不同氮磷比与水分添加对刨花楠光合生理特性的影响分析
2017-04-27邓兴宇钟全林范永香张中瑞郑跃芳裴盼
邓兴宇,钟全林,2,3,范永香,张中瑞,郑跃芳,裴盼
(1.福建师范大学地理科学学院,福建 福州 350007;2.湿润亚热带山地生态国家重点实验室培育基地,福建 福州 350007; 3.福建省植物生理生态重点实验室,福建 福州350007)
不同氮磷比与水分添加对刨花楠光合生理特性的影响分析
邓兴宇1,钟全林1,2,3,范永香1,张中瑞1,郑跃芳1,裴盼1
(1.福建师范大学地理科学学院,福建 福州 350007;2.湿润亚热带山地生态国家重点实验室培育基地,福建 福州 350007; 3.福建省植物生理生态重点实验室,福建 福州350007)
在全球变化的背景下,氮沉降现象普遍,降水分布不均,干旱变得更加频繁,植物对环境变化的响应与适应已成为亚热带地区研究的热点生态学课题之一。以亚热带珍贵常绿阔叶树种刨花楠(Machiluspauhoi)为对象,探讨不同氮磷比及水分添加对其光合生理特性各指标的影响。结果表明:(1)水分对刨花楠净光合速率(Pn)、光补偿点(LCP)、蒸腾速率(Tr)有显著影响,干旱胁迫下(田间持水量为40%)的刨花楠的Pn、LCP和Tr会明显下降,与正常水分条件(田间持水量80%)相比,Pn下降了52%,LCP下降了72%,Tr下降了38%;而水分对光饱和点(LSP)、气孔导度(Gs)、胞间CO2浓度(Ci)、水分利用效率(WUE)无显著影响;(2)不同氮磷添加对Pn、LSP、LCP、Gs、Ci、WUE等光合生理特性指标无显著影响。
氮磷比;水分;光合生理特性;刨花楠
光合作用是植物的重要生理过程,它既是能量转化过程,也是同化CO2并进行干物质积累从而形成初级生产力的过程;光合作用还是生态系统能量流动规律、植物生产力形成机制和全球碳平衡研究中的关键环节[1],植物的光合生理与生长密切相关,而施肥是调节作物生长发育的一项基本措施,提高光合速率是取得作物高产的主要途径[2]。N、P元素是控制陆地生态系统中植物生产力和代谢功能的关键因子,对植物的生长发育起着重要的作用,二者在植物体内存在着重要的相互作用[3,4]。植物叶片的氮磷比临界值被认为可以作为土壤对植物生长的养分供应状况的指标,并且氮磷比被广泛地用来诊断植物个体、群落和生态系统的氮、磷养分限制格局[5]。在一定范围内,随着土壤水分含量的增加,植物吸氮量及氮肥施用效果也逐渐提高,其变化还会影响养分的可利用性[6],进而对植物的生长发育起着重要作用。此外,受全球气候变化的影响,全球降水格局也在发生着变化[7],降水分布更加不均,例如中高纬度地区陆地表面降水增加,热带、亚热带地区陆地表面降水减少等[8],即使在非干旱地区季节性干旱也频繁发生[9],而干旱是限制植物生长的最频繁和严重的非生物胁迫因素。据预测,区域气候在未来的几十年里季节降水分布和夏季降水量将减少10%~30%[10]。因此,开展氮、磷养分与水分对植物光合生理特性的影响研究,对于研究未来我国南方亚热带地区季节性缺水环境下亚热带乔木树种培育与造林有着重要意义。
国内外有学者在施肥对植物光合生理特性的影响方面进行过相关研究,其研究对象主要为农作物及中高纬树种等[2,11,12]。有关水分对植物光合生理特性的影响方面,也主要集中在干旱条件或高寒条件下的探究[9,13],而对湿润亚热带区域常绿阔叶树种的影响研究较少,尤其是氮磷养分与水分对植物光合生理特性的耦合影响研究尚未见相关报道。为此,本文以湿润亚热带区域常绿阔叶树种刨花楠为对象,开展这方面研究,以期探讨我国南方亚热带地区季节性缺水与氮沉降是否会影响常绿阔叶树种的光合生理特性。
刨花楠(Machiluspauhoi)是亚热带常绿乔木阔叶林树种,既是珍贵的工业原料树种,又可作为观赏植物[14]。对于刨花楠的研究主要集中在育苗栽培技术及人工扦插繁殖[15,16]、生物量[17,18]、生长特性[19,20]、外界因素对其生长的影响[21]、种群结构及其分布格局[22]等,此外钟全林等人还对刨花楠的光响应特性[23]及刨花楠SLA与叶片 C、N、P 化学计量学的关系[24]等进行了研究。但有关氮磷养分与水分对刨花楠光合生理特性的耦合影响研究尚未报道。
