Kelch样环氧氯丙烷相关蛋白-核因子E2相关因子2-抗氧化反应元件信号通路在氧化应激中的作用及其调控剂
2017-04-24马燕芬吴志红
马燕芬 吴志红,2 赵 磊,2 高 民*
(1.内蒙古农牧业科学院动物营养与饲料研究所,呼和浩特 010031;2.内蒙古农业大学动物科学学院,呼和浩特 010018)
Kelch样环氧氯丙烷相关蛋白-核因子E2相关因子2-抗氧化反应元件信号通路在氧化应激中的作用及其调控剂
马燕芬1吴志红1,2赵 磊1,2高 民1*
(1.内蒙古农牧业科学院动物营养与饲料研究所,呼和浩特 010031;2.内蒙古农业大学动物科学学院,呼和浩特 010018)
Kelch样环氧氯丙烷相关蛋白-核因子E2相关因子2-抗氧化反应元件(Keap1-Nrf2-ARE)信号通路是机体细胞抵制氧化应激损伤和异生物质损伤最为重要的一种防御机制,且该通路与炎性疾病包括癌症、神经变性疾病、心血管疾病、衰老等密切相关。Nrf2信号的激活可诱导与ARE相关基因的各种解毒酶、抗氧化防御酶和抗氧化蛋白酶的表达的转录调控,且调控Keap1-Nrf2-ARE信号通路已成为预防和治疗氧化应激相关疾病和炎性疾病的一个强有力的靶目标。该文重点综述了氧化应激、Keap1-Nrf2-ARE信号通路在抗氧化应激中的作用及相关的调控剂。
氧化应激;Keap1-Nrf2-ARE信号通路;调控剂
机体经常暴露于各种氧化以及来自内源和外源的电化学物质而导致机体产生不同程度的氧化应激。一些活性氧(ROS)和活性氮在控制机体细胞氧化还原平衡中充当重要信使,持续的氧化应激通过破坏细胞包括脂质、蛋白质和核酸的结构,而使细胞产生不同程度的氧化损伤。此外,长期的氧化损伤还可导致机体慢性炎症并转为癌症、心血管疾病、神经变性疾病、炎性疾病和衰老。已产生氧化应激的细胞主要通过物理措施、预防、修复和抗氧化防御机制相结合的方式来抵御机体产生的各种氧化损伤。抗氧化防御系统是机体内一种主要的保护机制,细胞主要依靠抗氧化剂来中和氧化剂和电化学物质的破坏作用。抗氧化剂根据来源、性质和作用机制分为内源性(代谢和酶抗氧化剂)和外源性,或者直接抗氧化剂、间接抗氧化剂和多功能抗氧化剂[1]。直接抗氧化剂具有氧化还原活性、生命周期较短且在整个抗氧化过程中可被消耗掉,需要再生以提供进一步保护。而间接抗氧化剂可具有或不具有氧化还原活性,通过上调各种细胞保护蛋白,诸如还原型辅酶[NAD(P)H]、NAD(P)H醌氧化还原酶-1(NQO1)、超氧化物歧化酶(SOD)、谷胱甘肽巯基转移酶(GST)、谷胱甘肽过氧化物酶(GSH-Px)、血红素氧合酶-1(HO-1)、谷氨酸-半胱氨酸连接酶(GCL)、过氧化氢酶(CAT)和硫氧还蛋白(Prxl-Trx)而达到抗氧化效果[2]。这些细胞保护蛋白称为“终极抗氧化剂”,因为它们有比较长的半衰期,在他们的抗氧化作用过程中并不被消耗掉,还可以催化各种化学解毒反应,并参与一些直接抗氧化剂的再生[3]。研究表明核因子E2相关因子2(nuclear factor-E2-related factor 2,Nrf2)、胞浆蛋白Kelch样环氧氯丙烷相关蛋白1(Kelch-like epichlorohydrin-associated protein 1,Keap1)和抗氧化反应元件(antioxidant response element,ARE)是调控抗氧化反应的3种主要的细胞成分[2]。Keap1-Nrf2-ARE是一种主要的信号通路,该通路不仅可在转录水平上调控细胞保护蛋白,还可激活很多其他信号通路用于维持细胞氧化还原平衡和代谢平衡。Keap1-Nrf2-ARE信号通路可诱导氧化应激的适应性应答,否则可导致机体产生许多炎性疾病,包括癌症、老年痴呆症、帕金森氏病和糖尿病等[4]。