高山松不同种源苗木在云南松生境下的生物量与生长分析
2017-04-27朱存福许玉兰蔡年辉
周 丽,陈 诗,朱存福,许玉兰,李 悦,李 伟,蔡年辉
(1.攀枝花市国营林场总场,四川 攀枝花 617061;2.西南林业大学林学院,云南 昆明 650224;3.北京林业大学生物科学与技术学院,北京 100083;4.云南吉成园林科技股份有限公司,云南 弥勒 652300)
高山松不同种源苗木在云南松生境下的生物量与生长分析
周 丽1,2,陈 诗2,朱存福2,许玉兰2,李 悦3,李 伟4,蔡年辉2
(1.攀枝花市国营林场总场,四川 攀枝花 617061;2.西南林业大学林学院,云南 昆明 650224;3.北京林业大学生物科学与技术学院,北京 100083;4.云南吉成园林科技股份有限公司,云南 弥勒 652300)
以8个高山松种源4年生苗木为研究对象,测定其生物量和生长性状,并对各组分生物量的模型进行拟合,初步选出苗木质量较好的高山松优良种源和预估精度较高的生物量模型。结果表明:高山松各种源的生长性状除Ⅰ级侧根数和侧枝数的差异不显著外,其余性状均存在极显著差异;种源间各组分生物量均存在极显著差异,地上部分生物量是苗木生物量的主体部分,约占整株生物量的75.60%,地上部分生物量和整株生物量从大到小均表现为:西藏波密帕隆藏布江(PL)种源>四川宝兴(BX)种源>西藏林芝尼洋河(NY)种源>四川康定(KD)种源>西藏察隅(CY)种源>西藏雅鲁藏布江(YL)种源>四川理县(LX)种源>四川马尔康(MK)种源,各器官生物量的分配比例大小顺序则不尽相同;种源间除侧枝含水率外,其余组分的含水率均存在极显著差异;地径与各组分生物量之间的相关性均比苗高与各组分生物量之间的相关性更加密切,最优模型以地径(D)、地径—苗高(D2H)为自变量的幂函数生物量模型。综合比较发现,西藏波密帕隆藏布江(PL)种源的苗木质量较好,积累的干物质较多,能更好地适应试验地环境,有较大的碳储量和碳汇能力。
生物量;生长性状;模型;高山松;天然杂交种;杂种优势;含水率
高山松(Pinus densata Master.)是一种高海拔的松科松属植物[1],是已被证实的自然同倍体杂交种,在青藏高原东南缘海拔2 700~4 200 m的广大地域形成大面积稳定的纯林[2],其自然分布区与云南松自然分布区的北缘和油松自然分布区的南缘相接[3]。早期,对高山松的研究主要集中于其同倍体杂交起源方面,吴中伦[4]和管中天[5]分别于1956年和1981年从形态学和解剖学方面提出高山松可能是云南松(P. yunnanensis Franch.)与油松(P. tabulaeformis Carr.)的杂交种;王晓茹等[6-7]于1990年对云南松、高山松和油松3个树种的等位酶进行研究,首次从分子遗传学角度证实高山松起源于云南松和油松的天然杂交种。