Recombinant human fi broblast growth factor-2 promotes nerve regeneration and functional recovery after mental nerve crush injury
2017-05-03sunghoLeeweipengJinNariseoKangMipangBongjuKimsoungMinKimJonghoLee
sung ho Lee, wei-peng Jin, Na ri seo, Kang-Mi pang, Bongju Kim, soung-Min Kim, Jong-ho Lee,
1 Dental Research Institute, Department of Oral and Maxillofacial Surgery, Seoul National University School of Dentistry, Seoul, Korea
2 Dental Life Science Research Institute, Clinical Translational Research Center for Dental Science, Seoul National University Dental Hospital, Seoul, Korea
3 Dental Research Institute, Seoul National University, Seoul, Korea
Recombinant human fi broblast growth factor-2 promotes nerve regeneration and functional recovery after mental nerve crush injury
sung ho Lee1, wei-peng Jin1, Na ri seo1, Kang-Mi pang1, Bongju Kim2, soung-Min Kim1, Jong-ho Lee1,3,*
1 Dental Research Institute, Department of Oral and Maxillofacial Surgery, Seoul National University School of Dentistry, Seoul, Korea
2 Dental Life Science Research Institute, Clinical Translational Research Center for Dental Science, Seoul National University Dental Hospital, Seoul, Korea
3 Dental Research Institute, Seoul National University, Seoul, Korea
How to cite this article:Lee SH, Jin WP, Seo NR, Pang KM, Kim B, Kim SM, Lee JH (2017) Recombinant human fbroblast growth factor-2 promotes nerve regeneration and functional recovery after mental nerve crush injury. Neural Regen Res 12(4):629-636.
Open access statement:Tis is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under the identical terms.
Funding:Tis study was supported by a grant from the Korea Healthcare Technology R&D Project, Ministry for Health, Welfare & Family Affairs, Republic of Korea, No. A101578.
Several studies have shown that fi broblast growth factor-2 (FGF2) can directly af f ect axon regeneration aer peripheral nerve damage. In this study, we performed sensory tests and histological analyses to study the ef f ect of recombinant human FGF-2 (rhFGF2) treatment on damaged mental nerves.e mental nerves of 6-week-old male Sprague-Dawley rats were crush-injured for 1 minute and then treated with 10 or 50 μg/mL rhFGF2 or PBS in crush injury area with a mini Osmotic pump. Sensory test using von Frey fi laments at 1 week revealed the presence of sensory degeneration based on decreased gap score and increased difference score. However, at 2 weeks, the gap score and difference score were signif i cantly rebounded in the mental nerve crush group treated with 10 μg/mL rhFGF2. Interestingly, treatment with 10 μg/mL rhFGF had a more obviously positive ef f ect on the gap score than treatment with 50 μg/mL rhFGF2. In addition, retrograde neuronal tracing with Dil revealed a signif i cant increase in nerve regeneration in the trigeminal ganglion at 2 and 4 weeks in the rhFGF2 groups (10 μg/mL and 50 μg/mL) than in the PBS group.e 10 μg/mL rhFGF2 group also showed an obviously robust regeneration in axon density in the mental nerve at 4 weeks. Our results demonstrate that 10 μg/mL rhFGF induces mental nerve regeneration and sensory recovery aer mental nerve crush injury.
nerve regeneration; mental nerve; fbroblast growth factor; crush injury; sensory neuron; functional recovery; neural regeneration
Introduction
Damage to the inferior alveolar nerve often occurs during surgery or during other types of damage (Savignat et al., 2007; Sun et al., 2009; Kemp et al., 2011). For example, the inferior alveolar nerve (IVN) and mental nerve (MN) can be easily damaged during dental surgery, including dentoalveolar, tumor, and orthognathic surgeries (Savignat et al., 2008). Damage to the IVN can also occur during dental implantation (Alhassani and AlGhamdi, 2010).e MN originates from the trigeminal ganglion and innervates the lower lip (Imai et al., 2003). Several surgical procedures are used to treat peripheral nerve damage, including sutures, nerve graing, electrical stimulation, and stem cell therapy; however, these treatments are insufficient in recovering nerve function (Sullivan et al., 2016).
For nerve regeneration, neurotrophic factors such as nerve growth factor (NGF), fi broblast growth factor (FGF), and brain-derived neurotrophic factor (BDNF) are ef f ective treatments (Kerekes et al., 1997; Choi et al., 2008). These neurotrophic factors control the differentiation and proliferation of adult neuronal progenitor cells (Kuhn et al., 1997; Palmer et al., 1999; Binder and Scharfman, 2004; Werner et al., 2011; Li et al., 2012). FGF determines the size of the cerebral cortex during embryogenesis and reduces the glutamatergic pyramidal neurons in frontal and temporal cortex (Vaccarino et al., 1999; Shin et al., 2004; Rash et al., 2011). FGF also promotes the proliferation and differentiation of neurons, and regulates the repair of injured cervical root nerves (Li et al., 2012).
