利用SSR标记分析金观音(半)同胞茶树品种遗传差异
2017-04-22王让剑杨军孔祥瑞高香凤
王让剑,杨军,孔祥瑞,高香凤
福建省农业科学院茶叶研究所 国家茶树改良中心福建分中心,福建 福安 355015
利用SSR标记分析金观音(半)同胞茶树品种遗传差异
王让剑,杨军,孔祥瑞,高香凤
福建省农业科学院茶叶研究所 国家茶树改良中心福建分中心,福建 福安 355015
准确鉴定无性系茶树品种的遗传差异是对其进行保护与利用的重要前提。实验对金观音同胞及半同胞茶树品种的遗传差异进行了分析,结果表明,参试的31个SSR标记分型结果具有高度的稳定性。共扩增出117个等位位点,单个标记为2.0~8.0个,平均 3.77个。基因型数为 2.0~11.0个,平均5.16个。基因多样性指数范围为0.15~0.80,平均0.54。基因杂合度范围为0.17~0.94,平均0.61。引物多态信息含量范围为0.14~0.77,平均0.48。引物鉴别力范围为0.07~0.73,平均0.29。参试品种两两之间的遗传距离变化范围为0.10~0.52,平均0.35。遗传多样性大小顺序为:同胞茶树品种<半同胞茶树品种<非同胞茶树品种。利用系统发育树可将参试茶树品种分为5类,与茶树品种的叶色、制茶特征分类结果基本一致。任意两个品种同时在6个核心引物处(基因座)的基因型存在相同的可能性很低,为3.85×10-5,这组核心引物具有较高的鉴别力,可将参试茶树品种完全鉴定。
茶树;SSR;无性系品种;遗传差异;指纹图谱
茶树品种是茶叶生产最基本、最重要的生产资料,也是茶产业可持续发展的基础。铁观音与黄棪是我国著名的两个茶树品种,课题组前期从铁观音与黄棪杂交后代中选育出超亲优势或杂种优势强,融有两亲本优异品质性状于一体,香气、制优率、产量、抗性与适应性等性状超过两亲本的金观音、金牡丹、黄玫瑰等品种,以及金玫瑰、春桃香、紫观音等品系[1-3]。利用该系列茶树品种(系)制成的乌龙茶、红茶均曾获得“中茶杯”、“国饮杯”或“闽茶杯”的特等奖或金奖,其中金牡丹、黄玫瑰更被评为农业部“九五”国家重点科技攻关计划科技成果一级优异种质。
在众多DNA分子标记中,SSR由于具有共显性、位点丰富、多态性好、稳定性高等诸多优点而备受青睐[4-5]。茶树遗传多样性分析[6-8]、品种真实性鉴定[9-11]、遗传图谱构建[12-13]等方面均有 SSR分子标记的广泛应用。福建是我国茶叶主产区之一,无性系茶树品种推广率高居全国第一,但近年随着无性系茶树品种的持续大面积推广,品种混杂、近似品种难以辨认等现象逐渐突出。茶树是异花授粉的多年生作物,遗传基础非常复杂,育种年限长达20年以上,品种选育是一项非常艰苦的工作。为保障生产用种与保护育种者知识产权,在分子水平上对重点推广的茶树品种进行遗传差异分析与真实性鉴定具有重要的现实意义。金观音同胞及半同胞茶树品种适制性广,制优率高,制茶花香明显,品质优异,倍受茶叶生产者、经营者和消费者的欢迎,本研究利用SSR分子标记,开展金观音同胞及半同胞茶树品种遗传差异分析与真实性鉴定,以期为该系列茶树品种的保护与利用提供分子水平依据。
1 材料与方法
1.1 供试材料