此后,从同工酶、cpDNA、mtDNA、rDNA和染色体构相的遗传分析也得到证实[8-14]。对高山松生态位的研究发现,3个树种各自占据了适合生存的生态位,在地理位置上形成一个高山松居中、云南松在南、油松在北的替代模式,高山松占据了其亲本种较难正常生存和繁衍的高海拔生境[15-16]。近年来,对高山松的研究主要包括苗木生长节律及变异规律[3]、种群结构及动态特征[17]、种群分布格局[18]、林分蓄积量估测及模型构建[19-22]、苗期生长[23-24]以及光合生理[25]等方面,但对于苗期高山松在云南松生境下的生长以及与生物量关系的报道较少。本试验通过对8个高山松种源4 年生苗木的各组分生物量进行了测定分析,旨在了解在云南松生境下高山松的生长适应性及其差异性,初步评选出生物量及生长性状较优良种源,为培育和管理高山松提供参考,通过对各组分生物量模型的构建和优选,还可为高山松苗木生物量的估算以及含碳系数、储能系数的测定提供依据,对研究碳循环和能量流动具有重要意义。
1 材料与方法
1.1 试验地概况
试验地设在西南林业大学的苗圃,地处102°45′41′E、25°04′00′N,海拔1 945 m云贵高原中部云南松中心分布区,年均气温15℃,年降水量700~1 100 mm,全年明显的分为干、湿两季,属于北亚热带半湿润高原季风气候。全年晴天较多,年均日照率约为56%,年均总辐射量达543.23 kJ/cm2。
1.2 试验材料
试验材料为采自于高山松天然分布区内的成熟球果的种子培育而成的4年生苗木,各种源种子采集地的地理位置见表1。采用完全随机区组的试验设计设置5次重复,每个重复播种3行,每行播20粒,发芽后进行正常的苗期管理。

表1 高山松采样种源的地理位置
1.3 试验方法
1.3.1 材料收集 采用“全挖法”挖取高山松各种源的4年生苗木,挖取苗木时尽量保持根系完整,每个种源挖取12~15株苗木,从中随机选取根系保留较完整的9株用于生长特性及形态指标、生物量的测定。生长特性及形态指标测定主要包括苗高、地径、Ⅰ级侧根数(根长≥ 1 cm)[26]、主根长、侧枝数、侧枝直径、侧枝长等。
1.3.2 生物量测定 用误差±0.01 g电子天平分别称量地下部分(根)和地上部分(主干、侧枝、针叶)各器官的鲜质量,然后在105℃烘箱中杀青30 min后于80℃烘干至恒重,测量各器官的干质量,即为生物量。苗木各器官的含水率[27]在一定程度上反映了苗木干物质的积累程度,计算公式为:

1.3.3 苗木各组分生物量模型构建 常用的生物量模型有幂函数模型(3个)、指数函数模型(4个)和多项式模型(4个)3种类型的11个模型[28](表2),依据11个模型,以各器官生物量与地径(D)、地径—苗高(DH)的关系为基础,进行高山松各组分生物量预估模型构建。
1.4 数据处理
