南沙湿地公园红树林物种多样性与空间分布格局
2017-04-01邱霓徐颂军邱彭华宋焱牛安逸许观嫦
邱霓,徐颂军,邱彭华,宋焱,牛安逸,许观嫦
1. 华南师范大学地理科学学院,广东 广州 510631;2. 广州大学地理科学学院,广东 广州 510006;3. 海南师范大学地理与旅游学院,海南 海口 571158
南沙湿地公园红树林物种多样性与空间分布格局
邱霓1,2,徐颂军1*,邱彭华3,宋焱1,牛安逸1,许观嫦1
1. 华南师范大学地理科学学院,广东 广州 510631;2. 广州大学地理科学学院,广东 广州 510006;3. 海南师范大学地理与旅游学院,海南 海口 571158
红树林是海岸的特有植被类型,研究其物种多样性与空间分布,对红树林的恢复与保护具有重要意义。本文以南沙湿地公园红树林为典型案例,采用遥感影像解译和样方调查的方法,对红树林物种多样性、群落特征与空间分布格局进行研究。结果表明:(1)南沙湿地公园红树林物种多样性指数高于其他与之纬度相近的红树林群落;(2)以无瓣海桑Sonneratia apetala Buch-Ham.、桐花树Aegiceras corniculatum (L.) Blanco.、黄槿Hibiscus tiliaceus L.和芦苇Phragmites australis (Cav.) Trin. ex Steud.为优势种;(3)主要的群落类型为无瓣海桑群落、黄槿群落和芦苇群落等3种,总面积百分比达86.72%,无瓣海桑群落和黄槿群落面积占比分别为41.13%和36.44%,成为红树林的绝对优势群落;(4)红树林群落在空间上呈聚集态分布,聚集度最高的是无瓣海桑群落,其次是黄槿群落,面积占比最低的是木榄群落;(5)引进种无瓣海桑,发展态势好,抑制了乡土种红树植物生长。建议扩种白骨壤Avicennia marina (Forsk.) Vierh.、桐花树和秋茄Kandelia candel (L.) Druce.,扩大其斑块面积,以提升乡土红树植物群落的稳定性,达到有效保护红树林的目的。
红树林;景观格局;空间分布;物种多样性;南沙湿地公园
红树林是热带、亚热带海岸潮间带湿地生态系统中独特的植物群落(彭友贵等,2005),是海岸特有的植被类型。全世界现有真红树植物11科70种(Wang et al.,2003;李皓宇等,2016),中国有11科26种(廖宝文等,2014)。近60年来,中国红树林面积减少了约1/2(廖宝文等,2014),而红树林面积全国之最的广东省,尤其是珠江口海区,红树林面积减少了3/4(常永智等,2013)。由于围海造田、养殖,城市化扩张等因素导致红树林大面积消失,红树林亟待保护。近年来,中国实施红树林生态恢复和修复工程,广东红树林面积增长较快(但新球等,2016),而广州市的南沙红树林以人工种植为主,对其进行研究具有一定的现实意义,可为其他红树林区开展红树林生态恢复提供借鉴。