参试材料为金观音同胞与半同胞茶树品种(系),以及铁观音、黄棪、本山3个对照种,如表1所示。相同材料不同单株重复取样1次,样品采集后用液氮迅速冷冻处理,保存在-70℃冰箱中备用。
1.2 实验方法
1.2.1 茶树基因组DNA的提取

表1 参试材料Table 1 The tested materials
采用 CTAB法[14]按照生物学重复提取茶树基因组DNA。用1.0%的琼脂糖凝胶电泳进行茶树基因组分子量大小检测,利用Biospec-nano(230 V)核酸蛋白分析仪进行定量,检测DNA的纯度与浓度。
1.2.2 引物合成
按照均匀分布的原则,在茶树遗传连锁图谱[12]上随机选取45个SSR标记,以表1中的铁观音与黄棪人工杂交后代茶树品种DNA为模板进行初筛,去除无多态性的引物14对(未列),剩余31对标记引物合成荧光引物,如表2所示。1.2.3 PCR扩增与产物检测
反应体系:ddH2O 16 μL,10×Buffer 3 μL,dNTP(10 mmol·L-1)3 μL,MgCl22 μL,上、下游引物(10 μmol·L-1)各1.5 μL,Taq酶1 μL(0.5 U),模板DNA 2 μL。反应程序:94℃预变性5 min,使模板DNA充分变性,然后进入下列温度循环:94℃变性1 min,不同温度条件退火1 min,72℃延伸1 min,重复35个热循环;72℃延伸20 min,最后4℃保存。在96孔板内加入1 μL DNA扩增样品与10 μL ROX500分子量标准工作液,震荡混匀,使用 Applied Biosystems 3730xlDNA分析仪检测PCR扩增产物。PCR扩增、产物检测均重复2次。

表2 参试SSR分子标记Table 2 SSR markers selected from 15 linkage groups of tea plant
1.2.4 数据统计与分析
使用PowerMarker软件(v3.25)计算31对参试引物的遗传参数,包括等位位点频率、等位位点数、基因型数、观测基因杂合度、引物多态性息含量,以及参试材料间的Nei′s遗传距离。利用 MEGA软件(V4.0)绘制参试茶树品种系统发育树。根据等位位点的频率,计算各引物的PID值(Probability of identity, PID),PID值表示两个随机个体拥有相同基因型的概率,可以反映引物的鉴别力,PID值越大说明引物的鉴别力越小,PID值越小则说明引物的鉴别力越大,计算公式为:
PID=2(∑Pi2)2-∑Pi4,其中 Pi表示某个基因座第i个等位位点的频率[15]。
1.2.5 核心引物筛选
为快速鉴别参试茶树品种,在31对引物中筛选出 1组核心引物,筛选标准如下:①选择长基序(Motif)引物,碱基数≥4;②引物多态信息含量丰富,PIC值>0.5;③引物鉴别力较强,PID值<参试的31对引物PID的平均值。

表3 黄棪品种重复实验结果Table 3 The two independent detection results of Huang Dan
2 结果与分析
2.1 数据稳定性分析
[16]的方法,对检测结果的原始数据进行标准化处理。由表3可知,标准化处理结果重复之间无差异,说明 SSR分型结果具有高度的稳定性。
2.2 引物多态性分析