使用Excel2007进行数据的录入整理和图形绘制。根据统计学百分数≤30%或者≥70%的数值不满足方差齐性检验的原理[29],各组分生物量的分配比例、含水率均有超出此范围的数据,因此,对这部分数据先进行反正弦转化,再采用SPSS17.0软件进行单因素方差分析。

表2 常用的生物量模型
2 结果与分析
2.1 高山松不同种源苗木生长性状的测定
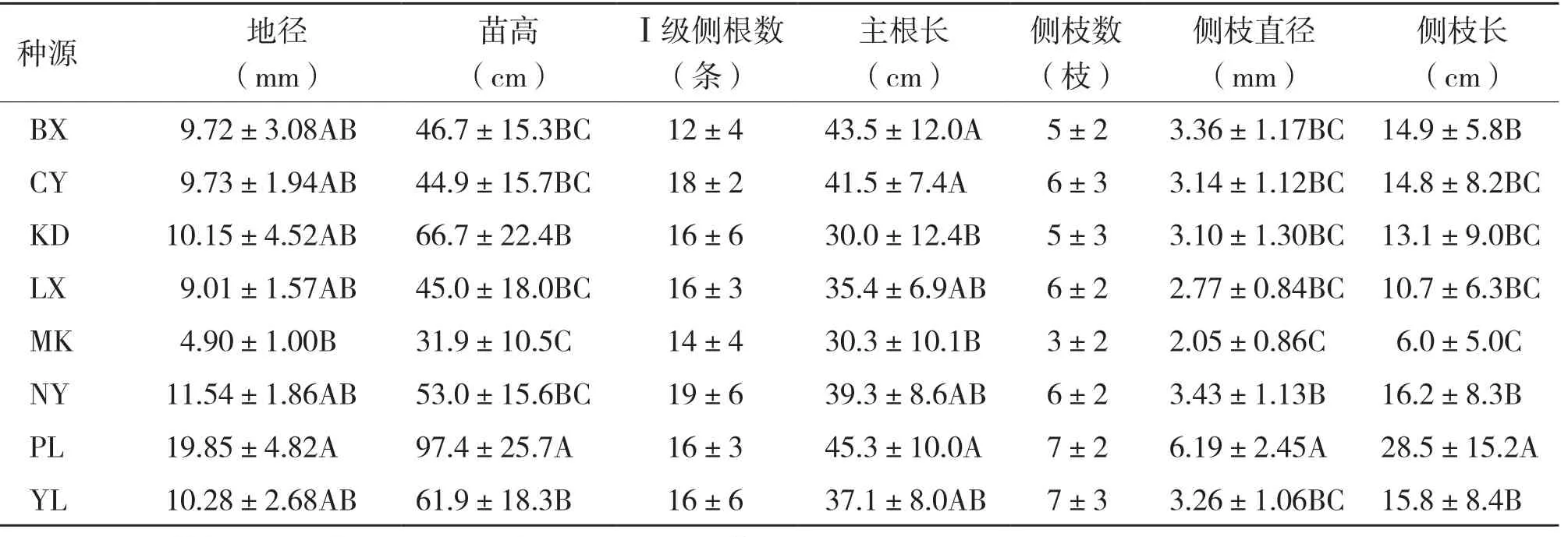
表3 高山松不同种源苗木生长性状分析
对高山松不同种源试验苗木的生长量及形态指标进行统计分析,结果见表3。
由表3可知,高山松不同种源间的地径、苗高、主根长、侧枝直径和侧枝长均存在极显著差异,Ⅰ级侧根数和侧枝数的差异不显著。8个高山松种源的地径、苗高、Ⅰ级侧根数、主根长、侧枝数、侧枝直径和侧枝长的平均值分别为10.65(±4.86) mm、55.9(±25.5) cm、16(± 5)条、37.8(±10.6) cm、6(±3)枝、3.54 (±1.79) mm和15.8(±10.8) cm,其中,PL种源的地径、苗高、侧枝直径和侧枝长分别高于平均值86.4%、74.2%、74.9%和80.4%,极显著高于其他种源。
2.2 高山松不同种源苗木各器官的生物量及其分配比例
对高山松8个种源苗木的各器官(根、主干、侧枝和针叶)生物量及其分配比例、地上部分生物量和整株生物量进行统计分析,结果见表4和图1。

表4 高山松不同种源苗木各组分生物量(g)比较
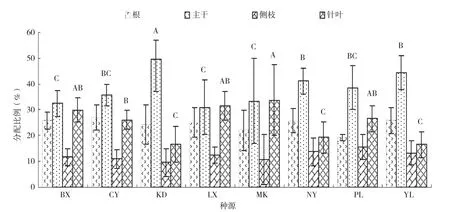
图1 高山松不同种源苗木各器官生物量的分配比例
由表4可知,高山松不同种源间的各器官生物量、地上部分生物量和整株生物量均存在极显著差异。种源间,PL种源的各器官生物量、地上部分生物量和整株生物量均最大,分别为29.52、59.74、22.54、42.18、124.45、153.98 g,均极显著高于其余种源,分别高于平均值231.69%、277.15%、315.59%、306.75%、293.46%和279.92%。各种源地上部分生物量和整株生物量从大到小均表现为:种源PL>BX>NY>KD>CY>YL>LX>MK,各器官生物量的大小顺序则不尽相同,但主要表现为:主干>针叶>根>侧枝。8个种源各器官生物量、地上部分生物量和整株生物量的变异系数均较大,多集中于50%~70%,其中,KD种源的各器官生物量、地上部分生物量和整株生物量以及MK种源侧枝生物量的变异系数均大于100%,CV分别为117.85%、118.04%、118.17%、140.10%、120.90%、119.58%和101.00%。
由高山松不同种源苗木各器官生物量的分配比例(图1)可知,8个高山松种源的主干生物量和针叶生物量的分配比例均存在极显著差异,根生物量和侧枝生物量的分配比例差异不显著。8个种源侧枝生物量的分配比例均最小,为9.60%~15.58%;主干和针叶生物量的分配比例较高,约为38%和25%;地上部分生物量是苗木生物量的主体,占整株生物量75.60%,波动于72.95%~80.68%,其分配比例从大到小表现为:种源PL>MK>KD>LX>NY>BX>YL>CY;根生物量的分配比例波动于19.32%~27.05%。
