 17. Arrangement of infundibular polykineties 1—3. A, aboral ciliary wreath; CV, contractile vacuole; GK, germinal kinety; HK, haplokinety; MA, macronucleus; MI, micronucleus; CPH, cytopharynx; PK, polykinety; P1—3, infundibular polykineties 1—3; SC, scopula. Scale bars: 50 μm

图版Ⅳ 鞘居虫未定种的形态PlateⅣ Morphology of Vaginicola sp.1. 伸展个员; 2. 收缩个员; 3. 口区纤毛图式; 4. 口区小膜的排布; 5. 伸展个员, 箭头指示伸缩泡; 6. 收缩个员; 7. 蛋白银染色个员; 8. 纵向纤维, 箭头指示反口纤毛环; 9. 口端纵向纤维; 10. 单动基列, 复动基列, 芽动基列和小膜1—2; 11. 小膜1—3的排布; 12. 小膜1—3,箭头指示小膜3上端的三列动基列。13. 表膜横纹; 14. 银线系。A, 反口纤毛环; CV, 伸缩泡; GK, 芽动基列; HK, 单动基列; LO, 鞘; MA, 大核; PK, 复动基列; P1–3, 小膜 1—3; ST, 柄。标尺: 50 μm1. Expanded colony; 2. Contracted colony; 3. Oral infraciliature; 4. The detailed arrangement of infundibular polykineties; 5. Expanded zooids in vivo, arrow marks contractile vacuole; 6. Contracted zooids in vivo; 7. Colony after protargol impregnation; 8. Longitudinal fibers, arrows mark aboral ciliary wreath; 9. Longitudinal fibers at anterior end; 10. Haplokinety, polykinety, germinal kinety and infundibular polykineties 1—2; 11. Arrangement of infundibular polykineties 1—3; 12. Infundibular polykineties 1—3, arrows mark the upper end of 3 rows of infundibular polykinety 3; 13. Transverse pellicular striations after protargol impregnation; 14. Silverline pattern. A, aboral ciliary wreath; CV, contractile vacuole; GK, germinal kinety; HK, haplokinety; LO, lorica; MA, macronucleus; PK, polykinety; P1—3, infundibular polykineties 1—3; ST, stalk. Scale bars: 50 μm
Q959.116
A
1000-3207(2017)03-0652-09
10.7541/2017.83
2016-09-26;
2016-12-26
国家自然科学基金(No. 31372168)资助 [Supported by the National Natural Science Foundation of China (No. 31372168)]
姜传奇(1988—), 女, 黑龙江伊春人; 博士研究生; 研究方向原生动物学。E-mail: jiang_chuanqi@163.com
缪炜(1974—), 男; 研究员; E-mail: miaowei@ihb.ac.cn