多数研究通过调查红树林种类组成信息分析物种多样性,如李皓宇等(2016)调查了粤东沿海红树林物种组成;廖宝文等(2014)对中国东南沿海红树林的种类组成、空间分布、面积变化进行研究;张伟等(2010)对湛江红树林19年的面积变化进行研究。红树林野外调查工作量大,且所能达到的时空尺度有限(李文华,2011),遥感技术成为了监测红树林动态变化的重要手段(Seto et al.,2007;Mas,1999;Simard et al.,2002;Giri et al.,2007),如黎夏等(2006)采用遥感方法,对珠江口红树林时空动态变化进行分析。以上对红树林的研究侧重于面积和种类组成方面,但仍存在群落的空间分布不清晰等问题。
2012年9月6日国务院正式批准成立南沙新区,成为全国第6个国家级新区。南沙新区湿地面积占土地总面积的89.2%(徐越等,2013),根据规划,到2025年南沙将初步建成岭南生态水乡之都,而红树林是其重要的湿地生态系统和独具特色的生态景观。南沙红树林主要集中在十九围的南沙湿地公园,该公园是广州市最大的湿地公园,也是广州地区候鸟最丰富、最集中的地区,更是中国候鸟迁徙的三大路径之一(常弘等,2009)。南沙红树林的重要性显著,许多专家学者对该区进行了研究,如张晓君等(2014)对南沙人工红树植物海桑属Sonneratia群落特征及其土壤特性的动态变化进行研究,缪绅裕等(2011)对南沙3种红树植物的生理生态特征进行比较,彭友贵等(2005,2004)和李玫等(2009)对南沙湿地生态系统的特征、服务功能和生物多样性进行研究,朱可峰等(2011)对南沙人工红树林凋落物组成与季节变化进行研究。
在广州市“南拓”的战略指导下,南沙开展了红树林恢复行动,1998年开始种植人工红树林,丰富了红树植物的物种多样性。然而,目前有关南沙红树林物种多样性和群落空间分布的研究比较少。本文在实地调研的基础上,采用高分辨率遥感影像,提取红树林信息,对南沙红树林群落空间分布进行量化研究,旨在推动实施红树林的恢复行动,为未来南沙红树林引种造林和修复工作提供信息,为保护沿海湿地红树林物种多样性、维护生态安全和营造健康生态系统提供支撑。
1 研究区概况
南沙湿地是珠江三角洲经济区的几何中心,研究区南沙湿地公园位于广州市南部万顷沙镇南端的十九围(东经113°37′10″~113°38′52″,北纬22°35′5″~22°37′28″),介于十八涌和十九涌之间,濒临伶仃洋,地处珠江出海口,咸淡水交界处(图1)。该区在原滨海滩涂上围筑而成,面积6.5421 km2,咸度1‰~8‰。属南亚热带海洋性季风气候,夏无酷暑,冬无严寒,年均气温21.8 ℃,年均降水量1635.6 mm(邱彭华等,2011)。
2 数据获取与方法
2.1 遥感数据
选用法国SPOT IMAGE公司的Pleiades高分辨率多光谱的卫星遥感影像(2012-09-22),0.5 m全色+2 m多光谱数据,辅之以南沙区划图,作为研究数据。图像处理软件平台主要为ENVI 5.1和ArcMap 10.2.1;景观指数计算采用Fragstats 4.2。