2.3 高山松不同种源苗木各器官的含水率
对高山松不同种源苗木各器官、地上部分和整株的含水率反正弦转化后进行统计分析,结果见表5。
由表5可知,8个高山松种源的根、主干、针叶、地上部分和整株含水率均存在极显著差异,侧枝含水率则差异不显著。种源MK和PL的各组分含水率均较大,其中,MK种源根和主干的含水率均最大,分别为61.92%和60.83%;而PL种源的侧枝、地上部分和整株含水率均最大,分别为55.80%、59.40%和59.45%。同一种源内的各组分含水率间,针叶含水率均最大(除MK种源为主干含水率最大外),侧枝含水率均最小(除LX种源为根含水率最小外),种源间CY、NY和YL种源3个种源各组分含水率的大小顺序相同,表现为针叶>地上部分>整株>主干>根>侧枝,其余种源各组分含水率的大小顺序均不相同。各组分含水率的变异系数集中于4%~17%,于2.45%~45.20%之间波动,最大的是MK种源的侧枝含水率,最小的是PL种源的针叶含水率。

表5 高山松不同种源苗木各组分含水率(%)比较
2.4 高山松不同种源苗木各组分生物量与生长性状的相关性分析
对高山松不同种源苗木各组分生物量与生长量进行相关性分析,结果见表6。由表6可知,生长性状(苗高、地径)和各组分生物量两两之间均存在极显著正相关,地上部分生物量与整株生物量的相关系数最大(r=0.999)。地径与各组分生物量之间的相关性均高于苗高与各组分生物量之间的相关性,其中,地径与整株生物量的相关性最大(r=0.919),苗高与主干生物量的相关性最大(r=0.833);各组分生物量之间的相关系数均较大,除主干与侧枝生物量的相关系数为0.866外,其余的均大于0.9。
2.5 高山松各组分生物量模型的拟合及优选
以高山松的各器官生物量和整株生物量为因变量,D、D2、DH和D2H为自变量,根据表2中的生物量模型进行回归拟合,得出55个生物量模型(每个模型5个),以决定系数(R2)和均方差(MSE)综合评价模型的拟合效果,综合比较选出高山松苗木各器官和整株生物量的5个最优模型,R2波动于0.970~0.997,MSE波动于0.163~20.248,结果见表7。
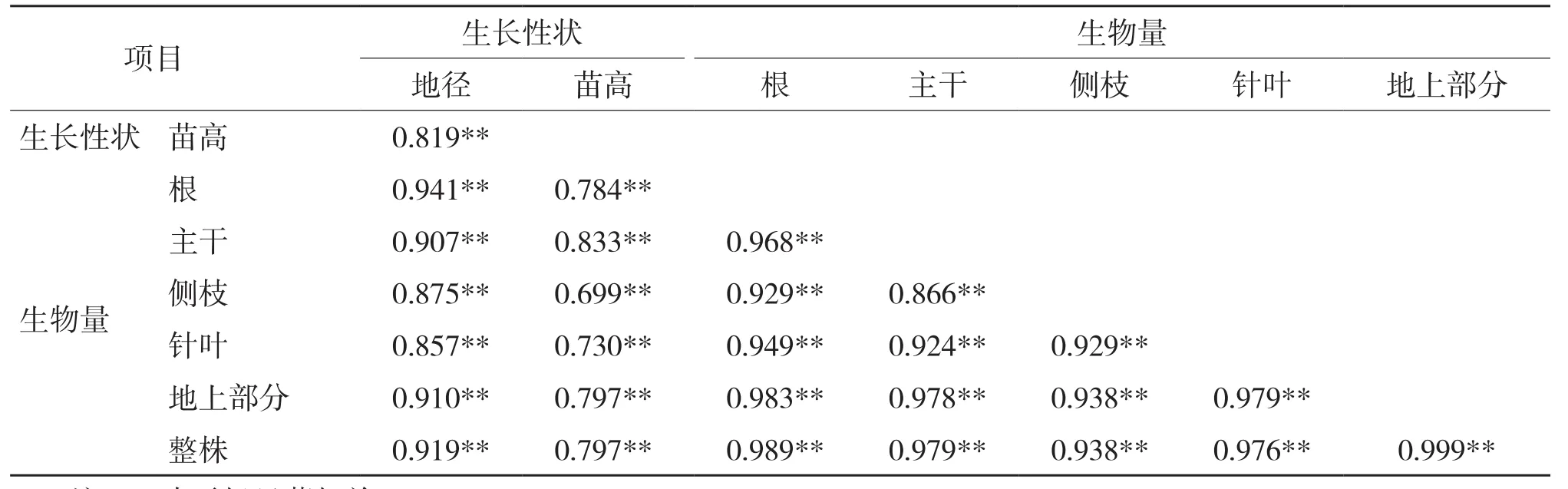
表6 高山松不同种源苗木各器官生物量与生长性状的相关性
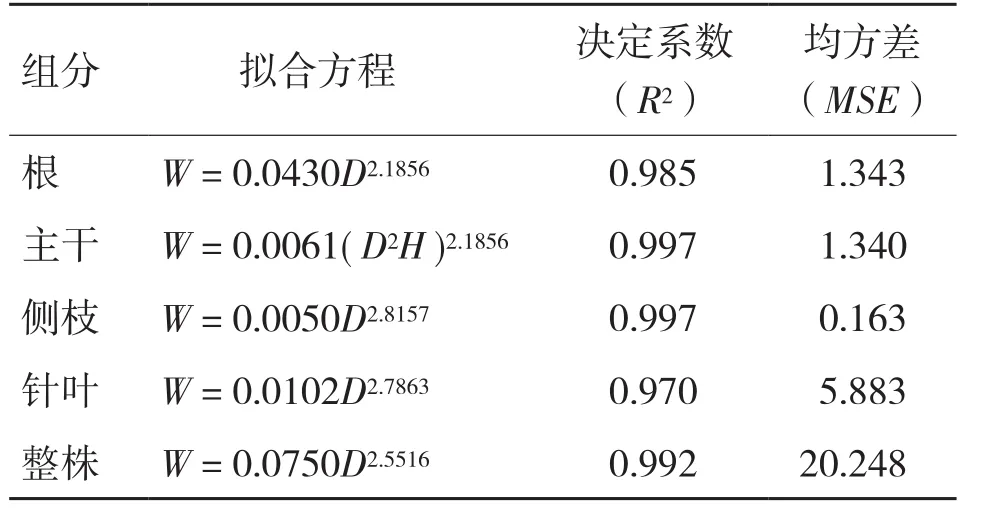
表7 高山松苗木各组分生物量的估算优选模型
由表7可知,幂函数生物量模型的拟合效果最好,拟合R2波动于0.959~0.997,MSE波动于0.163~49.850,均优于指数模型和二项式模型。最优模型中除主干生物量的最优估算模型是以D2H为自变量外,其余模型均是以D为自变量(模型形式为),其模型数占总模型数的80%。
