5-氮杂-2-脱氧胞苷对百草枯诱导SH-SY5Y细胞神经毒性的影响
2017-05-10石恩林李重阳
王 炜 侯 艳 石恩林 李重阳
(甘肃政法学院公安技术学院,甘肃 兰州 730070)
5-氮杂-2-脱氧胞苷对百草枯诱导SH-SY5Y细胞神经毒性的影响
王 炜1侯 艳2石恩林 李重阳1
(甘肃政法学院公安技术学院,甘肃 兰州 730070)
目的 探讨甲基转移酶抑制剂5-氮杂-2-脱氧胞苷(5′-aza-dC)对百草枯所诱导的SH-SY5Y细胞神经毒性的影响。方法 SH-SY5Y细胞预先用5′-aza-dC处理24 h,然后暴露于百草枯12 h。对细胞凋亡比例、相关蛋白Bcl-2和Bax表达以及培养液上清中活性氧簇(ROS)进行检测。结果 和百草枯加5′-aza-dC干预后SH-SY5Y细胞活性显著降低,凋亡增加。此外,与单独百草枯处理相比,在暴露于百草枯加5′-aza-dC的联合作用后,ROS水平显著增加(P<0.05)。抗凋亡蛋白Bcl-2表达降低,凋亡诱导蛋白Bax表达增加,Bcl-2/Bax的比例下降,同时,细胞色素C的表达增加。结论 5′-aza-dC通过干预调节DNA甲基化,引起细胞氧化应激增加,并启动线粒体凋亡途径,这些进一步加重百草枯对SH-SY5Y细胞的神经毒作用。
5-氮杂-2-脱氧胞苷;百草枯;帕金森病;凋亡;氧化应激
环境因素长久以来一直被认为可能参与帕金森病(PD)的发病,这是由于神经毒素优先破坏神经系统内多巴胺黑质纹状体通路。职业性接触农药和持续暴露于除草剂如百草枯可以增加患PD的风险〔1,2〕。但是,目前尚无单一环境中有害物质可引起PD的报道,说明其他风险因素必然参与散发性PD的发展。尽管PD也有遗传因素影响,导致部分人群存在PD易感性〔3〕。然而环境中一些潜在的可疑化合物对细胞的功能和代谢存在长期影响,这些化合物的长期暴露可以导致表观遗传学的改变,进而影响相关基因的表达,由于表观遗传学主要影响基因表达而对DNA序列基本无影响,因此是属于动态可变的调节因素,已经有不少的研究显示,基因启动子的甲基化水平不仅反映人衰老的程度,并可以促进衰老〔4〕。一些神经元保护性基因启动子区域的异常高甲基化,可以引起相关功能基因表达下调,因此,长期暴露在有毒化合物质中的人体是否会引起DNA甲基化的异常修饰,进而引起散发多巴胺能神经元退行性病变的发生亟待大量实验的验证和认知。本研究探讨百草枯暴露以及5-氮杂-2-脱氧胞苷(5'-aza-dC)干预后对SH-SY5Y细胞神经毒性的影响。为深入探索PD的发病机制提供新思路。
1 材料与方法
1.1 材料 DMEM,胎牛血清和0.25%胰蛋白酶购自Life公司,Annexin V-FITC/propidium凋亡检测试剂盒以及N′-硝基-L-精氨酸(NO合酶抑制剂)购自罗氏公司。5,6-羧基-2′,7′-二氯荧光黄-二乙酸酯(DCFH-DA,活性氧检测探针),四甲基偶氮噻唑蓝(MTT),5′-aza-dC和百草枯购自Sigma公司。大鼠抗人Bcl-2,Bax和细胞色素C的抗体购自Santa Cruz。兔抗GAPDH抗体自Abcam 公司。辣根过氧化物酶耦联的山羊抗兔IgG以及化学发光的检测系统(ECL检测试剂)均购自Cell Signaling Technology公司。
1.2 细胞培养以及药物处理 人神经胶质瘤SH-SY5Y细胞复苏后,培养在DMEM完全培养液中(含有10%胎牛血清,2 mmol/L L-谷氨酰胺,100 IU/ml青-链霉素),环境为37℃含5%二氧化碳培养箱中。百草枯于DMSO中制备成10 mmol/L的储液(使用时培养液稀释),5′-aza-dC溶解于的磷酸盐缓冲盐中,制成0.5 mmol/L的储液。处理时,这些细胞被随机分为四组:对照组,5′-aza-dC干预组,百草枯组和5′-aza-dC +百草枯组。5′-aza-dC +百草枯组预先用5′-aza-dC干预24 h,然后暴露于百草枯12 h。