参照相关研究成果(彭友贵等,2004;李玫等,2009;朱可峰等,2011;邱彭华等,2011),结合野外调查数据,对红树林群落信息进行提取。将南沙湿地公园红树林群落分为无瓣海桑Sonneratia apetala、桐花树Aegiceras corniculatum、秋茄Kandelia candel (L.) Druce、拉关木Laguncularia racemosa Gaertn. f.、木榄Bruguiera gymnorrhiza (L.) Lam、黄槿Hibiscus tiliaceus L.、水黄皮Pongamia pinnata (L.) Pierre、海芒果Cerbera manghas L.、杨叶肖槿Thespesia populnea (L.) Soland.和芦苇Phragmites australis (Cav.) Trin. ex Steud.等10个群落类型。采用目视解译的遥感分类方法,通过数字化提取研究区红树林群落信息。
2.2 样地调查
经过多次实地踏查,了解南沙湿地公园红树林群落分布的基本概况,验证Pleiades高分辨率多光谱的卫星影像解译信息。根据公园红树植物群落分布,选择一期10个、二期7个共17个代表性样地,样方面积乔木为10 m×10 m,灌木为5 m×5 m,草本为1 m×1 m,分别测量样方内植物的高度、胸径、基径,并记录其种类、株数等指标。实地调查样地位置如图1(2)所示,图中1~17为调研样点编号。野外调查工作于2013年8月—2014年3月进行。
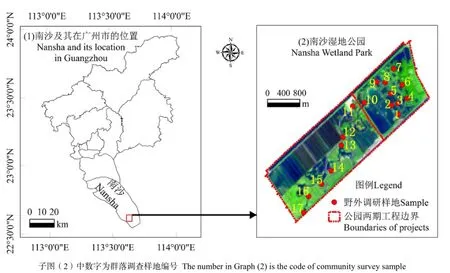
图1 研究区域位置图及群落调查样地Fig. 1 Sampling sites, study area and its location in Guangzhou of China
2.3 研究方法
2.3.1 物种多样性
物种多样性是度量群落稳定性的重要指标,受纬度、海拔和干燥度的影响,指数越高,群落越稳定。物种多样性测度指标选用物种多样性指数(Simpson指数SP与Shannon-Wiener指数SW)、和均匀度指数(E)等3个指标,计算公式如下(田广红等,2013):
Simpson指数(SP):

式中,N为全部样方中各植物种的个体数之和,ni为第i个种的个体数,Pi为第i个种的个体总数占比,S为样方中出现的植物种数。
2.3.2 优势度
根据野外样方调查所得的数据,计算种群的相对多度、相对频度、相对显著度和重要值等种群优势度指标,以反映种群在红树林群落的重要性。重要值=(相对多度+相对频度+相对显著度)/3(周葆华等,2008;马丹炜等,2009);草本以总优势度表示其在群落中的重要性,公式为:

式中,C′为相对盖度;H′为相对高度;重要值或总优势度的数值越大,表示该种群在群落中越重要,它是反映种群的地位和作用的一个综合指标(王伯荪等,1996)。
生态优势度是指优势度集中于一个或多个种群的程度,是综合各个种群重要性的优势度指标(王伯荪等,1996),指数值越大,说明优势种的地位越突出,种群分布越不均匀,其计算式为(李明顺等,1994):

式中,N为各植物种的重要值之和,ni为第i个种的重要值,S为植物种数。
2.3.3 空间分布格局