因此,Keap1-Nrf2-ARE信号通路是目前被认为是最为引人注目的发现,已知它是预防和治疗机体产生氧化应激和炎症的一条重要途径,被称为抗氧化炎症调控剂,用于治疗氧化应激和炎症等疾病[5]。本文主要综述了机体产生的氧化应激、Keap1-Nrf2-ARE信号通路在抗氧化应激中的作用及Keap1-Nrf2-ARE信号通路的调控剂,可为如何通过调控Keap1-Nrf2-ARE信号通路达到预防和治疗机体产生氧化应激方面提供理论参考。
1 氧化应激
在正常生理情况下,机体将氧化-抗氧化状态维持在一定的动态平衡状态,即使机体内环境达到一种稳态。但当机体受到应激源刺激时,机体代谢就会出现紊乱而产生大量的ROS,氧化物和抗氧化物之间的动态平衡被打破,此时机体就会处于氧化应激状态。氧化应激的产生既有内部因素也有外部因素。外因主要包括接触环境污染、石化制品或重金属;内因包括慢性感染或急性感染。此外,机体血糖浓度的变化也会导致机体产生氧化应激。氧化应激严重威胁着畜禽的健康生长,提升养殖动物患病几率,降低生产性能和经济效益,氧化应激还可导致人类和动物多种疾病的发生发展,甚至危及生命。有关氧化应激、氧化应激产生的原因及氧化应激对动物机体造成的损害已在马燕芬等[6]发表的文章中进行了详细的综述。因此,找寻降低机体产生氧化应激的方法并开发相应的调控剂已成为当前动物营养领域预防和治疗畜禽养殖业由于氧化应激而导致的疾病的一个亟待解决的问题。
2 Keap1-Nrf2-ARE信号通路在氧化应激中的作用
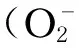
目前有关Keap1-Nrf2-ARE通路与氧化应激导致的相关疾病发病机制的研究逐渐增多。而Keap1-Nrf2-ARE通路之所以重要是因为其激活后能够启动下游多种保护性基因的表达,主要包括抗氧化酶类、解毒酶类、分子伴侣类以及抗炎因子类基因等。该信号通路中起关键作用的是Nrf2,Nrf2不仅是Keap1-Nrf2-ARE信号通路中的关键因子,还是细胞抗氧化防御机制的中枢调节者。Nrf2不仅可转录激活抗氧化酶和诱导Ⅱ相解毒酶的表达,诸如HO-1、NQO1、SOD、GST、γ-半胱氨酸连接酶(γ-GCL)[8],还与许多其他细胞作用过程密切相关,诸如分化、增殖、细胞生长、凋亡以及与不同细胞内调控蛋白密切作用的造血功能等。Nrf2还是维持细胞内氧化还原平衡的重要保障,能够降低细胞对死亡信号的敏感性,在抵抗外源性或内源性氧化应激的过程中起至关重要的作用。此外,Nrf2还能调控各种药物代谢酶、转运蛋白和细胞减少等价物(谷胱甘肽和还原型辅酶Ⅱ)等[9]。但若Nrf2缺失或存在激活障碍,则会进一步加重细胞的氧化性损伤和炎性损伤,导致细胞生理功能障碍,最后甚至导致细胞凋亡或坏死。此外,还可导致与氧化应激和炎症相关的疾病的发生,包括癌症、阿尔茨海默氏病、帕金森病、慢性阻塞性肺病、慢性阻塞性肺病、哮喘、动脉粥样硬化、糖尿病、多发性硬化、骨关节炎和类风湿性关节炎等[10-11]。研究表明,Nrf2基因普遍表达于机体内各种细胞中,主要部位为肝脏、肾脏或者与外界环境接触相对较多的组织如皮肤、肺、消化道等[12]。