参试引物的各遗传参数计算结果如表 4所示。31对引物共扩增出 117个等位位点,最多的为 8.00个(TM262),最少的为 2.00个(TM514、TM576、TM601),平均每对引物检测到3.77个;基因型数最多的为11.0个(TM428、TM262),最少的为2.0个(TM576、TM601),平均每对引物检测到5.16个;基因多样性指数在0.15~0.80之间,平均为0.54;基因杂合度为 0.17~0.94,平均为 0.61;引物多态信息含量在0.14~0.77之间,平均为0.48;引物鉴别力在0.07~0.73之间,平均为0.29。参试品种的分型模式如图 1所示,在TM324基因座处共有 3个等位位点即:171、177、183 bp,不同品种所包含的等位位点存在一定的差异。

表4 参试31个SSR标记主要遗传参数Table 4 Key genetic statistics of the 31 tested SSR markers
2.3 遗传差异分析
如表5所示,参试茶树品种两两之间的遗传距离变化范围为0.10~0.52,平均0.35。遗传距离最近的两个品种是金观音和紫观音,最远的两个品种是春闺和铁观音,说明金观音与紫观音之间的遗传差异最小,春闺与铁观音之间的遗传差异最大。比较同胞茶树品种、半同胞茶树品种及非同胞茶树品种之间的等位位点个数、基因多样性指数、多态信息含量、遗传距离可知,以上各指标均为同胞茶树品种<半同胞茶树品种<非同胞茶树品种,说明同胞茶树品种之间的遗传多样性最小(表6)。

图1 引物TM324对参试品种的分型模式图Fig. 1 Fingerprinting mode of the tested cultivars characterized by TM324
2.4 系统发育树分析
参试材料之间的亲缘关系可以通过系统发育树直观地体现出来。系统发育树分析结果(图2)表明,18个品种可以分为5类:第1类包括铁观音、凤圆春、本山3个茶树品种,叶色均为深绿,其中凤圆春、本山所制乌龙茶品质均酷似铁观音,滋味均具有铁观音的“音韵”特点,凤圆春、本山亩产高于铁观音,比铁观音容易种植,3个品种均为晚生种;第2类包括黄玫瑰、黄观音、黄棪、春闺、黄奇5个茶树品种,叶色均为黄绿,所制乌龙茶品质均具有黄棪的“透天香”特点,春茶新梢萌发期按早晚顺序排列为:黄棪>黄观音=黄玫瑰>黄奇>春闺(“>”表示“早于”,“=”表示“相当”,下同);第3类包括紫牡丹、金牡丹、春桃香3个茶树品种,叶色为深绿或绿,这类品种制乌龙茶均具有明显的“花果香”,且制优率高,亩产均高于铁观音,春茶新梢萌发期按早晚顺序为:金牡丹>春桃香>紫牡丹;第4类包括春兰、瑞香2个品种,叶色均为黄绿,制乌龙茶具有“兰花香”,亩产高于铁观音,春茶新梢萌发期春兰早于瑞香;第5类包括金桂观音、金玫瑰、紫观音、金观音、紫玫瑰5个品种,叶色均为深绿,这类品种制乌龙茶香气馥郁悠长,“韵味”显,亩产均高于铁观音,春茶新梢萌发期按早晚顺序为:金玫瑰>金观音>紫玫瑰>金桂观音>紫观音。系统发育树分类结果与茶树品种的叶色、制茶特征分类结果基本一致。

表5 参试材料遗传距离Table 5 Genetic distance of the tested cultivars based on SSR markers