Basic fibroblast growth factor (bFGF) has been shown to promote regeneration of damaged or injured peripheral nerves (Fujimoto et al., 1997). Fujimoto et al. (1997) demonstrated that FGF directly af f ected axon regeneration aer peripheral nerve damage. Grothe and Nikkhah (2001) demonstrated that exogenously applied bFGF mediates rescue ef f ects on injured sensory neurons and supports neurite outgrowth of transectioned nerves.
Materials and Methods
Animals
Surgical procedures
Rats were anesthetized with an intraperitoneal injection of a 4:1 mixture of hetamine hydrochloride (100 mg/kg, Ketara®, Yuhan, Korea) and xylazine hydrochloride (25 mg/kg, Rumpun®, Bayer, Korea). A 4-mm crush injury was induced by exposing the left MN and tightening the area 5 mm away from the mental foramen with the second clip of HEMOSTATS forceps (Baby-mixter, No. 13013-14, Canada) for 1 minute. Tagging with the ETHILONS Suture (ETHICON Inc., Somerville, NJ, USA) on the distal of the injured area makes reference to the harvesting of the MN crush injury (Figure 1A–C). An osmotic pump (1004 ALZET®mini-Osmotic Pump, CA, USA) was inserted into the neck back. A Rat Jugular Catheter (ALZET®, CA, USA) was used to connect the osmotic pump and the MN (Figure 1D, E).
rhFGF-2 treatment and experimental design
rhFGF-2 (GENOSS Co., Ltd, Suwon, Republic of Korea) was injected at concentrations of 0.1 μg/mL (kg/d), 0.2 μg/mL (kg/d), and 0.4 μg/mL (kg/d) into the osmotic pumps for 1, 2, and 3 weeks in a preliminary experiment (n= 6 in each group at each time point, totally 54 rats). Afterwards, the inserted osmotic pumps in animals were used for administration with rhFGF-2 either at low dose (10 μg/mL (kg/d)) or at high dose (50 μg/mL (kg/d)) (n= 8 in each group at each time point, totally 64 rats). In the PBS and rhFGF groups, 84 μL of PBS or rhFGF was injectedviathe osmotic pump. In this manner, PBS or rhFGF was administered at 0.5 μL/h for 1, 2, 3, and 4 weeks (Lin et al., 2011).e osmotic pumps were refreshed once a week.
Sensory test
According to the method from Seinoa et al. (2009), a sensory test was performed to compare the touch sensitivity to mechanical stimulation using von Frey filaments (Semmes-Weinstein Monofilaments, North Coast Medical, Inc., Arcata, CA, USA).e rapid movement of the front legs of a rat towards its head or face due to the bending force of a von Frey fi lament (bending force of 0.008, 0.02, 0.04, 0.07, 0.016, 0.4, 0.6, 1.0, 1.4, 2.0, 4.0, and 6.0 g) indicated a positive response to stimulation (Figure 2A). The touch threshold to mechanical stimulation (in g) of the ipsilateral and contralateral sides was calculated by subtracting the value of the contralateral mental nerve area from the value of the ipsilateral mental nerve area.e gap score was def i ned as the difference between the medial and distal sensitivity of the MN to mechanical stimulation (Figure 2B), and the difference score was calculated by subtracting the value of the contralateral lip area from the value of the ipsilateral lip area (Figure 2B) (Seino et al., 2009). Decreased gap score and increased difference score mean the presence of sensory degeneration.e sensory test was performed before and at 1, 2, 3, and 4 weeks aer surgery in each group.
Retrograde nerve labeling
Retrograde labeling and counting of back-labeled sensory neurons were performed as described before (Li et al., 2012).e MN crush injury site was retrograde labeled with 1,1′-dioctadecyl-3,3,3′3′-tetramethylindocarbocyanine perchlorate (Dil, C59H97ClN2O4, Molecular Probes; Eugene, OR, USA) to measure nerve regeneration 2 and 4 weeks aer the MN crush injury (n= 8 in each group at each time point). Five days after the Dil injection in mental nerve, the rats were rapidly anesthetized using an intraperitoneal (IP) injection of chloropent (3 mL/kg), and transcardially perfused using a 0.9% saline solution with 1% heparin (JW Pharmaceutical, Seoul, Korea) followed by 4% paraformaldehyde (Merck, Darmstadt, Germany) in 0.1 M sodium phosphate buffer (pH7.2). Immediately after fixation, the trigeminal ganglia were extracted and post-f i xed with the same fi xative at 4°C for 24 hours. To prevent the occurrence of freezing artifacts during the preparation of frozen tissue sections, the tissue was fi xed in 30% sucrose (Sigma, St, Louis, MO, USA) at 4°C for 24 hours.e tissue sections were observed using a confocal laser-scanning microscope (CLSM, LSM700, Carl Zeiss, Oberkochen, Germany).