机体在正常生理状态下,大部分Nrf2以抑制状态与Keap1偶联存在于细胞质中。当细胞在受到应激源或其他氧化应激刺激时,Nrf2与Keapl解偶联转运到细胞核内与ARE结合,并诱导下游编码各类抗氧化蛋白酶和Ⅱ相解毒酶的基因表达,促使抗氧化酶基因转录,从而增强细胞的抗氧化防御能力,达到保护细胞免受氧化损伤的目的。氧化还原平衡恢复之后,Nrf2核转录因子自动与ARE序列发生解离,重新回到细胞质中,Keap1-Nrf2-ARE信号通路关闭,Nrf2重新维持较低水平。如果Keap1-Nrf2-ARE信号通路关闭或者失活则可导致活性氧自由基在体内的积累,并产生多种与氧化应激和炎症相关的疾病。若Keap1-Nrf2-ARE信号通路过度活化在某些条件下也可能是有害的。例如,增加的Nrf2活性通过保护癌细胞而促进肿瘤的形成。因此,正确调节Keap1-Nrf2-ARE通路就可以保证细胞内氧化还原达到平衡状态,进而达到实现预防上述疾病发生的目的。因此,深入研究Keap1-Nrf2-ARE信号通路对于人类和动物生产中由氧化应激导致的疾病的预防和治疗具有重要意义。此外,开发相应的调控Keap1-Nrf2-ARE信号通路的调控剂对于由氧化应激介导的疾病的防治也具有重要意义。
3 Keap1-Nrf2-ARE信号通路的调控剂
越来越多的证据表明,通过调控细胞内信号激酶通路可调控化学预防调控剂激活的ARE相关基因。例如,通过调控细胞内丝裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)通路就可调控化学预防调控剂对ARE相关基因表达的调控,其中,MAPKs通路中的细胞外信号调节激酶(ERK)和c-Jun氨基末端激酶(JNK)是正向调控,而p38 MAPK是负向调控[13]。此外,调控磷脂酰肌醇3激酶(phosphatidylinositol 3′-kinase,PI3K)、PKR内质网激酶(PKR-like endoplasmic reticulum kinase,PERK)、蛋白激酶C(protein kinase C,PKC)、Fyn激酶和糖原合酶3β(glycogen synthase kinase-3β,GSK3β)通路也可调控化学预防调控剂对ARE相关基因表达的调控[14]。但是,除了PKCδ和Fyn激酶外,有关Nrf2磷酸化后如何调控ARE相关基因表达的详细分子机制在很大程度上不是很清楚。PKCδ通路激活后可导致Nrf2的Ser40磷酸化并有助于Nrf2与Keap1解偶联[15]。GSK3β通路激活后可直接磷酸化Fyn基因,磷酸化的Fyn基因又介导Nrf2的Tyr568基因磷酸化,进而直接抑制Nrf2的激活[16]。
除了采用调控细胞内信号激酶通路达到调控化学预防调控剂激活ARE相关基因外,还可采用激活剂和抑制剂来达到调控Keap1-Nrf2-ARE信号通路的表达的目的。现有的研究表明一种Nrf2激活剂是合成的三萜甲酯衍生物[2-cyano-3,12-dioxooleana-1,9(11)-dien-28-oic acid,CDDO-Me][17],但是从安全性方面考虑,由于CDDO-Me过量使用而造成的严重不良事件和高死亡率,CDDO-Me的临床试验于2012年10月终止。另一种Nrf2激活剂是富马酸二甲酯(dimethyl fumarate,DMF),通过采用DMF治疗原代培养的中枢神经系统细胞或者动物可提高核Nrf2和Ⅱ相解毒酶的活性,从而使动物机体免受氧化应激造成的损伤[18]。但是DMF是否具有化学预防激活作用目前还不清楚,还需要做进一步的研究。此外,其他抗氧化剂及合成药物也可作为Nrf2激活剂,诸如表没食子儿茶素没食子酸酯(epigallocatechin-3-gallate,EGCG)[19]、姜黄素[20]、吡唑[21]、胰岛素[22]、白藜芦醇[23]和莱菔硫烷[24]等也可通过激活人或者动物机体某一器官内部Keap1-Nrf2-ARE信号通路,通过显著上调Nrf2介导的靶基因的mRNA表达水平和蛋白表达水平而发挥抗氧化作用,达到降低活性氧的产生,提高细胞存活率,从而达到保护细胞免受氧化应激的损伤,治疗由氧化应激引起或介导的各种损伤和疾病。