本研究以1年生刨花楠实生苗为对象,通过不同处理的盆栽试验,测定不同氮磷添加及水分处理下刨花楠的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)及水分利用效率(WUE)等光合生理特性指标,探讨氮磷养分与水分,特别是不同氮磷比与水分对刨花楠光合生理特性的影响。研究结果将有助于理解刨花楠生长的影响机制与生存适应策略,对探究刨花楠对养分的资源利用效率以及选择适合刨花楠生长的最佳水肥管理措施等具有重要意义,并可为高效经营刨花楠人工林等提供理论指导和技术支持。
1 材料与方法
1.1 试验材料
本研究设在福建省南平市顺昌县林技中心进行,其地理坐标约为26°43′24″ N,117°38′37″ E,海拔约为552 m。该区域年平均气温为18.9 ℃,最冷月1月平均气温7.9 ℃,最热月7月平均气温28.1 ℃。无霜期为305 d,年平均降水量一般在1 600~1 900 mm,多集中在2—9月,约占全年降水量的86.6%。年平均日照时数约1 740.7 h。
以种子来源于江西省遂川县,并在顺昌县林技中心苗圃地培育的,大小规格基本相同的1年生刨花楠苗木为试验对象。试验前先准备硬质塑料花盆180个,花盆规格要求:上口径32 cm,下口径28 cm,高度30 cm。将来源于顺昌县造林地的底层山地红壤土混合拌匀,并按每盆8 kg将其装入盆内,该试验土的pH值及C、N、P含量分别为4.41、5.089 mg·g-1、0.769 8 mg·g-1和0.040 7 mg·g-1。按每盆栽植1株的方法将试验苗木移植到盛好土壤的盆内,共移植144盆,移植时间为2015年5月1日。在试验苗木上方搭建长12 m、宽5 m、高2 m的金属框架结构的防雨棚,防雨材料为加厚薄膜,在其上方加盖75%遮阳网,防雨棚四周不封闭,用于通风。对栽植后的苗木只进行常规的水分管理。在苗木移植盆内并生长了30 d时用油漆标出其地径固定测量位置,挂牌编号,测定各苗木的初始基径、株高、冠幅等指标,并开始实施N、P养分与水分相关试验。
1.2 试验设计
待苗木生长1个月并测定了其初始基径、株高、冠幅等指标后,开始进行模拟试验研究。考虑到区域N沉降特点及前期水分对刨花楠生长影响试验研究成果,以纯N添加量为100 kg·hm-2·a-1为标准,将N添加量固定,按N∶P比分别为8、10、12、15,即纯P添加量分别为12.5、10、8.33和6.67 kg·hm-2·a-1设置不同的P添加等级;水分按土壤田间持水量分别为80%和40%设置2个等级,另设置了2个不添加N、P,但水分分别为40%和80%的处理,以不添加N、P且水分为80%的处理作为对照(CK),共设置了10个处理,添加N、P的处理分别记作q1、q2…,q9,含对照CK(表1),每处理设2个重复,每重复9盆(即9株样木)。N用NH4NO3添加,P用P2O5添加。N、P添加方法是用电子天平按取相应质量的NH4NO3与P2O5,并将其完全溶于180 mL水中。每盆用移液枪吸取10 mL(合N添加量100 kg·hm-2·a-1)在距盆内侧1 cm处沿盆均匀滴注1圈,氮磷配施时间为7月1日。另预留一个装满土的空盆作为水分蒸发测定盆。每组选取一个标准盆,加水前称取其质量(精确到1 g),补充失去水分,每周补充3次。

表1 氮磷及水分添加处理
1.3 光合生理指标的测定
分别在处理前、处理12周时测定。选择典型晴朗天气,使用CIRAS-3便携式光合仪在上午9:30—11:00对每个处理组标准盆叶片的光合响应参数进行测定。光合有效辐射强度梯度设定依次为2 000、1 800、1 600、1 400、1 200、1 000、800、700、600、500、400、300、200、150、100、75、50、25、0 μmol·m-2·s-1,测定不同光合有效辐射强度下的光合参数指标。测定指标包括净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)等。暗呼吸速率在22:00时测定。利用不同光强对应的净光合速率采用二项式回归拟合光响应曲线,根据所得到的曲线方程,求得最大净光合速率(Pnmax)、光饱和点(LSP)、光补偿点(LCP),光饱和点为最大净光合速率所对应的光强,光补偿点为光响应曲线与X轴的交点的横坐标(净光合速率为零时所对应的光强)。并用内插法依次确定与最大净光合速率所对应的蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)、水分利用效率(WUE)的值。再利用双因素(氮磷,水分) 方差分析比较不同氮磷配比及水分添加对刨花楠最大净光合速率、光饱和点、光补偿点、蒸腾速率、气孔导度、胞间CO2浓度、水分利用效率等光合生理特性的影响,并分析讨论何种氮磷配比及水分最有利于刨花楠的生长。叶片WUE以公式WUE=Pn/Tr计算得出。