Histomorphometric evaluation
In the preliminiary experiment, animals were sacrificed at 1, 2 and 3 weeks (n= 6 in each group at each time point) and in the formal experiment, animals in each group were sacrif i ced at 2 and 4 weeks (n= 8 in each group at each time point). Once rats were euthanized, a 10-mm section of the MN including the MN crush injury site was incised and fi xed in PBS (pH7.4) and 2.5% glutaraldehyde. Next, nerve sample was transversely cut at the center, and 2% osmium tetroxide solution was added. Subsequently, the nerve tissue sample was embedded with Epon 812 (Nisshin EM, Tokyo, Japan), cut into 1 μm-thick sections, and stained with 1% toluidine blue. Images were captured using a SPOT RTTM-KE color mosaic,and digitized using SPOT software version 4.6 (Diagnostic Instruments, Inc., Sterling Heights, MI, USA). To simplify axon counting (total axon number), the total cross-section area of the nerve (total axon area) was measured at 40× magnif i cation and 3 sampling fi elds were randomly selected at 200× magnif i cation using a previously reported protocol (Li et al., 2012).e mean axon density was calculated by dividing the total number of nerve fi bers within the sampling field by its area (N/mm2). The total axon number (N) wasestimated by multiplying the mean fi ber density by the entire cross-section area of the whole nerve, assuming a uniform distribution of nerve fi bers across the entire section (Alrashdan et al., 2011).
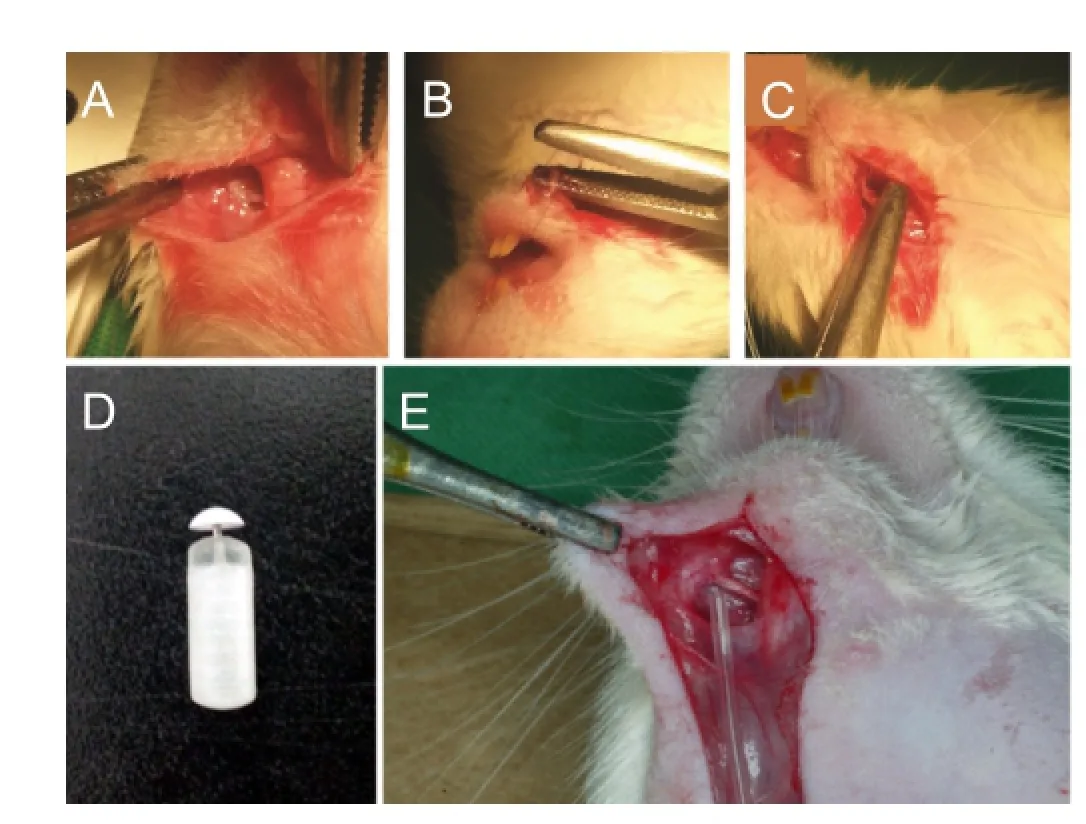
Figure 1 Animal surgery methods and materials.