Nrf2化学抑制剂一种是全反式维甲酸(all-trans retinoic acid,ATRA)。研究表明运用ATRA或其他维甲酸受体α(RARα)激活剂治疗后,可降低Nrf2与ARE的结合能力,并通过丁基化羟基苯甲醚(BHA)和叔丁基氢醌(tBHQ)抑制ARE基因的诱导[25]。其他Nrf2化学抑制剂主要为天然化学物质如木犀草素[26]、原花青素[27]、芹菜素[28]和白杨素[29]等,这些天然化学物质主要通过降低Nrf2的活性而抑制Nrf2-ARE通路的表达,但其具体分子机制不清楚,还需要做进一步研究。
4 小 结
Keap1-Nrf2-ARE信号通路是机体内一种重要的激活细胞适应性应答抵制氧化应激和其他类型应激的抗氧化防御机制。Keap1是Nrf2的负调控因子,Nrf2是ARE系统内的主效应因子。通过反向调控Keap1蛋白,激活的ARE系统可诱导抗氧化应答基因预防氧化损伤、炎症和肿瘤的形成。此外,掌握Keap1-Nrf2-ARE通路的药理活性可为开发新型抗氧化剂、抗炎剂和抗癌剂提供新的理论基础。而来自于不同类型的天然产物和合成材料中的调控剂都可作为Keap1-Nrf2-ARE信号通路的激活剂或者抑制剂。因此,通过运用天然植物制剂或者药物制剂调控Keap1-Nrf2-ARE信号通路可为由于氧化应激而导致的疾病提供新的防治策略。
[1] LYAKHOVICH V V,VAVILIN V A,ZENKOV N K,et al.Active defense under oxidative stress.The antioxidant responsive element[J].Biochemistry,2006,71(9):962-974.
[2] DINKOVA-KOSTOVA A T,TALALAY P.Direct and indirect antioxidant properties of inducers of cytoprotective proteins[J].Molecular Nutrition & Food Research,2008,52(Suppl.1):S128-S138.
[3] HOLTZCLAW W D,DINKOVA-KOSTOVA A T,TALALAY P.Protection against electrophile and oxidative stress by induction of phase 2 genes:the quest for the elusive sensor that responds to inducers[J].Advances in Enzyme Regulation,2004,44(1):335-367.
[4] STEWART J D,HENGSTLER J G,BOLT H M.Control of oxidative stress by the Keap1-Nrf2 pathway[J].Archives of Toxicology,2011,85:239.
[5] TAGUCHI K,MOTOHASHI H,YAMAMOTO M.Molecular mechanisms of the Keap1-Nrf2 pathway in stress response and cancer evolution[J].Genes to Cells,2011,16(2):123-140.
[6] 马燕芬,宋利文,高民,等.氧化应激对围产期奶牛乳房炎的影响及其调控机制[J].动物营养学报,2015,27(3):671-676.