应用景观生态学中景观结构研究理论进行空间分布格局分析,采用景观格局分析方法(邬建国,2007)从集聚和斑块面积两个方面进行测度,选用相似邻接百分比(PLADJ)、丛生性指数(CLUMPY)、斑块聚合度AI和最大斑块指数(LPI)4个指标(何鹏等,2009;董翠芳等,2014;布仁仓等,2005)量化红树林群落的分布格局。其中,CLUMPY指数值为-1~1,PLADJ等指数值为0~100。
3 结果与分析
南沙湿地公园土地利用分布如图2,水域及水利设施用地和林地为其主要土地利用类型,分别占研究区面积的62.77%和29.34%;其他用地包括堤岸、裸地和空闲地,仅占总面积的4.29%;余下的草地、商服用地、住宅用地、交通运输用地、风景名胜及特殊用地等也只占总面积的3.60%。
3.1 南沙湿地公园红树林的物种多样性
根据野外调查数据,结合相关研究成果(廖宝文等,2014;林雪玲,2008),南沙红树林湿地公园有红树植物17种,占广东红树植物的63%,仅比粤东地区少1个种(李皓宇等,2016)。其中,真红树植物8科9属10种(彭友贵等,2005),占全国真红树种数的38.5%;半红树有6科7属7种(宋焱等,2016),占全国半红树种数的58.3%。真红树植物中有60%是乡土种,40%是引种栽培;半红树植物也有85.7%是乡土种,只有1种是引种栽培,具体见表1。物种多样性指数3个指标的测度结果分别为:Shannon-Wiener指数SW为3.438,Simpson指数D为0.890,均匀度指数E为0.880。
3.2 南沙湿地公园红树林的优势度
优势度按生态优势度和种群优势度进行测度,生态优势度测算结果为0.12,低于深圳福田红树林的0.56(李明顺等,1994),与鼎湖山常绿阔叶林的0.12(李明顺等,1994)相同。由此说明,南沙红树林优势种地位不如福田红树林的显著,群落内种群数量分布较均匀,类似鼎湖山常绿阔叶林。
红树林植物的种群优势度根据野外样方法调查数据计算所得,选用相对多度、相对显著度、相对频度和重要值共4个指标进行测算,结果见图3。重要值在10%以上的红树植物有无瓣海桑(S7)、桐花树(S1)和黄槿(S9),其值分别是24.41%、11.30%和10.26%。可见,南沙湿地公园红树林人工引进种以无瓣海桑为优势种;乡土种以桐花树和黄槿为优势种,二者重要值之和为21.56%。重要值在4%~8%之间的有秋茄(S2)、老鼠簕(S4)、水黄皮(S12)和杨叶肖槿(S10),其值分别为7.47%、6.17%、5.27%和4.70%。重要值在4%以下的有海芒果(S11)、木榄(S3)、拉关木(S8)和海桑(S6),四者之和仅占重要值的10.49%。此外,红树林中伴生种芦苇在该区比例较高,总优势度为76.06%,是草本层的优势种群。

图2 南沙湿地公园土地利用及红树林分布格局Fig. 2 Land use and distribution pattern of mangrove in Nansha Wetland Park
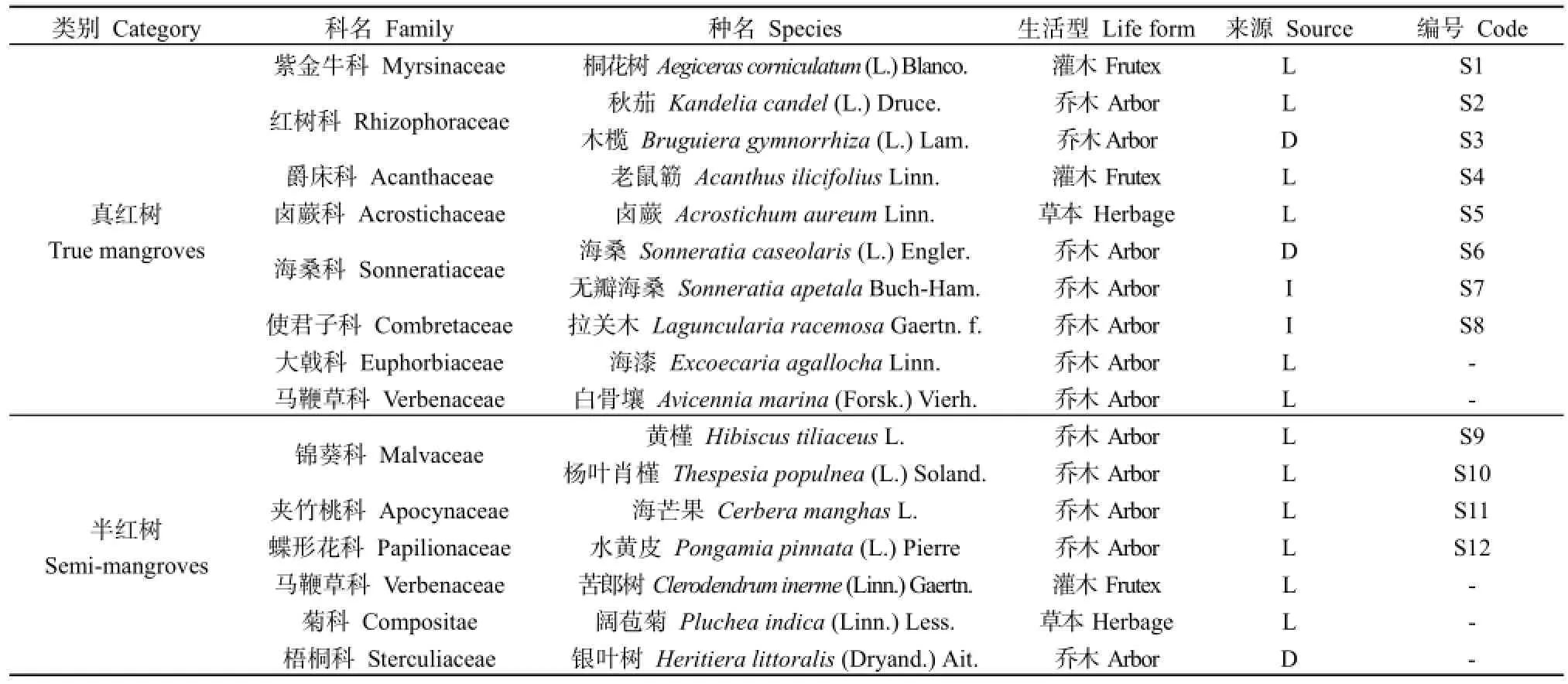