Figure 2 Calculation method of gap score and diference score using von Frey fi laments.
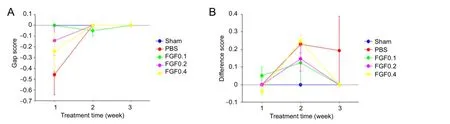
Figure 3 Preliminary test of the sensory functional recovery of rats with mental nerve crush injury assessed by von Frey fi laments test aer recombinant human fi broblast growth factor-2 (rhFGF2) treatment.

Figure 4 Efect of recombinant human fi broblast growth factor-2 (rhFGF2) treatment on touch sensory recovery of rats with mental nerve crush injury.
To calculate myelin thickness, frozen sections were cut using the cryostat (Leica, Ultracut, UCT, Vienna, Austria) and double-stained with hematoxylin (Sigma, St. Louis, MO, USA) and eosin (Sigma). Then the sections were analyzed using OPTIMAS Ver. 6.5 (Media Cybernetics, Bethesda, MD, USA) to measure myelin thickness. G-ratio was def i ned as the ratio of axon diameter to the myelinated fi ber diameter.e size of soma was measured as described before (Li et al., 2012). All measurements and histological observations were performed by individuals who were not involved in collecting samples for the experiment.
Statistical analysis
All variables were expressed as the mean ± SD. Data analysis was conducted using one-way analysis of variance with STATVIEW software (Abacus, Berkeley, CA, USA). The Mann-WhitneyUtest for nonparametric analysis was used. A level ofP< 0.05 was considered statistically signif i cant.
Results
Sensory function
In a preliminary test, sensory nerve impairment and functional recovery were not observed in injured mental nerve aer rhFGF2 treatment (Figure 3).
A decline in sensory function was indicated by the gap score in the rhFGF groups at 1 week aer the crush injury. However, a sensory functional recovery was observed at 2–4 weeks (Figure 4A;P< 0.05). From the viewpoint of difference score, sensory function was improved at 1 week but it maintained a stable level from week 2 from which the nerve injury began to recover as indicated by the gap score. However, sensory function was signif i cantly different at 3 and 4 weeks between 10 μg/mL rhFGF2, 50 μg/mL rhFGF2 and PBS groups (Figure 4B;P< 0.05).
Nerve regeneration in the trigeminal ganglia of rats treated with rhFGF2 aer mental nerve crush injury
Histomorphometric evaluation results
Although axonal changes were dif fi cult to be observed (Figure 6A), rhFGF2 treatments did not induce rebounding of total axonal number (Figure 6B). Comparisons of axon density and total area between groups showed that rhFGF2 treatment did not lead to recovery from nerve injury (Figure 6C, D).
No obvious axonal changes were observed in the PBS group, but total axon numbers increased in the FGF10 and FGF50 groups (Figure 7A). The total axon numbers were signif i cantly different between the PBS, FGF10, and FGF50 groups at 2 and 4 weeks (Figure 7B;P< 0.001).ere was statistical signif i cance in total axon number between FGF10 and FGF50 groups and PBS group at 2 and 4 weeks (Figure 7B;P< 0.001). Total axon area increased with rhFGF2 treatment and there was significant difference between FGF10 and FGF50 groups at 2 and 4 weeks (Figure 7C;P< 0.001). Axon density (total axon numbers/total axon area) was calculated to verify the increase in axons per unit area. Axon density was significantly different in both the FGF10 and FGF50 groups compared to the PBS group at 2 and 4 weeks (Figure 7D;P< 0.001), and axon density increased in the FGF10 and FGF50 groups than in the PBS group at 2 and 4 weeks (Figure 7D;P< 0.001).
According to a previous report by our laboratory (Li et al., 2012), the thickness of myelin sheath was measured in each group at 4 weeks (Figure 8A–D). For measurement of myelin sheath thickness, axon surrounded by uniform-thickness myelin layer was chosen. Mean myelin thickness and G-ratio in the FGF10 and FGF50 groups were signif i cantly increased than in the PBS group (Figure 8E and F,P< 0.001).