1.4 数据处理
数据处理利用SPSS20.0进行分析,通过双因素( 氮磷和水分)方差分析比较氮磷和水分对刨花楠光合生理生态特征的影响,图表制作工具为Excel2010。
2 结果与分析
2.1 不同处理下光响应曲线拟合
由图1可知,不同氮磷添加处理的Pn-PAR曲线均呈抛物线形式,而且增长速率都是先较大后来稍平缓。就不同处理而言,田间持水量为40%的Pn-PAR曲线变化相比田间持水量为80%的处理数值较低且较为平缓。处理2的曲线变化最为明显。
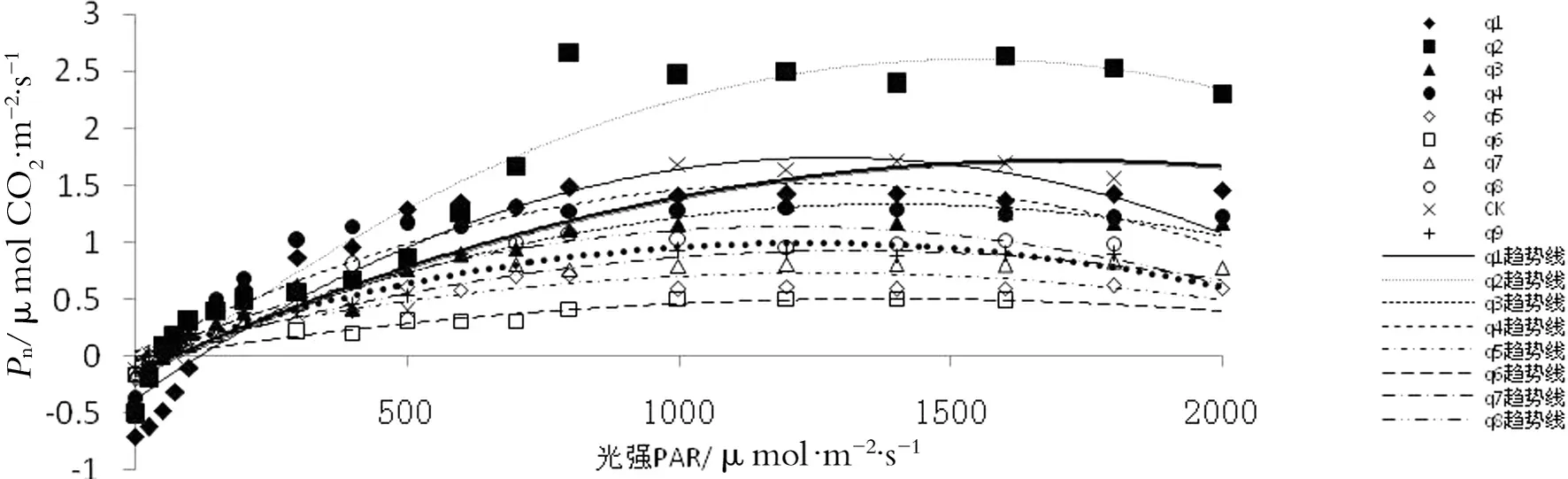
图1 不同处理下的光响应曲线
注:注:不同处理下光响应曲线方程如下(R2表示相关性,P表示显著性,N表示样本数):
q1:y1=-1.287×10-6x2+0.003 3x-0.371(R2=0.887,P<0.01,N=19)
q2:y2=-1.219×10-6x2+0.003 74x-0.272(R2=0.944,P<0.01,N=19)
q3:y3=-7.546×10-7x2+0.002 1x-0.136(R2=0.938,P<0.01,N=19)
q4:y4=-9.545×10-7x2+0.002 37x-0.04(R2=0.866,P<0.01,N=19)
q5:y5=-6.478×10-7x2+0.001 41x-0.023(R2=0.867,P<0.01,N=16)
q6:y6=-2.777×10-7x2+0.000 769x-0.027(R2=0.880,P<0.01,N=14)
q7:y7=-5.573×10-7x2+0.001 47x-0.045(R2=0.934,P<0.01,N=16)
q8:y8=-7.991×10-7x2+0.001 92x-0.012(R2=0.937,P<0.01,N=16)
CK:yCK=-6.371×10-7x2+0.002 18x-0.155(R2=0.957,P<0.01,N=17)
q9:y9=-8.01×10-7x2+0.001 81x-0.033(R2=0.954,P<0.01,N=15)
2.2 光合气体交换特征
紫坪铺工程在高速水流洞段普遍采用C50硅粉混凝土作为过流面材料,出口挑流鼻坎段全部采用C50硅粉混凝土,并在边墙铺1 cm厚环氧砂浆,保证一定抗冲耐磨能力。江坪河可研阶段在选用泄洪建筑物的抗冲耐磨混凝土材料过程中,进行了硅粉混凝土、聚羧酸高效减水剂、HF抗冲磨剂、掺玄武岩纤维等方案研究,设计推荐采用 “聚羧酸高效减水剂+掺玄武岩纤维”的方案。随着工程运行检验总结、建设财力增强和条件好转,我国西部地区新近设计中已较大幅度提高隧洞混凝土材料耐久性和强度等级标准。
2.2.1 不同氮磷比添加对刨花楠净光合速率(Pn)的影响 净光合速率(Pn)是光合作用强弱的重要指标,净光合速率的快慢直接反映出植物叶片合成有机物质能力的强弱,表明了植物积累营养物质和储存能量的能力[25]。从表2看出,不同的氮磷配比添加对刨花楠净光合速率没有显著性影响(P>0.05)。从多重比较结果来看,各处理组之间的差异性并不显著(P>0.05)但净光合速率N∶P=10较其他组高(表3),说明N∶P=10处理有利于提高刨花楠的净光合速率。从各处理组标准差来看,N∶P=10和N∶P=8两种处理的标准差比其他3个处理大得多,说明在施肥条件一致的情况下,不同的水分等级之间N∶P=10和N∶P=8两种处理的净光合速率存在较大差异。

表2 不同氮磷比与水分添加对刨花楠光合生理特性各指标的影响
注:*表示P<0.05,ns表示P>0.05
2.2.2 不同水分对刨花楠净光合速率(Pn)的影响 W1为田间持水量为80%的正常水分条件下的处理,W2为田间持水量为40%,即处于干旱胁迫条件下的处理(下同)。从表2水分对刨花楠净光合速率影响的方差分析可知,不同水分处理对刨花楠净光合速率具有显著影响(P<0.05)。从多重比较结果来看W1与W2也存在显著的差异性(表4),净光合速率W2较W1下降了52%。说明土壤水分是影响植物光合作用的重要因素,干旱胁迫下植物净光合速率会明显降低。