1.3 细胞增殖检测 使用MTT比色法对培养细胞的生存率进行了评价。实验前1 d细胞按1×104/孔铺96孔板,按指定时间用化合物处理后,加入5 mg/ml MTT溶液,37℃孵育4 h后在酶标仪上570 nm处读数。以对照组OD570 nm为空白对照,计算细胞生存率。
1.4 细胞凋亡检测 SH-SY5Y细胞分别用5′-aza-dC,百草枯组和5′-aza-dC+百草枯处理24 h,并设置对照组。胰酶消化为单细胞悬液后,PBS漂洗2次,按照试剂盒说明书,加入含Annexin-V和碘化丙啶的试剂,暗处避光常温孵育10~15 min,FACS Calibur流式细胞仪检测荧光强度。激发波长为488 nm,并分析凋亡细胞比例。
1.5 活性氧簇(ROS)检测 暴露于5′-aza-dC或百草枯或5′-aza-dC+百草枯12 h后,使用氧化敏感荧光探针DCFH-DA检测细胞内ROS水平,细胞胰酶消化后处理成单细胞悬液。用无血清培养液稀释DCFH-DA储液,使终浓度为10 μmol/L,37℃孵育30 min,PBS洗涤细胞2次,在488 nm激发,525 nm波长检测荧光水平。
1.6 免疫印迹 细胞取出后弃上清,冷PBS淋洗细胞2遍,RIPA裂解液中冰上裂解细胞5 min,离心后收集蛋白上清在10%的SDS-page胶上电泳。蛋白分离后经恒流1.5 h电转移到PVDF膜后,使用含5%脱脂牛奶的封闭液封闭1 h,与Bcl-2以及Bax等特异性抗体4℃孵育过夜后,次日加入二抗室温孵育1 h,TBST洗膜3次,取出膜后加入ECL发光液,暗室中压片显影。NIH Image J(Version 1.48)软件对样品中各个条带的灰度值进行计算,并用β-actin的灰度值进行均一化处理。
1.7 统计学方法 应用SPSS17.0统计软件进行单因素方差分析及t检验。
2 结 果
2.1 5′-aza-dC干预促进百草枯对细胞活力的影响 MTT法研究显示,当单独使用百草枯处理神经胶质瘤细胞SH-SY5Y 24 h后,当百草枯浓度≤80 μmol/L时,对细胞增殖无显著影响,当浓度>80 μmol/L时,随着剂量的增加对细胞的抑制增殖作用逐渐递增,320 μmol/L时,抑制率达到(60±8)%。5′-aza-dC在0.5 μmol/L时对细胞增殖无显著影响。因此,后续时使用80 μmol/L和5′-aza-dC联合作用。当培养的细胞用5′-aza-dC(0.5 μmol/L)作用24 h,然后暴露在百草枯(80 μmol/L)12 h,存活率明显低于仅暴露于5′-aza-dC干预或百草枯中12 h。见表1。
2.2 5′-aza-dC +百草枯对SH-SY5Y细胞凋亡的影响 通过Annexin V/PI双染结果表明,暴露于5′-aza-dC(0.5 μmol/L)或百草枯(80 μmol/L)不会导致显著细胞凋亡或坏死。然而,当培养的细胞进行预处理以5′-aza-dC(0.5 μmol/L)处理24 h,然后暴露百草枯(100 μmol/L)12 h后,与对照组相比,细胞凋亡比例(44.4±3.2)%是显著大于细胞暴露于5′-aza-dC干预或百草枯单独干预组,见表1。
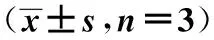
表1 不同实验组处理后SH-SY5Y细胞各项指标测定结果
与对照组比较:1)P<0.05
2.3 5′-aza-dC+百草枯对SH-SY5Y细胞活性氧的影响 SH-SY5Y细胞经过不同组处理后,流式细胞仪显示,在对照组中平均DCFH-DA荧光强度为(550±40)。在5′-aza-dC干预(0.5 μmol/L)和百草枯(80 μmol/L)组,DCF平均荧光强度分别为(520±33)和(620±29)。ROS水平与对照组比较无统计学差异(P>0.05)。而5′-aza-dC+百草枯组中,平均的DCF荧光强度为(888±45),提示ROS水平显著高于对照组及单用5′-aza-dC或百草枯干预组,表明5′-aza-dC处理后,细胞对百草枯的氧化应激水平提高,这可能与细胞的凋亡增加密切相关。