Discussion
FGF2 triggers mitogenesis and the proliferation of neural stem cellsin vitro(Vescovi et al., 1993; Palmer et al., 1999). FGF2 also plays important physiological rolesin vivo, including neurogenesis that occurs in the subventricular zone and dentate gyrus of the hippocampus (Vescovi et al., 1993; Craig et al., 1996; Kuhn et al., 1997; Nakagami et al., 1997; Palmer et al., 1999; Yoshimura et al., 2001). Lin et al. (2011) found that FGF2 administration reduced allodynia, and Collombet et al. (2011) reported that cytokine treatment using FGF2 or epidermal growth factor is ef f ective for nerve regeneration and cognitive behavior recovery. To induce long distance regeneration, leupeptin promotes FGF2-induced axonal growth (Hausott et al., 2008), and inhibition of FGFR1 endocytosis stimulates the axonal branching of sensory neurons (Hausott et al., 2011).
Here, we provide evidence that administration of rhFGF2 can be used to treat MN crush injury in rats. Typically, for nerve injuries, neurotrophic factors are primarily administered subcutaneously, intramuscularly or intravenouslyviaan osmotic pump or a Hamilton syringe. In the current study, we directly and constitutively administered rhFGF2 to the crush injury area using an osmotic pump.
The purpose of our study was to investigate the possible recovery of sensory neurons by administering rhFGF2 to the nerve injury area. Because the rhFGF2 that we used had not been used previously in clinical or pre-clinical tests, a preliminary experiment was performed using low doses of rhFGF2 to determine a suitable dose for sensory recovery. However, these low doses were not associated with functional recoveryin the sensory neurons.
Based on previous studies (Hausott et al., 2008; Collombet et al., 2011; Lin et al., 2011; Li et al., 2012), we performed rhFGF2 treatments using 2 dosage groups: a low dose group that was treated with 10 μg/mL rhFGF2 (FGF10) and a high dose group that was treated with 50 μg/mL rhFGF2 (FGF50). We investigated the functional recovery of sensory neurons aer rhFGF2 treatments in rats with a MN crush injury.
In the sensory test that we used based on the method from Seino et al. (2009), the subjective will of an examiner can affect the experimental results. To avoid this issue, the examiner was blinded to the experimental groups. We recorded the absolute values of the results from the sensory test for each rat to reveal the ef f ects of the experimental conditions (Chaplan et al., 1994). However, the inf l uence of the experimental conditions of the rats was minimized by measuring each score not as an absolute value but as the mean value for each group. Raising a front leg to touch the filament indicated a ‘positive’ touch response because the lower lip of the rat was pierced so that the animal could not turn its head and bite the fi lament with its teeth.e sensory test was performed once 3 days before the surgery and once a week from weeks 1 to 4 aer the surgery.e sensory test score was expressed as a gap score and a difference score. Although sensory loss based on a comparison of the preand post-surgery gap score at week 1 occurred in all groups except the sham group, sensory recovery indicated by a gap score closer to 0 was observed in the rhFGF2 group at 2 weeks. In contrast, the difference score increased in the rhFGF2 group at 1 week and began to decrease at 2 weeks.e symmetric graphs of the difference score and gap score show that the extent of the recovery in sensory function progressed over time in the rhFGF2-treated groups.
Nerve regeneration of the trigeminal ganglia decreased over time in the PBS group. Compared to the PBS group, the rhFGF2 group had significantly more nerve regeneration over time. Axonal density and total axon number were signif i cantly greater in the rhFGF2 groups than in the PBS group. Moreover, over time, the total axon number, total axon area, and axon density increased in the rhFGF2 groups.
There are many studies reporting that stem cells can be used to repair peripheral nerve injury. But there are few clinical studies about it. There is evidence that at 8 weeks after transplantation of human hair follicle stem cells for repair of sciatic nerve injury, the growth of spindle cells was observed (Amoh et al., 2005, 2008, 2009, 2012; Mii et al., 2013; Hof f man, 2015). Li et al. (2012) recombinant human nerve growth factor-β gene transfer promotes regeneration of crush-injured mental nerve in rats. As in the abovementioned experiments, stem cells or stem cell growth factors have been reported to repair peripheral nerve damage. However, we evaluated neurological recovery after intravenous injection of growth factor for mental nerve crush injury.
Our results suggest that rhFGF2 treatment affects the recovery of sensory function from a MN crush injury. However, the rhFGF2 used in our study had not been tested clinically; therefore, a more accurate dose calculation is required for the clinical application of rhFGF2. In addition, future studies should investigate the mechanism underlying down-regulation of FGF activity and the mechanism by which rhFGF2 directly affects nerve regeneration and the functional recovery of sensory neurons.