2.5 核心引物筛选与参试品种快速鉴定
按上述筛选标准,从 31对参试引物中进一步筛选出6对核心引物,如表7所示。假设每个位点都是独立遗传的,根据引物的PID值,理论上可以计算出利用6对核心引物同时进行参试茶树品种鉴定,任意两个品种的基因型存在相同的可能性为3.85×10-5,即每100 000个品种中有 3.85个品种具有相同的基因型,说明这组核心引物具有较高的鉴别力。按照核心引物 PID值由小到大的顺序排列,只需利用TM440、TM348、TM241作为首选引物组合即可将参试材料快速鉴别出来,其余3对核心引物TM324、TM569、TM461可以作为补充引物进行补充鉴定。
3 讨论
荧光标记电泳检测方法通过与内标比较,依据待检样品目标峰的位置,即可直接读出DNA片段大小,具有良好的准确性,但部分SSR标记易产生等位位点“漂移”(Allelic drift),扩增片段大小与实际等位位点大小产生±2.1 bp的波动[17],此时应对照SSR标记重复基序(motif)碱基个数对原始数据进行纠偏及标准化处理。本实验参试引物中,TM445、TM569、TM461、TM262、TM348、TM351的扩增结果均产生不同程度的“漂移”,但标准化后重复实验间无差异,确保了SSR标记分型数据的准确性。
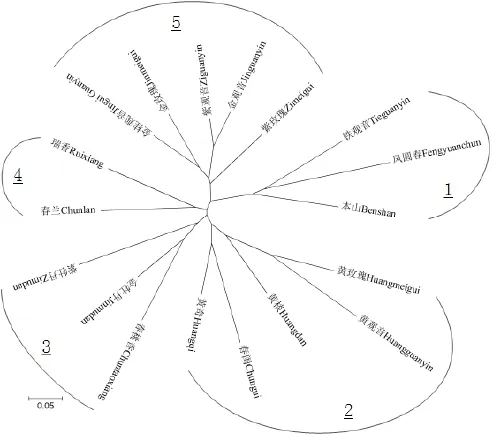
图2 参试无性系茶树品种系统发育树Fig. 2 Phylogeny tree of the tested clonal tea cultivars
利用 SSR分子标记进行品种遗传差异分析及真实性鉴定,引物的筛选是重点,本实验在引物筛选方面进行了较充分的考虑。首先,初选31个SSR引物覆盖了茶树15个连锁群,确保了遗传差异分析具有一定的代表性,从系统发育树分类结果也可以看出基于31个参试SSR标记的分类结果与茶树品种的叶色、制茶特征分类结果基本一致,推测参试 SSR标记可能与叶色和制茶特征性状具有较高的关联性;其次,核心引物均为长基序引物,基序碱基数均大于4,不受等位位点“漂移”的影响,相邻的等位位点容易鉴别,读取的电泳数据更可靠,更适宜用来进行品种的真实性鉴定;第三,核心引物PIC值均大于0.5,PID值均小于全部参试引物的平均数,具有较高的多态性和鉴别力,能够利用尽量少的引物鉴定尽量多的品种,可以降低鉴定成本和提高鉴定效率。
铁观音与黄棪品种历史悠久,享誉国内外,但由于历史原因,在苗木繁育过程中产生了人为或本身遗传变异的混杂,尤以铁观音混杂更甚,严重影响了其茶叶的品质[18]。从表6可知,铁观音与黄棪的人工杂交后代品种在TM440、TM348、TM461 3个基因座处均产生了非父母本来源的等位位点,如:TM440处的“160”与“170”、TM348处的“248”、TM461处的“204”,而铁观音的近亲本山品种恰恰在TM440基因座上拥有“160”位点,在TM348处拥有“248”位点,这说明参试的铁观音与金观音等同胞茶树品种真正的母本存在一定的差异,这也从分子水平上揭示了现存铁观音品种的混杂现象。