2.4 SH-SY5Y细胞中凋亡线粒体途径中相关蛋白表达情况 与对照组相比,Bax蛋白和细胞色素C的水平在联合处理组中水平要高于单独处理的水平。而Bcl-2蛋白的表达则相反,在联合组中要低于单独处理组,通过对免疫印迹条带进行光密度分析、定量和均一化(与内参GAPDH比较)后发现(图1),联合组中Bax和细胞色素C的表达水平与对照组比较,差异有统计学意义(P<0.05)。Bax和细胞色素C的体现在5′-aza-dC干预或百草枯单独组没有变化。然而5′-aza-dC+百草枯组中,表达Bcl-2降低,Bax蛋白表达增加,Bcl-2/Bax的速度下降,因而细胞色素C表达增加。见表1。

1~4:对照组、5-aza-dC组、百草枯组、5′-aza-dC+百草枯组图1 5′-aza-dC及百草枯处理SY-SH5Y细胞后 对线粒体凋亡途径中相关蛋白表达的影响
3 讨 论
暴露于环境毒素和遗传修饰被认为可能参与PD发病中〔5〕。本研究的结果显示,在正常对细胞增殖无显著影响的浓度下,当百草枯和甲基转移酶抑制剂5′-aza-dC联合作用后,可以引起细胞增殖的抑制,进而对神经细胞有比较大的损伤,这也说明DNA甲基化程度的削弱,可能导致PD发生的门槛降低,从而提高PD发生的风险。
百草枯作为常用用于控制杂草生长的除草剂,过去的国内外研究显示,百草枯不仅可以严重损伤多巴胺能神经元,而且与PD风险增加有关〔6,7〕。本研究结果也证实,当百草枯的浓度大于80 μmol/L时候,抑制细胞增殖效应显著增加,而低浓度的百草枯对细胞增殖的影响不显著,一般情况下,就算长期接触百草枯,体内血液中的浓度很少会达到这个水平。说明百草枯若要对人体神经细胞发挥显著影响,存在某个特定条件。5′-aza-dC作为常用的甲基转移酶抑制剂可以引起基因组DNA全局化的甲基化抑制,从而使得不少基因上游启动子的甲基化水平降低,某些基因表达增加〔8〕。本研究中发现,当5′-aza-dC预先处理后,可以显著提高百草枯对神经胶质瘤细胞的毒性,抑制其增殖。进一步通过凋亡分析显示,5′-aza-dC+百草枯后细胞凋亡也增加。PD的神经变性过程往往伴随细胞氧化应激水平的提高〔9〕。本文结果显示,通过削弱基因组的甲基化程度,使得环境毒物百草枯对细胞的毒性增加,其作用可能是通过提高ROS水平,进而引起细胞凋亡而引起,推及到体内,以前的研究表明,随着人体老化的发生,一些神经元的保护基因的功能可以因表遗传学改变而受损〔10〕,环境毒物及重金属等的暴露也可以改变基因表达,以上二者交互作用后引起迟发性的神经退行性疾病发生。
有充分的证据显示,线粒体凋亡信号途径在PD的发病机制中起重要作用〔11〕。DNA甲基化也已牵涉许多宿主细胞的很多信号通路,尤其是神经系统〔12,13〕,包括线粒体凋亡的信号转导。Bcl-2蛋白家族通过调控线粒体功能中起着凋亡信号转导中起重要作用。Bax和Bcl-2是主要的凋亡蛋白,并分别作为凋亡诱导剂和抑制剂,Bax蛋白也被证明在诱导细胞凋亡响应于应激刺激发挥至关重要的作用,Bax蛋白的很大一部分是位于正常细胞的细胞质中,在氧化应激压力下,Bax蛋白重新分布到线粒体,易位到线粒体后,Bax蛋白可以诱导细胞色素C释放,外线粒体细胞膜上形成孔道,导致细胞色素C的释放,这也是诱导caspase-3活化的充分条件,是神经细胞凋亡内源性途径的标志性事件〔14〕。很少有研究着重对在百草枯所诱发线粒体凋亡信号通路进行研究,我们的研究表明,5′-aza-dC+百草枯暴露后抗凋亡关键蛋白(Bcl-2蛋白)的表达显著下降,增加Bax蛋白的表达,并刺激了细胞色素C从线粒体释放,从而最终促进线粒体介导的细胞凋亡。
综上所述,DNA甲基化修饰减少,使得PD发生的门槛降低,类似百草枯这样的环境毒物可以对细胞的损伤增加,抑制细胞增殖,诱导凋亡发生,并使得多巴胺能神经元氧化应激水平增加,并可以引起线粒体途径的活化,启动凋亡内源性途径,总的效果是引起神经元细胞的损伤和死亡,这些或许可以用于解释长期暴露百草枯类除草剂后,可以诱导老年人发生PD。后续的研究将着重探讨体内模型中,在暴露百草枯及影响甲基化水平后对PD发生和进展的影响。