表1 南沙湿地公园红树植物种类组成Table 1 Plant flora or composition of the Mangrove in Nansha Wetland Park
3.3 南沙湿地公园红树林的群落特征
南沙湿地公园红树林群落主要包括无瓣海桑群落、桐花树群落、秋茄群落、拉关木群落、木榄群落、黄槿群落、水黄皮群落、海芒果群落、杨叶肖槿群落和芦苇群落共10个群落,从图2(3)、(4)红树林分布可见,无瓣海桑群落、黄槿群落和芦苇群落等为主要的红树林群落,总面积占比达86.72%(表2),其中无瓣海桑群落和黄槿群落面积占比分别为41.13%和36.44%,是研究区占主要优势的红树林群落;芦苇群落面积占比9.15%;桐花树群落、水黄皮群落、秋茄群落和拉关木群落面积占比接近,为2%~3%;海芒果群落面积占比不到2%;余下的2个群落面积占比都小于1%。
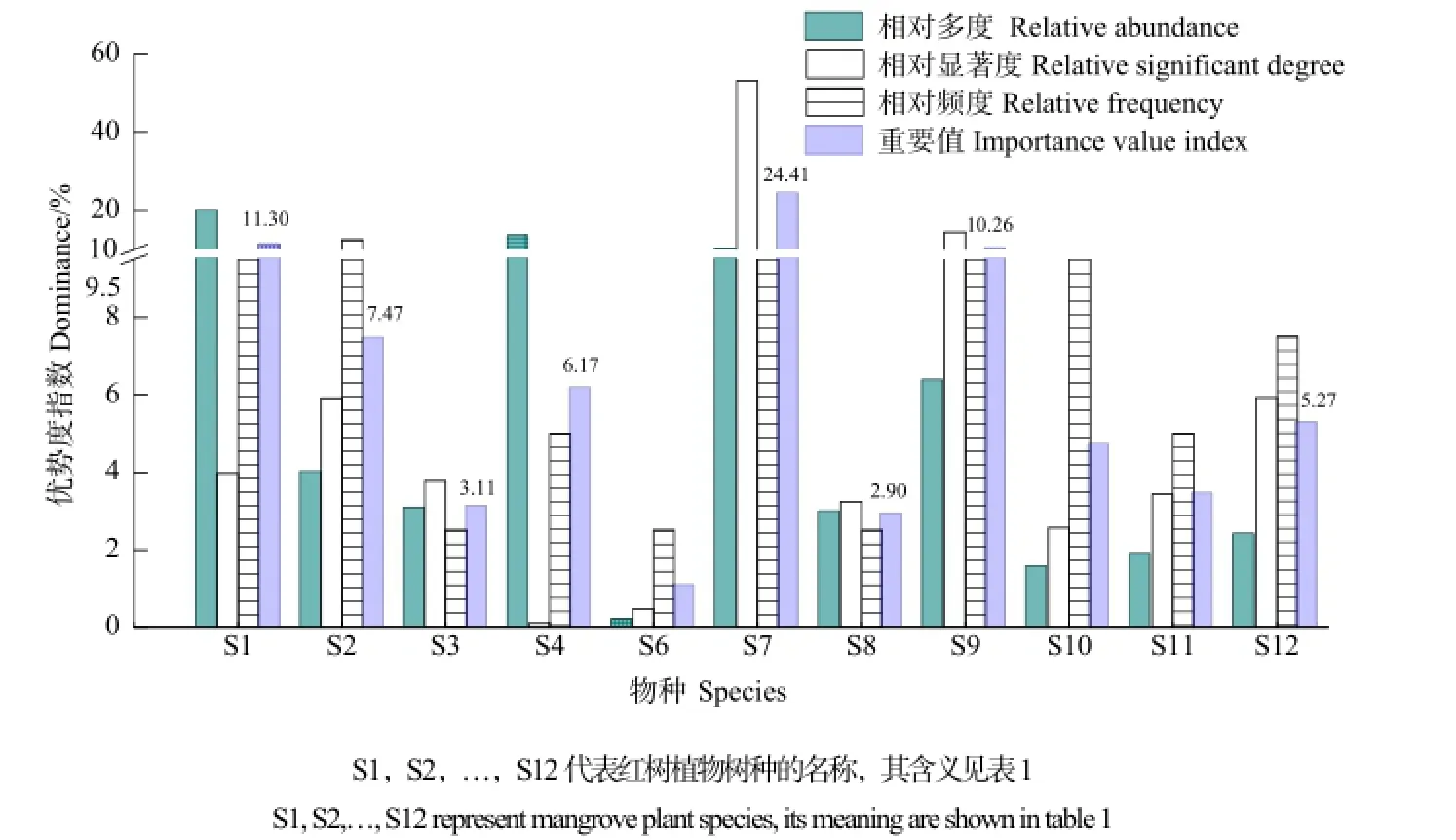
图3 南沙湿地公园主要红树林植物优势度测算Fig. 3 Determined dominance of the main mangrove plants in Nansha Wetland Park
南沙湿地公园第一期以无瓣海桑群落、芦苇群落和桐花树群落为主要群落类型,面积占比分别为67.46%、12.93%和5.87%;以真红树植物群落为主,面积占比达82%;其次是伴生种芦苇群落,面积占比是12.93%;半红树植物群落面积最小,仅有5.07%。第二期以黄槿群落、无瓣海桑群落和芦苇群落为主要群落类型,面积占比分别为70.48%、16.55%和5.63%;以半红树植物群落为主,面积占比达74.95%;其次是真红树植物群落,面积占比是19.43%;伴生种芦苇群落面积最小,仅有5.63%。
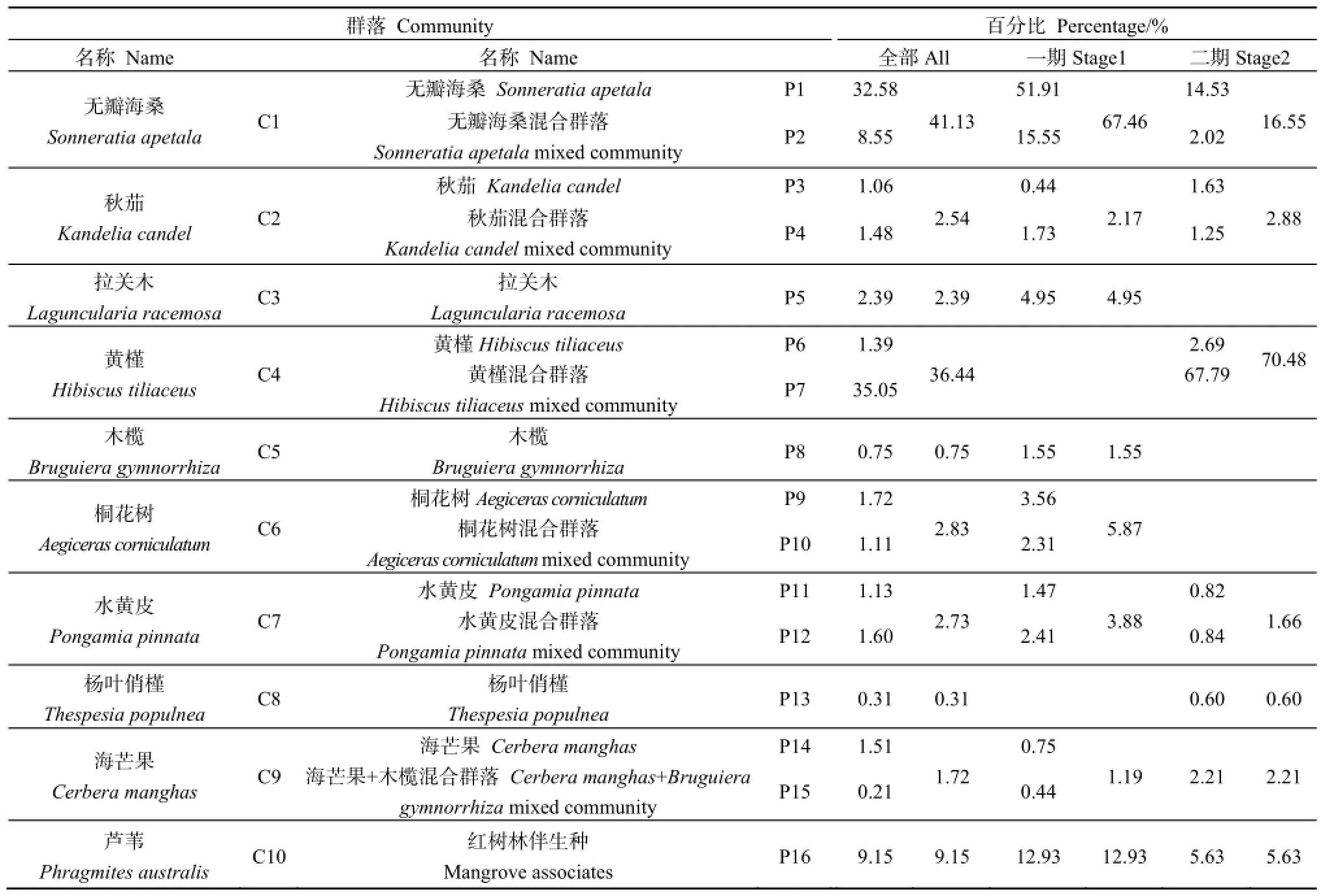
表2 南沙湿地公园红树林群落构成及比例Table 2 Composition and area percentage of mangrove community in Nansha Wetland Park
南沙湿地公园红树林主要群落特征:
(1)无瓣海桑群落(C1),群落外貌灰绿色,树形高大;乔木层常为单优种;郁闭度0.8~0.9;高度3.5~16 m;无瓣海桑株距有1 m和2 m两种,膝根多,在1 m×1 m样方中,有48根;有少量秋茄,偶见卤蕨Acrostichum aureum。呈片状分布在一期东北部,二期北部。
(2)秋茄群落(C2),群落外貌翠绿色,整齐;乔灌两层;郁闭度0.7~0.8;高度1.8~4.5 m;伴生少量水黄皮、桐花树,老鼠簕Acanthus ilicifolius散生其中,偶见海芒果、卤蕨;呈带状、块状分布在水塘堤岸上。
(3)拉关木群落(C3),群落外貌林相整齐;单优种,有层间藤本;郁闭度0.6~0.7;高度3~4.5 m;株距2 m;林下有卤蕨、海芋Alocasia macrorrhizos、华南毛蕨Cyclosorus parasiticus等草本植物;层间多微甘菊Mikania micrantha,茎粗1.9 cm,缠绕树上;幼苗在1 m×1 m样方中,有250棵,均高20 cm;呈块状分布在一期码头附近,主航道两侧。