3 结论与讨论
地径和苗高作为评价苗木质量的优先指标,可以反映苗木在一定时间营养物质的积累情况[30]。对8个高山松种源的生长性状进行分析,结果表明,PL种源在云南松生境下的生长状况较好,其苗木具有较好的质量,能很好地适应试验地的环境。
生物量的测定为测定含碳系数、储能系数奠定基础,含水率的测定可以反映一定阶段植株对干物质的积累程度,反映植物对环境的适应能力[31-32]。对高山松不同种源的各组分生物量及其含水率的分析比较结果表明,各组分生物量及其含水率(除侧枝含水率差异不显著外)均存在极显著差异,其中,PL种源具有最大的生物量和较大的含水率。植株的栽植密度对林木各器官生物量的分配比例有较大影响,对高山松不同种源各组分生物量及其分配比例的研究表明,种源间根生物量的分配比例均大于侧枝生物量,这在其他研究中也有相似报道[33]。根生物量较大,说明植株的根系较大,能更好地保持土壤不被流失,也能更多的从土壤中获取养分;侧枝生物量较小,对于用材树种的可用木材较大,此外,在相同栽植密度的林地中,林木的侧枝较少,则林分的郁闭度降低,利于树种的天然更新。
对高山松各组分生物量与苗高、地径的相关性分析表明,苗高、地径与各组分生物量均存在极显著相关,其中,地径与各组分生物量之间的相关性均比苗高与各组分生物量之间的相关性更加密切,该结果与薛云英[34]的研究结果相似,因此,在生物量模型参数的选择上主要以地径(D、D2)和地径—苗高(DH、D2H)的综合作用为自变量。本研究对高山松各组分生物量模型的拟合及优选的结果表明,采用幂函数模型拟合的生物量模型的R2较大,MSE相对较小,拟合效果较好,指数函数模型的拟合效果次之,多项式模型最差,与李燕等[35]研究结果相似,且最优模型主要是以D为自变量,只有主干的最优模型是以D2H为自变量。因此,以D、D2H为自变量的幂函数生物量模型对预测高山松苗木生物量有较高的精确性和准确度。
综合比较发现,PL种源的地上部分生长较好,苗木粗壮,根系较深且发达,各器官生物量、地上部分生物量和总生物量均最大,各组分的含水率也较高,对环境的适应能力较强,固定和积累的干物质较多,具有较大的碳储量和碳汇能力。
[1]Xing F Q,Mao J F,Meng J X,et al. Needle morphological evidence of the homoploid hybrid origin of Pinus densata based on analysis of artificial hybrids and the putative parents,Pinus tabuliformis and Pinus yunnanensis[J]. Ecology Evolution,2014,4(10):1890-1902.