1 Taetzsch T,Block ML.Pesticides,microglial NOX2,and Parkinson′s disease〔J〕.J Biochem Mol Toxicol,2013;27(2):137-49.
2 Tsuboi Y.Environmental-genetic interactions in the pathogenesis of Parkinson′s disease〔J〕.Exp Neurobiol,2012;21(3):123-8.
3 封 华,刘承伟.遗传基因与帕金森病相关性的研究进展〔J〕.中国老年学杂志,2011;31(10):1911-3.
4 尹献辉,刘 佳,李雪芹,等.衰老的表观遗传机制研究进展〔J〕.中国细胞生物学学报,2013;35(7):1027-34.
5 吴远双,孟庆雄.神经退行性疾病的表观遗传学〔J〕.生命科学研究,2013;17(4):370-6.
6 Kamel F.Epidemiology.Paths from pesticides to Parkinson′s〔J〕.Science,2013;341(6147):722-3.
7 Goldman SM,Kamel F,Ross GW,etal.Genetic modification of the association of paraquat and Parkinson′s disease〔J〕.Mov Disord,2012;27(13):1652-8.
8 苏 玉,王 溪,朱卫国.DNA甲基转移酶的表达调控及主要生物学功能〔J〕.遗传,2009;31(11):1087-93.
9 袁 红,梁立武,刘 平,等.氧化应激与帕金森病多巴胺神经元死亡〔J〕.武警医学,2007;18(6):448-50.
10 Kwok JB.Role of epigenetics in Alzheimer′s and Parkinson′s disease〔J〕.Epigenomics,2010;2(5):671-82.
11 Berry C,La Vecchia C,Nicotera P.Paraquat and Parkinson′s disease〔J〕.Cell Death Differ,2010;17(7):1115-25.
12 Ravi B,Kannan M.Epigenetics in the nervous system:an overview of its essential role〔J〕.Indian J Hum Genet,2013;19(4):384-91.
13 Rudenko A,Tsai LH.Epigenetic modifications in the nervous system and their impact upon cognitive impairments〔J〕.Neuropharmacology,2014;80:70-82.
14 Levy OA,Malagelada C,Greene LA.Cell death pathways in Parkinson′s disease:proximal triggers,distal effectors,and final steps〔J〕.Apoptosis,2009;14(4):478-500.
〔2015-02-23修回〕
(编辑 苑云杰/曹梦园)
国家社科基金青年项目(No.15CGL064);甘肃政法学院科研青年项目(No.GZF2d3XQNLW015)
王 炜(1982-),男,理学硕士,讲师,主要从事药理学、毒理学及毒物分析等相关领域研究。
R992
A
1005-9202(2017)08-1878-03;
10.3969/j.issn.1005-9202.2017.08.024
1 甘肃政法学院甘肃省证据科学技术研究与应用重点实验室
2 中国人民解放军兰州军区兰州总医院呼吸内科