2.2.3 不同氮磷比添加及水分处理对刨花楠光饱和点(LSP)的影响 从表2可知,氮磷配比添加对刨花楠光饱和点无显著影响(P>0.05),多重比较的结果表明,各处理组之间并无显著差异,其光饱和点变动范围在1 100~1 450 μmolCO2·m-2·s-1之间。从表3可以看出CK的标准差明显高于其他组,说明在无氮磷添加情况下,不同水分处理之间的光饱和点存在较大的差异,但是氮磷配施的各处理组标准差较小,标准差N∶P=15 从表2水分对刨花楠净光合速率影响的方差分析可知,水分对刨花楠光饱和点无显著影响(P>0.05),从表4也可看出,不同水分等级处理下刨花楠的光饱和点差异并不明显,其变动范围在1 200~1 450 μmol CO2·m-2·s-1之间。 表3 不同氮磷比处理下的刨花楠叶片光合生理特性各指标特征 注:图中不同小写字母表示不同处理组之间具有显著的差异性。 表4 不同水分处理下的刨花楠叶片光合生理特性各指标特征 注:图中不同小写字母表示不同处理组之间具有显著的差异性。 2.2.4 不同氮磷比添加及水分处理对刨花楠光补偿点(LCP)的影响 由表2可知,不同氮磷添加对刨花楠光补偿点没有显著影响(P>0.05),各处理组之间也无显著差异性。但处理N∶P=15的光补偿点明显低于其他组,且N∶P=15 不同水分对刨花楠光补偿点影响显著(P<0.05)(表2)。多重比较的结果表明,不同水分等级之间刨花楠光补偿点存在显著差异(表4),W2的光补偿点仅有W1的28%,即干旱胁迫下刨花楠的光补偿点要比正常水分条件下刨花楠的光补偿点降低了72%,在一定程度上说明干旱条件下植物对弱光的利用能力更强。 2.3 不同氮磷比与水分添加对刨花楠蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)的影响 从表2可知,不同水分对刨花楠蒸腾速率有显著影响(P<0.05),不同水分等级之间刨花楠的蒸腾速率存在显著差异性(表4),干旱胁迫下的蒸腾速率较正常水分条件下蒸腾速率下降了38%。由此可见,在供水不足的情况下其蒸腾速率会受到明显的抑制,保证土壤水分充足可提高刨花楠的吸收和运输水分的能力,从而提高蒸腾速率,保证植株正常成长。 2.3.2 不同氮磷比添加及水分处理对刨花楠气孔导度(Gs)的影响 气孔是光合作用和呼吸作用过程中叶片内外进行气体交换的主要通道,气孔导度表示的则是气孔张开的程度,气孔导度控制着叶片内外气体交换的速率,因此气孔导度对光合作用、呼吸作用和蒸腾作用均有着十分重要的影响[27]。从表2可知,不同的氮磷添对刨花楠气孔导度无显著影响(P>0.05),多重比较结果也表明各处理组之间刨花楠气孔导度无显著差异。有氮磷添加的各处理组的标准差显著大于无氮磷添加的对照处理(表3),表明氮磷添加处理相比于无氮磷添加处理组不同水分等级之间刨花楠的气孔导度存在较大差异,说明氮磷添加会使得不同水分等级之间的气孔导度的差异性增大。 水分对刨花楠气孔导度并无显著影响(P>0.05)(表2)。从表4可以看出不同水分等级之间气孔导度的差异性不显著,但不同水分等级之间标准差较大,并且W1 2.3.3 不同氮磷比添加及水分处理对刨花楠胞间CO2浓度(Ci)的影响 由表2可知,不同氮磷比添加对刨花楠胞间CO2浓度(Ci)的影响不显著(P>0.05)。此外,不同氮磷比添加的各处理组之间差异性也并不明显,各处理胞间CO2浓度(Ci)值最大为192.50×10-6,最小为154.59×10-6,变动较小。仅从有氮磷添加的各处理组来看处理N∶P=12的标准差明显高于其他三组,且随着磷添加量的减少,标准差先增大后减小,说明同一氮磷添加不同水分等级之间的差异性随着磷添加量的减少先增大后减小,处理N∶P=12的差异性最大。 水分对刨花楠胞间CO2浓度的影响并不显著(P>0.05)(表2)。由表4可知,不同水分等级之间的差异性也不显著。标准差W1>W2,说明在正常的水分条件下,刨花楠的胞间CO2浓度的差异要大于干旱处理下的胞间CO2浓度差异。 2.4 不同氮磷添加及水分对刨花楠暗呼吸速率(Rd)的影响 由表3可知,不同氮磷添加对刨花楠暗呼吸速率无显著影响(P>0.05)。但多重比较的结果显示,CK与N∶P=8、N∶P=12和N∶P=15均存在显著差异性,暗呼吸速率N∶P=8>N∶P=15>N∶P=12>N∶P=10>CK,说明氮磷养分添加使得刨花楠的暗呼吸速率有明显提高。