[2]毛建丰,李悦,刘玉军,等. 高山松种实性状与生殖适应性[J]. 植物生态学报,2007,31 (2):291-299.
[3]蔡年辉,许玉兰,白青松,等. 不同种群高山松1年生播种苗木生长节律及其变异[J]. 东北林业大学学报,2013,41(5):11-15,74.
[4]吴中伦. 中国松属的分类与分布[J]. 植物分类学报,1956,5(3):131-163.
[5]管中天. 四川松杉类植物分布的基本特征[J].植物分类学报,1981,19(4):393-407.
[6]Wang X R,Szmidt A E,Lewandowski A,et al. Evolutionary analysis of Pinus densata Masters,a putative Tertiary hybrid. 1. Allozyme variation [J]. Theoretical and Applied Genetics,1990,80(5):635-640.
[7]Wang X R,Szmidt A E,Lewandowski A,et al. Evolutionary analysis of Pinus densata Masters,a putative Tertiary hybrid. 2. A study using species specific chloroplast DNA markers[J]. Theoretical and Applied Genetics,1990,80 (5):641-647.
[8]Wang X R,Szmidt A E. Hybridization and chloroplast DNA variation in a Pinus species complex from Asia[J]. Evolution,1994,48 (4):1020-1031.
[9]Yu H,Ge S,Hong D Y. Allozyme diversity and population genetic structure of Pinus densata Master in northwestern Yunnan,China[J]. Biochemical Genetics,2000,38(5):139-148.
[10]Wang X R,Szmidt A E,Savolainen O. Genetic composition and diploid hybrid speciation of a high mountain Pine,Pinus densata,native to the Tibetan Plateau[J]. Genetics,2001,159(1):337-346.
[11]Song B H,Wang X Q,Wang X R,et al. Maternal lineages of Pinus densata,a diploid hybrid[J]. Molecular Ecology,2002,11(6):1057-1063.
[12]Song B H,Wang X Q,Wang X R,et al. Cytoplasmic composition in Pinus densata and population establishment of the diploid hybrid pine[J]. Molecular Ecology,2003,12(11):2995-3001.
[13]Liu Z L,Zhang D,Hong D Y,et al. Chromosomal localization of 5S and 18S-5. 8S-25S ribosomal DNA sites in five Asian pines using fluorescence in situ hybridization[J]. Theoretical and Applied Genetics,2003,106(2):198-204.
[14]Liu Z L,Zhang D M,Wang X Q,et al. Intragenomic and interspecific 5S rDNA sequence variation in five Asian Pines[J]. American Journal of Botany,2003,90(1):17-24.
[15]Mao J F,Wang X R. Distinct niche shift characterizes the homoploid hybrid speciation of Pinus densata on the Tibetan Plateau[J]. The American Naturalist,2011,177(4):424-439.
[16]梁冬,毛建丰,赵伟,等. 高山松及其亲本种群在油松生境下的苗期性状[J]. 植物生态学报,2013,37(2):150-163.
[17]卢杰,郭其强,郑维列,等. 藏东南高山松种群结构及动态特征[J]. 林业科学,2013,49 (8):154-160.
[18]沈志强,卢杰,华敏,等. 西藏色季拉山高山松种群点格局分析[J]. 西北农林科技大学学报(自然科学版),2016,44(5):73-81.
[19]付虎艳,徐云栋,李圣娇,等. 基于SVM方法的高山松林蓄积量遥感估测研究[J]. 西部林业科学,2014,43(4):83-88.
[20]徐云栋,舒清态,李圣娇,等. 基于优化BP神经网络的香格里拉高山松蓄积量模型研究[J].西北林学院学报,2015,30(6):190-195.
[21]王永刚,舒清态,李圣娇,等. 香格里拉高山松天然林林分蓄积混合效应模型构建[J]. 西南林业大学学报,2016,36(3):121-125.
[22]孙雪莲,熊河先,胥辉,等. 高山松天然林单木生物量因子模型构建[J]. 林业资源管理,2016 (3):49-53,60.