Author contributions:SHL, SMK, and JHL had full access to all data and participated in data integrity and data accuracy analysis. WPJ, NRS, KMP, and BK participated in data collection. SHL, SMK, and JHL participated in study design, study supervision, and manuscript instruction. All authors approved the fnal version of this paper.
Conficts of interest:None declared.
Plagiarism check:This paper was screened twice using CrossCheck to verify originality before publication.
Peer review:Tis paper was double-blinded and stringently reviewed by international expert reviewers.
Alhassani AA, AlGhamdi AST (2010) Inferior alveolar nerve injury in implant dentistry: diagnosis, causes, prevention, and management. J Oral Implantol 36:401-407.
Alrashdan MS, Sung MA, Kwon YK, Chung HJ, Kim SJ, Lee JH (2011) Ef f ects of combining electrical stimulation with BDNF gene transfer on the regeneration of crushed rat sciatic nerve. Acta Neurochir (Wien) 153:2021-2029.
Amoh Y, Li L, Katsuoka K, Hof f man RM (2008) Multipotent hair follicle stem cells promote repair of spinal cord injury and recovery of walking function. Cell Cycle 7:1865-1869.
Amoh Y, Li L, Campillo R, Kawahara K, Katsuoka K, Penman S, Hof fman RM (2005) Implanted hair follicle stem cells form Schwann cells that support repair of severed peripheral nerves. Proc Natl Acad Sci U S A 102:17734-17738.
Amoh Y, Kanoh M, Niiyama S, Hamada Y, Kawahara K, Sato Y, Hof fman RM, Katsuoka K (2009) Human hair follicle pluripotent stem (hfPS) cells promote regeneration of peripheral-nerve injury: An advantageous alternative to ES and iPS cells. J Cell Biochem 107:1016-1020.
Amoh Y, Aki R, Hamada Y, Niiyama S, Eshima K, Kawahara K, Sato Y, Tani Y, Hof f man RM, Katsuoka K (2012) Nestin-positive hair follicle pluripotent stem cells can promote regeneration of impinged peripheral nerve injury. J Dermatol 39:33-38.
Binder DK, Scharfman HE (2004) Mini review. Growth factors 22:123-131.
Chaplan SR, Bach F, Pogrel J, Chung J, Yaksh T (1994) Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Methods 53:55-63.
Choi KC, Yoo DS, Cho KS, Huh PW, Kim DS, Park CK (2008) Ef f ect of single growth factor and growth factor combinations on differentiation of neural stem cells. J Korean Neurosurg Soc 44:375-381.
Collombet JM, Béracochéa D, Liscia P, Piérard C, Lallement G, Filliat P (2011) Long-term ef f ects of cytokine treatment on cognitive behavioral recovery and neuronal regeneration in soman-poisoned mice. Behav Brain Res 221:261-270.
Craig CG, Tropepe V, Morshead CM, Reynolds BA, Weiss S, Van der Kooy D (1996) In vivo growth factor expansion of endogenous subependymal neural precursor cell populations in the adult mouse brain. J Neurosci 16:2649-2658.
Fujimoto E, Mizoguchi A, Hanada K, Yahima M, Ide C (1997) Basic fibroblast growth factor promotes extension of regenerating axons of peripheral nerve. In vivo experiments using a Schwann cell basal lamina tube model. J Neurocytol 26:511-528.
Hausott B, Schlick B, Vallant N, Dorn R, Klimaschewski L (2008) Promotion of neurite outgrowth by fi broblast growth factor receptor 1 overexpression and lysosomal inhibition of receptor degradation in pheochromocytoma cells and adult sensory neurons. Neuroscience 153:461-473.