但N∶P=8、N∶P=10、N∶P=12和N∶P=15四种处理之间暗呼吸速率并无显著差异,其暗呼吸速率的值在0.31~0.41 μmol CO2·m-2·s-1,变动较小,说明了氮磷配比添加对刨花楠暗呼吸速率的影响不够显著。此外,CK标准差最大,有氮磷添加处理的标准差相对较小,N∶P=12和N∶P=15较小,N∶P=10接近零,说明无氮磷添加不同水分等级之间的刨花楠的暗呼吸速率差异性更大,有氮磷添加不同水分等级之间的暗呼吸速率差异性较小。由此可见氮磷添加可以在一定程度上缓解水分对刨花楠暗呼吸速率的影响。 水分对刨花楠暗呼吸速率无显著影响(P>0.05)(表3),从表4也可以看出,不同水分等级之间刨花楠的暗呼吸速率几乎无差异性,两者之间暗呼吸速率的值仅差0.02 μmolCO2·m-2·s-1。标准差W1>W2表明,正常水分条件下刨花楠暗呼吸速率的差异性要比干旱条件下暗呼吸速率的差异性大。 2.5 不同氮磷比与水分添加对刨花楠水分利用效率(WUE)的影响 水分利用效率是衡量植物物质生产和水分消耗之间关系的重要指标[28],影响水分利用效率的直接因子是光合作用和蒸腾作用,用Pn和Tr的比值来表示植物叶片水分利用效率(WUE),任何影响Pn和Tr的因子都对水分利用效率有不同程度的影响[29]。由表3可知,不同氮磷添加刨花楠水分利用效率(WUE)的影响未达到显著水平(P>0.05)。此外,各处理组之间的差异性也并不显著,其值在1.83~4.25 mmol·mol-1之间,但N∶P=10的水分利用效率较其他处理组高(表3),主要是因为N∶P=10的净光合速率较其他组高,而暗呼吸速率较其他组低,因此水分利用效率N∶P=10较高。N∶P=10的标准差较大且明显高于其他组,说明在N∶P=10处理下不同水分等级刨花楠的水分利用效率差异较其他组大。 由表3可知,水分对刨花楠水分利用效率的影响不显著(P>0.05),从表4可以看出不同水分等级之间的水分利用效率差异性也不明显,但W1略高于W2,W1的标准差值较W2大,表明同一水分条件下,刨花楠的水分利用效率的差异性为W1大于W2。 土壤水分是植物光合速率的主要限制因子之一[30,31],可通过测量叶片气体交换,尤其是净光合作用,气孔导度和蒸腾速率来反映干旱胁迫对植物的影响[32],光合作用对叶片水分亏缺十分敏感,当植物处于在轻度干旱胁迫时,光合速率的下降幅度较小,但随干旱胁迫加剧,光合速率开始大幅下降[33]。本研究也得出类似结果,处于干旱胁迫下的净光合速率较正常水分条件下的净光合速率下降了52%,气孔导度、胞间CO2浓度也较正常水分条件下低。原因可能是植物为了减少水分散失,缩小或关闭气孔,降低气孔导度,减少叶片的气体交换[34],限制了CO2的进入,从而对光合作用产生影响,使Pn降低。这与许大全的研究结果一致,即气孔限制是导致植物光合能力下降的因素之一[35]。在干旱胁迫下刨花楠的蒸腾速率下降了38%,这可能是由于刨花楠为了维持自身生理生化运转,通过降低蒸腾速率,减少植物体内水分的损失来应对胁迫环境,从而使其生长受到抑制。研究结果还表明干旱条件下,刨花楠的光补偿点会明显降低,其利用弱光的能力会增强。 光合作用是绿色植物生长的物质基础,是植物整个生命活动的重要生理过程,受到多种内外因素的影响。营养元素的作用主要是扩大光合面积,延长光合时间,提高光合能力[36]。鲁显楷等人通过模拟氮沉降试验, 不同强度水平的氮处理对光叶山黄皮和厚壳桂的最大净光合速率具有明显的促进作用[37]。张卫强在研究氮沉降对外来植物薇甘菊光合特性的影响发现,氮处理一定程度提高了薇甘菊的光合作用,提高了叶肉细胞CO2的固定能力,促进了气孔的开放,有利于CO2扩散到叶片内,为光合作用提供更多的原料,一定程度增加了薇甘菊最大光合潜力和对弱光与强光的利用能力[38]。本文研究结果表明,氮磷配比添加对刨花楠光合生理特性没有显著影响。分析其原因可能有以下几点:(1)刨花楠幼苗对外源施肥不够敏感,或者土壤中的氮磷可以满足其生长;(2)本试验研究周期较短,从而降低了氮磷添加效应;(3)氮磷添加会得刨花楠暗呼吸速率加快,而呼吸需要消耗更多的光合产物,进而影响其净光合速率的提高;(4)与研究区域条件、氮磷施肥水平以及试验时间尺度等有关,有待于以后进一步的深入研究[39]。 [1] 王冉,何茜,李吉跃,等.