(4)黄槿群落(C4),群落外貌深绿色;乔木层种类多;郁闭度0.6~0.7;高度2.5~3 m;常与小叶榕Ficus microcarpa、柳树Salix babylonica L.、楹树Albizia chinensis等非红树植物混生,林下有老鼠簕,也有些是单优种纯林;呈块状分布在西南部。
(5)木榄群落(C5),群落外貌为林相整齐;纯林;郁闭度0.6~0.7;高度2~2.3 m;偶见非红树植物混生其中;呈条带状分布在一期中部水塘堤岸。
(6)桐花树群落(C6),群落外貌黄绿色,平整灌丛;结构简单;郁闭度0.5~0.7;高度1.5 m;伴生少量银叶树Heritiera littoralis、芦苇,偶见秋茄,有非红树植物海芋、美人蕉Canna indica混生其中,群落边缘散生卤蕨;呈带状、环状分布在一期木榄群落西侧的南北两端。
(7)水黄皮群落(C7),群落外貌树冠平整;乔灌两层;郁闭度0.7;高度2.2~3.5 m;伴生少量桐花树,有非红树植物大叶榕Ficus virens混生;呈带状、块状分布在公园中部。
(8)杨叶俏槿群落(C8),群落外貌林相整齐;结构完整,有层间藤本;郁闭度0.6;高度3~6 m;植株茂密分枝多,多为7~8分枝,林下有老鼠簕、卤蕨,层间非红树植物微甘菊多,茎粗2 cm;呈块状散生在二期东北部。
(9)海芒果群落(C9),群落外貌树冠平整;结构简单;郁闭度0.7;高度3 m;林下有少量木榄,部分是纯林;呈块状分布在公园中部。
(10)芦苇群落(C10),群落外貌灌状草丛;单一草本层;郁闭度0.95;高度1.6~2.3 m;单优种草本群落;呈片状集中分布在一期无瓣海桑群落的西侧,二期的东北部。
南沙湿地公园人工次生湿地红树林群落外貌呈波状起伏,结构简单,大多为乔木状林,草本植物以伴生种芦苇为主,其群落面积较大。真红树植物群落主要集中分布在公园一期,以无瓣海桑群落面积为最大;半红树植物群落主要分布在公园二期,以黄槿群落面积为最大。由公园东北向西南方向,一期树高由高→低→高,红树林长势较好,植株高大,郁闭度也高于二期样地。
3.4 南沙湿地公园红树林群落空间分布格局
运用景观生态学方法,探讨南沙湿地公园红树林群落的空间分布。红树林群落的景观格局指数曲线见图4,3个反映景观集聚度的指数(CLUMPY、PLAD、AI)都显示[如图4(a)、4(b)],红树林各群落在空间上都呈聚集态分布,各指数值不同,但曲线波动基本一致。具体表现为丛生性指数(CLUMPY)值都大于0,指数值最高的是无瓣海桑群落(C1),为0.7181,最低的是木榄群落(C5),为0.0274;相似邻接百分比(PLADJ)和斑块聚合度(AI)值相近。红树林各群落的3种指标的指数值表明,聚集度最高的是无瓣海桑群落(C1),黄槿群落(C4)次之,这两个群落的指数值接近;最低的是木榄群落(C5),其指数值与其他群落差距较大。
最大斑块指数(LPI)[图4(c)]显示,无瓣海桑群落(C1)的斑块面积最大,其最大斑块面积占研究区总面积的6.39%;黄槿群落(C4)的斑块面积次之,其最大斑块面积占比是2.88%;余下的红树林群落斑块面积占比都比较小,均在1%以下。