虽然金观音同胞茶树品种的遗传多样性受两亲本的局限,其遗传多样性低于半同胞及非同胞茶树品种,但多年的田间试验及生产实践均表明,金观音(半)同胞茶树品种香气、制优率、产量、抗性与适应性等性状超过两亲本,已融有两亲本的优异性状,推测其中可能包含大量有利的等位基因资源,因此金观音(半)同胞茶树品种除继续在生产上发挥作用,亦可作为优异茶树种质资源用于茶树品种选育。因人工选择而导致品种多样性降低,是作物品种改良中的共性问题[19-20],今后在茶树育种过程中应更加重视茶树种质资源的收集、保护、鉴定工作,加强种质资源创新,为育种不断提供新基因源,同时积极探索并开展分子标记辅助选择育种,缩短育种年限。
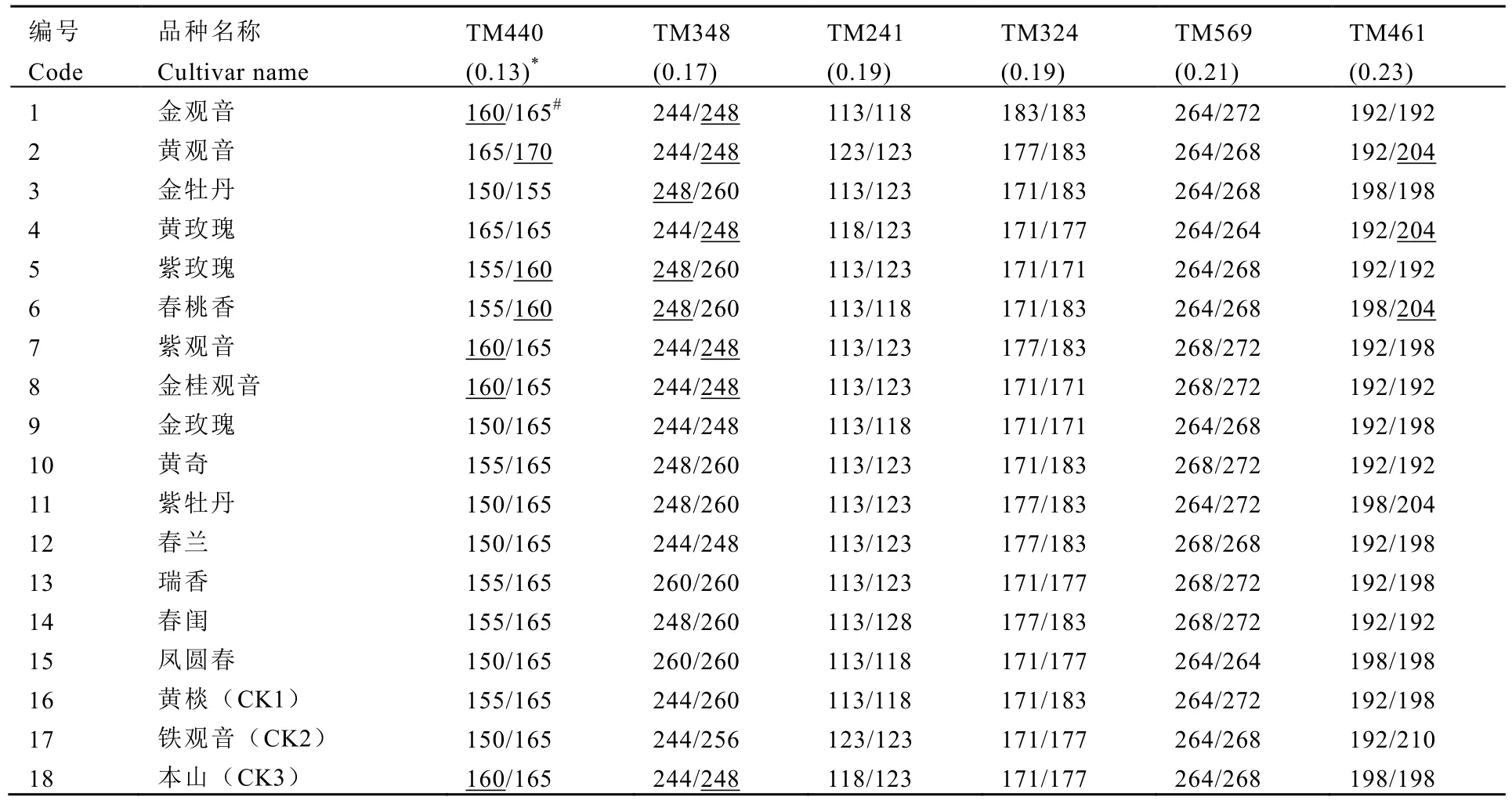
表7 参试茶树品种6对核心引物的基因型Table 7 Genotyping results of 18 cultivars by six core primer pairs
参考文献
[1]郭吉春, 叶乃兴, 何孝延, 等. 乌龙茶品系比较鉴定与选择[J]. 茶叶科学简报, 1994, 145(4): 13, 14-21.
[2]郭吉春, 杨如兴, 叶乃兴, 等. 10个茶树高优新品种的育成与应用[J]. 福建茶叶, 2003, 96(3): 19-21.
[3]郭吉春, 杨如兴, 张文锦, 等. 茶树杂交种金观音与黄观音的选育及应用[J]. 贵州科学, 2008, 26(2): 20-24.
[4]唐浩, 余汉勇, 张新民, 等. 水稻新品种测试的标准品种DNA 指纹图谱多样性分析[J]. 植物遗传资源学报, 2015, 16(1): 100-106.
[5]江锡兵, 汤丹, 龚榜初, 等. 基于 SSR标记的板栗地方品种遗传多样性与关联分析[J]. 园艺学报, 2015, 42(12): 2478-2488.
[6]YAO Mingzhe, MA Chunlei, QIAO Tingting, et al. Diversity distribution and population structure of tea germplasms inChina revealed by EST-SSR markers [J]. Tree Genetics & Genomes, 2012, 8(1): 205-220.
[7]李赛君, 雷雨, 段继华, 等. 基于 EST-SSR的祁门种群体遗传多样性和亲缘关系分析[J]. 茶叶科学, 2015, 35(4): 329-335.
[8]黄晓霞, 唐探, 姜永雷, 等. 千家寨不同海拔野生茶树的EST-SSR 遗传多样性研究[J]. 茶叶科学, 2015, 35(4): 347-353.
[9]TAN Liqiang, PENG Min, XU Liyi, et al. Fingerprinting 128 Chinese clonal tea cultivars using SSR markers provides new insights into their pedigree relationships [J]. Tree Genetics & Genomes, 2015, 11: 90. DOI: 10.1007/s11295-015-0914-6.