中国12种珍稀树种光合生理特性研究[J].东北林业大学报,2010,38(11):15-20 [2] 郭盛磊,阎秀峰,白冰,等.供氮水平对落叶松幼苗光合作用的影响[J].生态学报,2005,25(6):1291-1298 [3] Marklein A R,Houlton B Z.Nitrogen inputs accelerate phosphors cycling rates across a wide variety of terrestrial ecosystems[J].New Phytologist,2012,193(3):695-704 [4] Agren G I.Stoichiometry and nutrition of plant growth in natural communities[J].Annual Review of Ecology Evolu-tion and Systematics.2008,39:153-170 [5] 宋彦涛,周道玮,李强,等.松嫩草地80种草本植物叶片氮磷化学计量特征[J].植物生态学报,2012,36(3):222-230 [6] Lu D Q,Yang J R,Ma L Y.Study on effect of irrigation on soil nitrate leaching and uptake[J].Plant Nutrition and Fertilizer Science,1999,5(4):307-315 [7] Solomon S,Qin D,Manning M,et al.Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change[M].Cambridge University Press,Cambridge,United Kingdom/New York,NY,USA,2007 [8] Easterling D ,Rmeehl G A,Parmesan C,et al.Climate extremes:observations,modeling,and impacts[J].Science,2000,289(5487):2068-2074 [9] Zhu X C,Song F B,Liu S Q,et al.Arbuscular mycorrhizaeimproves photosynthesis and water status of ZeamaysL.under drought stress [J].Plant Soil Environ,2012,58:186-191 [10] Cubasch U,Kadow C.Globalclimate change and aspects of regional climate change in the Berlin-Brandenburgregion[J].Erde,2011,142(1-2):3-20 [11] 张丽光,李丹,刘磊,等.不同施肥种植模式对玉米光合特性养分效率及产量性状的影响[J].水土保持学报,2013,27(2):115-125 [12] 何茜,苏艳,晏紫伊,等.增施氮肥对欧洲云杉光合生理特性的影响[J].生态科学,2015,34(3):109-115 [13] 于文颖,纪瑞鹏,冯锐,等.不同生育期玉米叶片光合特性及水分利用效率对水分胁迫的响应[J].生态学报,2015,(35)9:2902-2909 [14] 徐朝斌,钟全林,程栋梁,等.江西4刨花楠种源苗木叶片表型性状与生物量分配的比较[J].安徽农业大学学报, 2012, 39(6): 920-924 [15] 胡松竹,钟全林,黄志强,等.刨花楠人工栽培技术初探[J].江西农业大学学报,2001,23(3):332-335 [16] 徐奎源,徐永星,徐裕良.红楠等4种楠木树种的栽培试验[J].江苏林业科技,2005,32(2):26-27 [17] 钟全林,张振瀛,张春华,等.刨花楠生物量及其结构动态分析[J].江西农业大学学报,2001,23(4):533-536 [18] 邓仕坚,廖利平,汪思龙,等.湖南会同红栲-青冈-刨花楠群落生物生产力的研究[J].应用生态学报,2000,11(5):651-654 [19] 廖龙泉.刨花楠生长规律的初步研究[J].江苏林业科技,1997,24(1):39-41 [20] 钟全林,胡松竹,黄志强,等.