4 讨论
4.1 物种多样性的对比与变化
南沙红树林与其他与之纬度相近的红树林和南亚热带常绿阔叶林相比,物种多样性指数SW高于惠州澳头镇红树林(1.904)(陈桂珠等,1994)、湛江红树林(2.232)(缪绅裕等,1998),但明显低于黑石顶(4.57)(李明顺等,1994)、南昆山(5.37)(李明顺等,1994)的南亚热带常绿阔叶林。南沙湿地公园红树林群落的物种多样性较其他与之纬度相近的红树林群落高,这得益于1998年南沙湿地公园引种无瓣海桑、海桑、拉关木、木榄等红树植物,丰富了该区的红树林种类;较常绿阔叶林低,是受红树林特殊生长环境限制,相比常绿阔叶林,其种类较少,群落结构简单。就红树林群落而言,均匀度指数E表现为南沙高于惠州澳头镇(0.737)(陈桂珠等,1994),该区中、低盐度是红树林最适合的生境(李皓宇等,2016),可见,南沙湿地公园红树林能较好地适应本区咸淡水交界的湿地生态环境。在经历了引种扩种的人为干预后,南沙湿地红树林在与自然环境的相互作用下多样性指数仍高于其他与之纬度相近的澳头和湛江的红树林,说明该区红树林群落较这两地更趋于稳定。
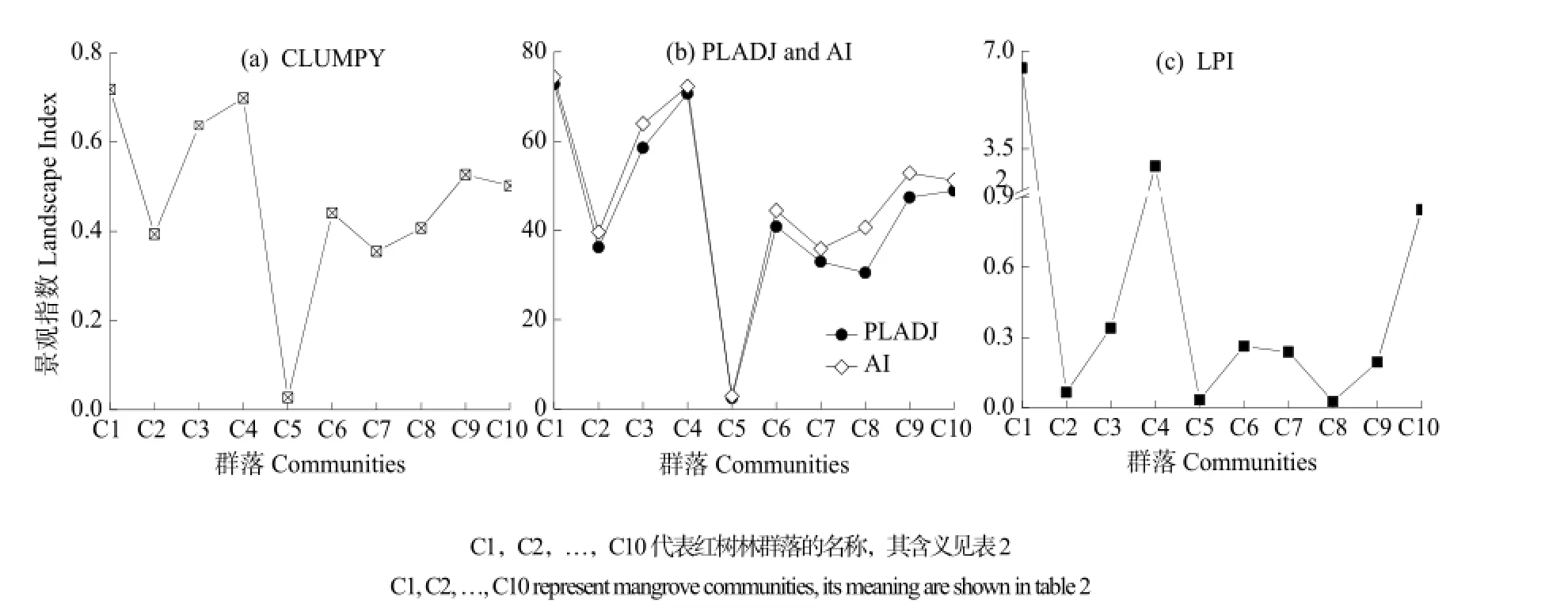
图4 2013年南沙湿地公园红树林群落的景观格局指数曲线Fig. 4 Curves of landscape indices for mangrove community in Nansha Wetland Park in 2013
种群重要值的测算结果表明,无瓣海桑(S7)、桐花树(S1)、黄槿(S9)和芦苇(S13)为优势种。引进种无瓣海桑(S7)植株高大,长速快,最矮的植株3.5 m,最高的16 m,均高8.54 m;冠幅大;膝根多,均高22 cm;抗寒性强,不受寒害影响。拉关木(S8)在珠江口红树林区均有引种,该区长势比珠海淇澳岛的好,高达4.5 m,林下幼苗在1 m×1 m的样方中多达250株,均高20 cm,有自然更新现象,具有抗寒性,受寒害影响程度弱。海桑(S6)是高大乔木,从海南岛引种,属嗜热性种类,不耐寒冷,受2008年南方寒害影响,大部分死亡,剩余数量较少,未能成为优势种群。乡土种桐花树(S1)和秋茄(S2)是真红树植物,应该是本区红树林的优势种,但长速慢,被引进种无瓣海桑(S7)抢据优势,桐花树(S1)重要值偏低,秋茄(S2)未能成为优势种。伴生种芦苇(S13)的总优势度占草本层的76.06%,成为草本层的绝对优势种。该区通过引种丰富了物种多样性,引进种除海桑外,其他红树植物长势良好。
4.2 红树林群落变动与分布
南沙湿地公园是省级红树林湿地公园,先后分两期对外开放,第一期2008年,第二期是2013年(宋焱等,2016)。红树林群落总共有十大类,第一期有八类,第二期有七类。第一期以真红树植物为主,无瓣海桑群落(C1)具有最大斑块面积;第二期以半红树植物为主,黄槿群落(C4)具有最大斑块面积,此外,还有引进种无瓣海桑群落(C1),面积占比仅次于黄槿群落(C4)。此格局的形成是由于公园第一期引种外来种,物种数量增加,引进种生长迅速,成林分布,秋茄在无瓣海桑群落中零星分布,引进种无瓣海桑抑制了乡土种秋茄的增长。公园第二期主要是以乡土种半红树植物黄槿为主,真红树植物所占比例较少。黄槿多与非红树植物混生,长在第二期公园西南部高潮位上。