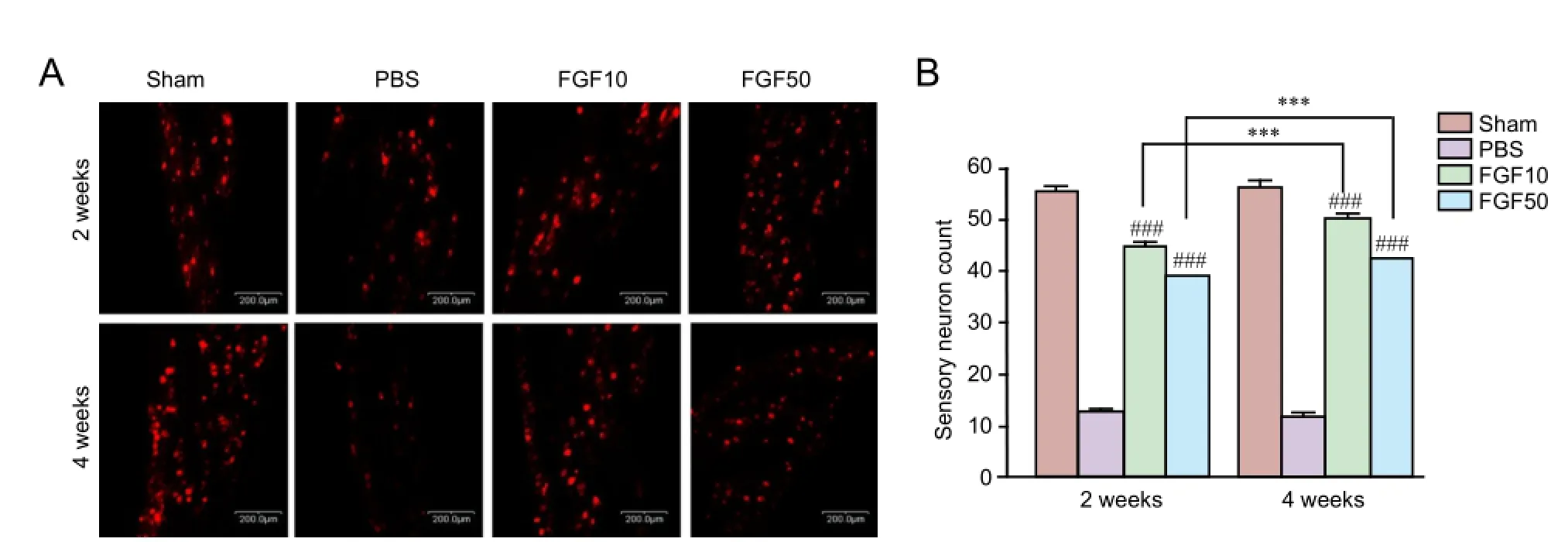
Figure 5 Retrograde tracing of the trigeminal ganglion with Dil.


Figure 7 Axon observation in injured mental nerve aer recombinant human fi broblast growth factor-2 (rhFGF2) treatment.

Figure 8 Changes in myelin sheath thickness in injured mental nerve.
Hausott B, Rietzler A, Vallant N, Auer M, Haller I, Perkhofer S, Klimaschewski L (2011) Inhibition of fi broblast growth factor receptor 1 endocytosis promotes axonal branching of adult sensory neurons. Neuroscience 188:13-22.
Hof f man RM (2015) Nestin-expressing hair follicle-accessible pluripotent stem cells for nerve and spinal cord repair. Cells Tissues Organs 200:42-47.
Imai T, Atsumi Y, Matsumoto K, Yura Y, Wakisaka S (2003) Regeneration of periodontal Ruf fini endings of rat lower incisors following nerve cross-anastomosis with mental nerve. Brain Res 992:20-29.
Kemp SW, Webb AA, Dhaliwal S, Syed S, Walsh SK, Midha R (2011) Dose and duration of nerve growth factor (NGF) administration determine the extent of behavioral recovery following peripheral nerve injury in the rat. Exp Neurol 229:460-470.
Kerekes N, Landry M, Rydh-Rinder M, H?kfelt T (1997)e ef f ect of NGF, BDNF and bFGF on expression of galanin in cultured rat dorsal root ganglia. Brain Res 754:131-141.
Kuhn HG, Winkler J, Kempermann G,al LJ, Gage FH (1997) Epidermal growth factor and fi broblast growth factor-2 have different effects on neural progenitors in the adult rat brain. J Neurosci 17:5820-5829.
Li BH, Kim SM, Yoo SB, Kim MJ, Jahng JW, Lee JH (2012) Recombinant human nerve growth factor (rhNGF-β) gene transfer promotes regeneration of crush-injured mental nerve in rats. Oral Surg Oral Med Oral Pathol Oral Radiol 113:e26-34.
Lin YL, Kuo HS, Lo MJ, Tsai MJ, Lee MJ, Huang WC, Kuo WC, Shih YH, Cheng H, Huang MC (2011) Treatment with nerve gras and aFGF attenuates allodynia caused by cervical root transection injuries. Restor Neurol Neurosci 29:265-274.
Mii S, Duong J, Tome Y, Uchugonova A, Liu F, Amoh Y, Saito N, Katsuoka K, Hof f man RM (2013)e role of hair follicle nestin-expressing stem cells during whisker sensory-nerve growth in long-term 3D culture. J Cell Biochem 114:1674-1684.