[7] SURH Y J,KUNDU J K,NA H K.Nrf2 as a master redox switch in turning on the cellular signaling involved in the induction of cytoprotective genes by some chemopreventive phytochemicals[J].Planta Medica,2008,74(13):1526-1539.
[8] KUNDU J K,SURH Y J.Inflammation:gearing the journey to cancer[J].Mutation Research/Reviews in Mutation Research,2008,659(1/2):15-30.
[9] MA Q.Transcriptional responses to oxidative stress:pathological and toxicological implications[J].Pharmacology & Therapeutics,2010,125(3):376-393.
[10] COPPLE I M.The Keap1-Nrf2 cell defense pathway—a promising therapeutic target?[J].Advances in Pharmacology,2012,63:43-79.
[11] MAGESH S,CHEN Y,HU L Q.Small molecule modulators of Keap1-Nrf2-ARE pathway as potential preventive and therapeutic agents[J].Medicinal Research Reviews,2012,32(4):687-726.
[12] CHO H Y,REDDY S P,KLEEBERGER S R.Nrf2 defends the lung from oxidative stress[J].Antioxidants & Redox Signaling,2006,8(1/2):76-87.
[13] KEUM Y S,JEONG W S,KONG A N T.Chemoprevention by isothiocyanates and their underlying molecular signaling mechanisms[J].Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis,2004,555(1/2):191-202.
[14] KEUM Y S.Regulation of Nrf2-mediated phase Ⅱ detoxification and anti-oxidant genes[J].Biomolecules and Therapeutics (Seoul),2012,20(2):144-151.
[15] NITURE S K,JAIN A K,JAISWAL A K.Antioxidant-induced modification of INrf2 cysteine 151 and PKC-δ-mediated phosphorylation of Nrf2 serine 40 are both required for stabilization and nuclear translocation of Nrf2 and increased drug resistance[J].Journal of Cell Science,2009,122(24):4452-4464.
[16] JAIN A K,JAISWAL A K.Phosphorylation of tyrosine 568 controls nuclear export of Nrf2[J].The Journal of Biological Chemistry,2006,281(17):12132-12142.
[17] LIBY K T,SPORN M B.Synthetic oleanane triterpenoids:multifunctional drugs with a broad range of applications for prevention and treatment of chronic disease[J].Pharmacological Reviews,2012,64(4):972-1003.
[18] SCANNEVIN R H,CHOLLATE S,JUNG M Y,et al.Fumarates promote cytoprotection of central nervous system cells against oxidative stress via the nuclear factor (erythroid-derived 2)-like 2 pathway[J].The Journal of Pharmacology and Experimental Therapeutics,2012,341(1):274-284.
[19] 杨光照.EGCG通过上调Nrf2/HO-1途径降低PM2.5所致HUVEC氧化损伤的机制研究[D].博士学位论文.太原:山西医科大学,2015.
[20] FAROMBI E O,SHROTRIYA S,NA H K,et al.Curcumin attenuates dimethylnitrosamine-induced liver injury in rats through Nrf2-mediated induction of heme oxygenase-1[J].Food and Chemical Toxicology,2008,46(4):1279-1287.
[21] ZHONG J L,EDWARDS G P,RAVAL C,et al.The role of Nrf2 in ultraviolet a mediated heme oxygenase 1 induction in human skin fibroblasts[J].Photochemical & Photobiological Science,2010,9(1):18-24.
[22] OKOUCHI M,OKAYAMA N,ALEXANDER J S,et al.NRF2-dependent glutamate-L-cysteine ligase catalytic subunit expression mediates insulin protection against hyperglycemia-induced brain endothelial cell apoptosis[J].Current Neurovascular Research,2006,3(4):249-261.
[23] 蒋怡芳.白藜芦醇对过氧化氢诱导的原代脊髓星形胶质细胞毒性的保护作用[D].硕士学位论文.石家庄:河北医科大学,2009.
[24] 鲍兵.莱菔硫烷激活Nrf2-ARE通路拮抗PC12细胞氧化应激损伤的机制研究[D].硕士学位论文.南昌:南昌大学,2013.