刨花楠生长特性及其生态因子影响分析[J].林业科学,2002,38(2):165-168 [21] 鲁美娟,江洪,李巍,等.模拟酸雨对刨花楠幼苗生长和光合生理的影响[J].生态学报,2009,29(11):5986-5994 [22] 朱宏光,杨柳,温远光,等.广西大明山刨花润楠种群结构[J].广东农业科学,2015,(12):147-151 [23] 钟全林,程栋梁,胡松竹,等.刨花楠和华东润楠叶绿素含量分异特征及与净光合速率的关系[J].应用生态学报,2009,20(2):271-276 [24] 徐朝斌,钟全林,程栋梁,等.基于地理种源的刨花楠苗木比叶面积与叶片化学计量学关系[J],生态学报,2015,35(19):6510-6515 [25] 谢田玲,沈禹颖,邵新庆,等.高原4种豆科牧草的净光合速率和蒸腾速率日动态及水分利用效率[J].生态学报.2004,24(8):1678-1685 [26] 关义新,裁俊英,林艳.水分胁迫下植物叶片光合的气孔和非气孔限制[J].植物生理学报,1995,31(4):293-297 [27] 孟繁静.植物生理学基础[M].北京:农业出版社,1987 [28] 周国逸.生态系统水热原理及其应用[M].北京:气象出版社,1997 [29] Radin J W.Reconciling water-use efficiencies of cotton in field and laboratory. Crop Science. 1992;32(1):13-8 [30] Colom M R,Vazzana C.Drought stress effects on three cult-ivars of Eragrostiscurvula:photosynthesis and water relations[J].Plant Growth Regulation,2001,34:195-202 [31] Raendrudu G,Naidu C V,MallikrjunaK.Effect of water stress on photosynthesis and growth in two teak phenotypes[J].Photosynthetica,1999,36(4):627-630 [32] Nichols S N,Hofmann R W,Williams W M.Physiological drought resistance and accumulation of leaf phenolics in white clover interspecific hybrids[J].Environmental and Experimental Botany,2015,119:40-47 [33] Xu N, Guo W, Liu J, et al. Increased nitrogen depositionalleviated the adverse effects of drought stress on Quercusvariabilis and Quercusmongolica seedlings[J].ActaPhysiologiaePlantarum, 2015, 37(6): 1-11 [34] 张淑勇,张光灿,陈建,等.土壤水分对五叶爬山虎光合与蒸腾作用的影响[J].中国水土保持科学,2006,4(4):62-66 [35] 许大全.光合作用效率[M].上海:上海科学技术出版社,2002:163-170 [36] 武维华.植物生理学[M].北京:科学出版社,2003 [37] 鲁显楷,莫江明,李德军,等.鼎湖山主要林下层植物光合生理特性对模拟氮沉降的响应[J].北京林业大学学报,2007,29(6):1-9 [38] 张卫强,肖辉林,殷祚云,等.模拟氮沉降对入侵植物薇甘菊光合特性的影响[J].生态环境学报,2013,22(12): 1859-1866 [39] 吴永宏,钟全林,程栋梁,等.氮、水处理对刨花楠一年生扦插苗光合特性和生物量分配的影响[J].