Nakagami Y, Saito H, Matsuki N (1997) Basic fi broblast growth factor and brain-derived neurotrophic factor promote survival and neuronal circuit formation in organotypic hippocampal culture. Jpn J Pharmacol 75:319-326.
Palmer TD, Markakis EA, Willhoite AR, Safar F, Gage FH (1999) Fibroblast growth factor-2 activates a latent neurogenic program in neural stem cells from diverse regions of the adult CNS. J Neurosci 19:8487-8497.
Rash BG, Lim HD, Breunig JJ, Vaccarino FM (2011) FGF signaling expands embryonic cortical surface area by regulating Notch-dependent neurogenesis. J Neurosci 31:15604-15617.
Savignat M, De-Doncker L, Vodouhe C, Garza J, Lavalle P, Libersa P (2007) Rat nerve regeneration with the use of a polymeric membrane loaded with NGF. J Dent Res 86:1051-1056.
Savignat M, Vodouhe C, Ackermann A, Haikel Y, Lavalle P, Libersa P (2008) Evaluation of early nerve regeneration using a polymeric membrane functionalized with nerve growth factor (NGF) after a crush lesion of the rat mental nerve. J Oral Maxillofac Surg 66:711-717.
Seino H, Seo K, Maeda T, Someya G (2009) Behavioural and histological observations of sensory impairment caused by tight ligation of the trigeminal nerve in mice. J Neurosci Methods 181:67-72.
Shin DM, Korada S, Raballo R, Shashikant CS, Simeone A, Taylor JR, Vaccarino F (2004) Loss of glutamatergic pyramidal neurons in frontal and temporal cortex resulting from attenuation of FGFR1 signaling is associated with spontaneous hyperactivity in mice. J Neurosci 24:2247-2258.
Sullivan R, Dailey T, Duncan K, Abel N, Borlongan CV (2016) Peripheral Nerve Injury: Stem Cellerapy and Peripheral Nerve Transfer. Int J Mol Sci 17:2101.
Sun W, Sun C, Lin H, Zhao H, Wang J, Ma H, Chen B, Xiao Z, Dai J (2009) The effect of collagen-binding NGF-β on the promotion of sciatic nerve regeneration in a rat sciatic nerve crush injury model. Biomaterials 30:4649-4656.
Vaccarino FM, Schwartz ML, Raballo R, Nilsen J, Rhee J, Zhou M, Doetschman T, Cof fi n JD, Wyland JJ, Hung Y-TE (1999) Changes in cerebral cortex size are governed by fi broblast growth factor during embryogenesis. Nat Neurosci 2:246-253.
Vescovi AL, Reynolds BA, Fraser DD, Weiss S (1993) bFGF regulates the proliferative fate of unipotent (neuronal) and bipotent (neuronal/ astroglial) EGF-generated CNS progenitor cells. Neuron 11:951-966.
Werner S, Unsicker K, von Bohlen und Halbach O (2011) Fibroblast growth factor-2 deficiency causes defects in adult hippocampal neurogenesis, which are not rescued by exogenous fi broblast growth factor-2. J Neurosci Res 89:1605-1617.
Yoshimura S, Takagi Y, Harada J, Teramoto T,omas SS, Waeber C, Bakowska JC, Breakefield XO, Moskowitz MA (2001) FGF-2 regulation of neurogenesis in adult hippocampus aer brain injury. Proc Natl Acad Sci U S A 98:5874-5879.
Copyedited by Li CH, Song LP, Zhao M
*< class="emphasis_italic">Correspondence to: Jong-Ho Lee, D.D.S., M.S.D., Ph.D., leejongh@snu.ac.kr.
Jong-Ho Lee, D.D.S., M.S.D., Ph.D., leejongh@snu.ac.kr.
orcid: 0000-0002-8843-545X (Jong-Ho Lee)
10.4103/1673-5374.205104
Accepted: 2017-03-17
杂志排行

中国神经再生研究(英文版)的其它文章
- Recovery of multiply injured ascending reticular activating systems in a stroke patient
- Neuroprotective mechanism of Kai Xin San: upregulation of hippocampal insulin-degrading enzyme protein expression and acceleration of amyloid-beta degradation
- Mitomycin C induces apoptosis in human epidural scar fi broblasts after surgical decompression for spinal cord injury
- Exenatide promotes regeneration of injured rat sciatic nerve
- Ca2+involvement in activation of extracellular-signalregulated-kinase 1/2 and m-calpain after axotomy of the sciatic nerve
- Low frequency repetitive transcranial magnetic stimulation improves motor dysfunction after cerebral infarction