[10]张成才, 刘园, 姜燕华, 等. SSR标记鉴定浙江省主要无性系茶树品种的研究[J]. 植物遗传资源学报, 2014, 15(5): 926-931.
[11]黄丹娟, 马建强, 陈亮. 茶树PVP申请品种的SSR分子标记鉴定和系谱关系分析[J]. 茶叶科学, 2016, 36(1): 68-71.
[12]MA Jianqiang, YAO Mingzhe, MA Chunlei, et al. Construction of a SSR-based genetic map and identification of QTLs for catechins content in tea plant (Camellia sinensis) [J]. Plos One, 2014, 9(3): e93131.
[13]TAN Liqiang, WANG Liyuan, XU Liyi, et al. SSR-based genetic mapping and QTL analysis for timing of spring bud flush, young shoot color, and mature leaf size in tea plant (Camellia sinensis) [J]. Tree Genetics & Genomes, 2016, 12: 52. DOI: 10.1007/s11295-016-1008-9.
[14]Dellaporta SL, Wood J, Hicks JB. A plant DNA minipreparation: version II [J]. Plant Mol Biol Rep, 1983(1): 19-21.
[15]Taberlet P, Luikart G. Non-invasive genetic sampling and individual identification [J]. Biol J Linn Soc, 1999, 68(1/2): 41-55.
[16]王让剑, 杨军, 孔祥瑞, 等. 基于荧光标记SSR的CID法快速鉴定福建茶树品种[J]. 茶叶科学, 2016, 36(2): 210-218.
[17]Ramana M Idury, Lon R Cardon. A simple method for automated allele binning in microsatellite markers [J]. Genome Research, 1997, 7(11): 1104-1109.
[18]詹梓金, 李宗恒, 徐彪, 等. 保护铁观音原种保持优异品质—论铁观音品种提纯复壮[J]. 茶叶科学技术, 2006, 191(2): 20.
[19]杨玉蓉, 孙建昌, 王兴盛, 等. 宁夏不同年代水稻品种的遗传多样性比较[J]. 植物遗传资源学报, 2014, 15(3): 457-464.
[20]赵檀, 金柳艳, 李远, 等. 基于全基因组的河北省小麦品种遗传多样性分析[J]. 植物遗传资源学报, 2015, 16(1): 45-53.
Genetic Analysis of Full- and Half-sib Families of Tea Cultivar Jinguanyin Based on SSR Molecular Markers
WANG Rangjian, YANG Jun, KONG Xiangrui, GAO Xiangfeng
Tea Research Institute, Fujian Academy of Agricultural Science, Fujian Branch, National Center for Tea Improvement, Fu′an 355015, China
Accurate identification of the genetic differences among tea cultivars is the prerequisite for conservation and utilization. Genetic differences of the full- and half-sib families of tea cultivar Jinguanyin were analyzed based on SSR molecular markers. The results showed that the genotyping data of the tested 31 SSR markers were highly stable, with totally 117 alleles and averagely 3.77 alleles per marker (ranging from 2 to 8). The number of genotypes, gene diversity index, gene heterozygosity, polymorphism information content (PIC) and discrimination power (PID), genetic distance ranged from 2 to 11, 0.15 to 0.8, 0.17 to 0.94, 0.14 to 0.77, 0.07 to 0.73 and 0.10 to 0.52, averagely 5.16, 0.54, 0.61, 0.48, 0.29 and 0.35, respectively. The genetic diversity followed the order that full-sib tea cultivars< half-sib tea cultivars < non-sib tea cultivars. The tested tea cultivars could be divided into 5 categories by the phylogenetic tree, which was closely correlated with leaf colour and tea manufacture characteristics. It’s a low probability (3.85×10−5) to identify the wrong cultivar using 6 core SSR markers, which were highly accurate in identification and could be used for fingerprinting analysis in the tested clonal tea cultivars.
tea [Camellia sinensis (L.) O. Kuntze], SSR, clonal tea cultivar, genetic differences, fingerprinting
S571.1;Q52
A
1000-369X(2017)02-139-10
2016-09-29
2016-12-05
福建省自然科学基金(2015J01098)、福建省省属公益类科研院所基本科研专项(2014R1012-3、2014R1012-6)
王让剑,男,助理研究员,主要从事茶树种质资源与遗传育种研究,E-mail: rangjian.wang@163.com