安徽农业大学学报, 2016, 43(2): 1-6 Effect of N/P Ratio and Water on Photosynthetic Physiological Characteristics ofMachiluspauhoi Deng Xingyu1, Zhong Quanlin1,2,3, Fan Yongxiang1, Zhang Zhongrui1,Zheng Yuefang1,Pei Pan1 (1. College of Geographical Sciences,Fujian Normal University, Fuzhou 350007, China; 2. State Key Laboratory Breeding Base of Humid Subtropical Mountain Ecology, Fuzhou 350007, China; 3.Institute of Geography, Fujian Normal University, Fuzhou 350007, China) Under the background of global change, phenomenon of nitrogen deposition generally exists, with an uneven distribution of precipitation, and more and more frequent drought. Response and adaptation of plant to environmental change has become one of the hot ecological subjects in subtropical region. Taking an excellent subtropical evergreen broad-leaved tree species--Machiluspauhoi,as research object, the impact of different N:P ratios and water addition on indexes of photosynthetic physiological characteristics was discussed. Result indicates that: (1) Water have significant influences on net photosynthetic rate (Pn), light compensation point (LCP) and transpiration rateTrofMachiluspauhoi. Under dry conditions (field capacity being 40%),Pn,LCP& transpiration rate (Tr) reduce sharply. Compared with normal water conditions (field capacity being 80%),Pn,LCP&Trdecrease 52%, 72% & 38%, respectively. But water have no obvious effects on light saturation point (LSP), stomatal conductance (Gs), intercellular CO2concentration (Ci) & water use efficiency (WUE);(2)Different nitrogen & phosphorus addition have no obvious effects onPn,LSP,LCP,Gs,Ci&WUE. N/P ratio;water;photosynthetic physiological characteristics;Machiluspauhoi 1005-5215(2017)04-0004-07 2017-02-27 国家自然科学基金项目(31170596,31170374,31370589);福建省种业创新和产业化工程项目(2014S1477-4);福建省科技厅重大项目(2014N5008) 邓兴宇(1992-),男,硕士,从事自然资源与环境研究,Email:17805966210@163.com S792 A 10.13601/j.issn.1005-5215.2017.04.002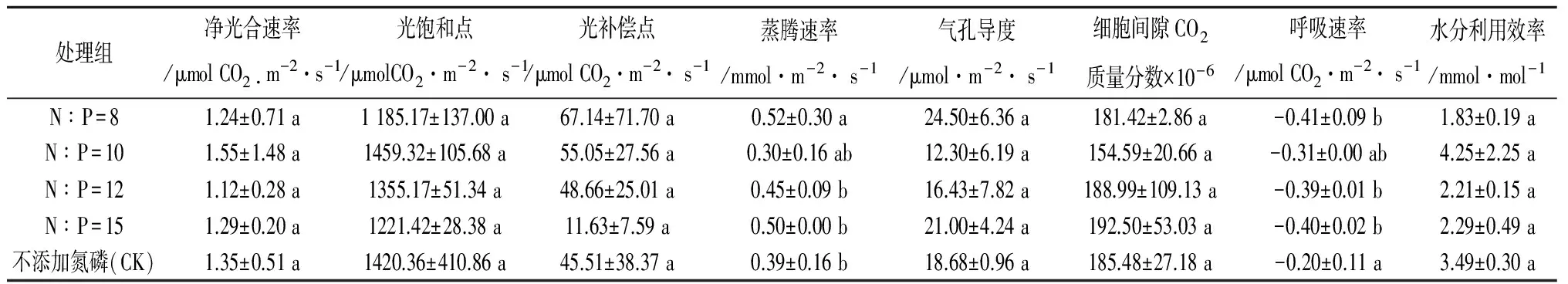

3 结论与讨论