[25] WANG X J,HAYES J D,HENDERSON C J,et al.Identification of retinoic acid as an inhibitor of transcription factor Nrf2 through activation of retinoic acid receptor alpha[J].Proceedings of the National Academy of Sciences of the United States of America,2007,104(49):19589-19594.
[26] TANG X W,WANG H Y,FAN L F,et al.Luteolin inhibits Nrf2 leading to negative regulation of the Nrf2/ARE pathway and sensitization of human lung carcinoma A549 cells to therapeutic drugs[J].Free Radical Biology and Medicine,2011,50(11):1599-1609.
[27] OHNUMA T,MATSUMOTO T,ITOI A,et al.Enhanced sensitivity of A549 cells to the cytotoxic action of anticancer drugs via suppression of Nrf2 by procyanidins from cinnamomi cortex extract[J].Biochemical and Biophysical Research Communications,2011,413(4):623-629.
[28] GAO A M,KE Z P,WANG J N,et al.Apigenin sensitizes doxorubicin-resistant hepatocellular carcinoma BEL-7402/ADM cells to doxorubicin via inhibiting PI3K/Akt/Nrf2 pathway[J].Carcinogenesis,2013,34(8):1806-1814.
[29] GAO A M,KE Z P,SHI F,et al.Chrysin enhances sensitivity of BEL-7402/ADM cells to doxorubicin by suppressing PI3K/Akt/Nrf2 and ERK/Nrf2 pathway[J].Chemico-Biological Interactions,2013,206(1):100-108.
*Corresponding author, professor, E-mail: gmyh1588@126.com
(责任编辑 田艳明)
Role of Kelch-Like Epichlorohydrin-Associated Protein 1-Nuclear Factor-E2-Related Factor 2-Antioxidant Response Element Signaling Pathway in Anti-Oxidative Stress and Its Regulators
MA Yanfen1WU Zhihong1,2ZHAO Lei1,2GAO Min1*
(1.AnimalNutritionandFeedInstitute,AgriculturalandAnimalHusbandryAcademyofInnerMongolia,Hohhot010031,China; 2.CollegeofAnimalScience,InnerMongoliaAgriculturalUniversity,Hohhot010018,China)
Kelch-like epichlorohydrin-associated protein 1-nuclear factor-E2-related factor 2-antioxidant response element (Keap1-Nrf2-ARE) signaling pathway is one of the most important cellular defense mechanisms against oxidative stress and xenobiotic damage, and it is closely associated with inflammatory diseases, including cancer, neurodegenerative diseases, cardiovascular diseases, aging and so on. Activation of Nrf2 signaling induces the transcriptional regulation of ARE-dependent expression of various detoxifying and antioxidant defense enzymes and proteins. Keap1-Nrf2-ARE signaling pathway has become an attractive target for the prevention and treatment of oxidative stress-related diseases and inflammatory diseases. Oxidative stress, the role of Keap1-Nrf2-ARE signaling pathway in anti-oxidative stress and the regulators of Keap1-Nrf2-ARE signaling pathway were reviewed in this paper.[ChineseJournalofAnimalNutrition, 2017, 29(4):1091-1095]
oxidative stress; Keap1-Nrf2-ARE signaling pathway; regulators
10.3969/j.issn.1006-267x.2017.04.001
2016-10-13
国家自然科学基金(30460616,31601975);内蒙古自然科学基金(2014BS0348);内蒙古农牧业科学院青年创新基金(2015QNJJM07);国家现代农业(奶牛)产业技术体系项目(CARS-37)
马燕芬(1979—),女,内蒙古呼和浩特人,研究员,博士,主要从事反刍动物营养调控理论与技术研究。E-mail: ma2999@163.com
*通信作者:高 民,研究员,硕士生导师,E-mail: gmyh1588@126.com
S811
A
1006-267X(2017)04-1091-05