[23]代剑峰,孟景祥,毛建丰,等. 油松×云南松人工杂种与亲本种在高山松生境下的苗期指标对比分析[J]. 北京林业大学学报,2014,36 (1):8-14.
[24]Xu Y L,Cai N H,He B,et al. Germination and early seedling growth of Pinus densata Mast. Provenances[J]. Journal of Forestry Research,2016,27(2):283-294.
[25]高琼,王维有,孟景祥,等. 油松×云南松杂种与亲本种和高山松的光合特性比较[J]. 北京林业大学学报,2016,38(2):37-43.
[26]张锁,樊军锋,刘永红,等. 陕西省油松不同种源和家系苗期性状的遗传变异分析[J]. 西北农林科技大学学报(自然科学版),2010,38 (4):64-70.
[27]吴志树. 4种林木幼苗含水率与生物量的关系研究[J]. 现代农业科技,2014(21):156-157,159.
[28]孟晓清. 大兴安岭地区落叶松生物量模型及碳储量研究[D]. 北京:北京林业大学,2014.
[29]袁志发,周静芋. 试验设计与分析[M]. 北京:高等教育出版社,2000.
[30]闫彩霞,杨锦昌,尹光天,等. 米老排不同高度级苗木形态特征的分析[J]. 林业资源管理,2013(5):98-102.
[31]陈振雄,贺东北. 南方马尾松含水率特征及其模型研建[J]. 中南林业调查规划,2011,30 (2):56-60,64.
[32]王月海,许景伟,韩友吉,等. 黄河三角洲五个耐盐树种苗木生物量比较[J]. 林业科技开发,2013,27(4):52-55.
[33]刘斌,刘建军,任军辉,等. 贺兰山天然油松林单株生物量回归模型的研究[J]. 西北林学院学报,2010,25(6):69-74.
[34]薛云英. 长白山几种树种幼树生物量及其模型研究[J]. 黑龙江科技信息,2013(31):248.
[35]李燕,张建国,段爱国,等. 杉木人工林生物量估算模型的选择[J]. 应用生态学报,2012,21 (12):3036-3047.
(责任编辑 白雪娜)
Biomass and growth analysis of different provenances of Pinus densata seedling in P. yunnanensis habitat site
ZHOU Li1,2,CHEN Shi2,ZHU Cun-fu2,XU Yu-lan2,LI Yue3,LI Wei4,CAI Nian-hui2
(1.Panzhihua State Forest Center,Panzhihua 617061,China;2. College of Forestry,Southwest Forestry University,Kunming 650224,China;3.College of Biological Science and Technology,Beijing Forestry University,Beijing 100083,China;4.Yunnan Jicheng Landscape Technology Co.,Ltd., Mile 652300,China)
The superior provenance of four-year-old Pinus densata was selected by the determination of biomass and growth characteristics at seedling stage. Different biomass models for components were constructed and then the optimum estimating models were selected based on the fitting analysis. The results showed that there were extremely significant difference among different provenances of P. densata for all growth and biomass characteristics except the primary lateral root number and lateral branch number. Aboveground biomass was the main part of the seedling biomass,accounting for about 75.60%,and aboveground biomass and total biomass was in the order:provenance Palung Zangbo > Baoxing > Niyang valley > Kangding > Chayu > Yarlung Zangbo > Lixian > Maerkang. There were different biomass distributions in various organs. The moisture content of all components were extremely significanly different among provenances of P. densata except the moisture content of branch. The correlation was more close between the ground line diameter and all components biomasses than the correlation with seedling height’s. The optimal model was the power function biomass model,in which the independent variables were the ground line diameter (D),the ground line diameter and seedling height (D2H) of all components biomass. Comprehensive comparison showed that Palung Zangbo provenance had higher seedling quality than others,which accumulated more dry matter to adapt to the environment of testing location,and had the great ability in carbon storage and carbon sequestration capacity.
biomass;growth traits;model;Pinus densata;natural hybrid species;heterosis;moisture content
S757.7
A
1004-874X(2017)02-0068-08
2016-12-05
国家自然科学基金(31360189,31070591)
周丽(1990-),女,硕士,E-mail:zhouli901009@163.com
蔡年辉(1975-),男,博士,讲师,E-mail:cainianhui@sohu.com
周丽,陈诗,朱存福,等.高山松不同种源苗木在云南松生境下的生物量与生长分析[J].广东农业科学,2017,44 (2):68-75.