与2008年相比(见表3),群落结构变动大,群落演替也由逆向演替(林雪玲,2008)向顺向演替发展,群落稳定性较2008年的好。群落变动的原因,一是群落类型增加,二是部分群落类型面积减少。由于海桑受寒害影响严重,大部分冻死,导致无瓣海桑+海桑群落消失,取而代之的是无瓣海桑纯林或以无瓣海桑为优势种的混合群落(如无瓣海桑+秋茄群落)。黄槿+杨叶肖槿群落已不是主要的群落类型,黄槿和非红树植物混生的群落面积更大。公园第二期,红树植物群落除秋茄群落(C2)和无瓣海桑群落(C1)为真红树植物群落外,其余4个群落都为半红树植物群落。无瓣海桑与生态位较高的秋茄以及高生态位的海芒果、杨叶肖槿混生,可见无瓣海桑适应性强。

表3 2008年和2013年南沙湿地公园主要红树植物群落比较Table 3 Comparison of main mangrove community between in 2008 and 2013 in Nansha Wetland Park
4.3 红树林群落聚集分布格局
基于景观生态学的红树林群落空间分布格局显示,南沙湿地红树林植物群落呈聚集分布。聚集度最高的是无瓣海桑群落(C1),其次是黄槿群落(C4),最低的是木榄群落(C5)。南沙湿地公园红树林聚集分布,一是与其人工种植方式直接相关;二是由于红树植物具有独特的胎生现象,种子在母树发育,胚轴成熟后,直接掉落在母树周围,发育成幼苗,形成聚集分布格局。该研究区群落处于演替早期阶段,聚集度较高,群落正处于正向演替进程中,聚集度将会逐渐降低。
群落构成与分布结果表明,南沙湿地红树林形成了以无瓣海桑群落(C1)、黄槿群落(C4)和芦苇群落(C10)等3种群落为主的红树林群落分布格局。面积占比最大的无瓣海桑是引进种,而余下的两类乡土种群落,均不是真红树植物群落;面积位居第二的黄槿群落(C4)其面积占比是36.44%,其中35.05%是混合群落,占黄槿群落总面积的96.18%,主要是与非红树植物混生。由此说明,该研究区红树林群落具有不稳定性。
优势种与主要群落类型存在差异,这源于优势种的测算所采用的数据为样方调查所得,样方点设置的数量直接影响优势种的测度结果。公园一期主要群落类型与优势种一致,而公园二期有差异。一般而言,主要群落是由优势种形成的群落,这与本研究结论一致。
5 结论
南沙湿地公园红树林物种多样性指数较高,形成了以无瓣海桑(S7)、桐花树(S1)、黄槿(S9)和芦苇(S13)为优势种,以无瓣海桑群落(C1)、黄槿群落(C4)和芦苇群落(C10)等3种群落为主的红树林群落分布格局。无瓣海桑(S7)重要值占比24.41%,以纯林为主,纯林面积占比32.58%,与秋茄、海芒果、杨叶肖槿等分别形成的混合群落面积占比仅有8.55%;无瓣海桑混合群落面积仅占无瓣海桑群落总面积的20.79%。而乡土树种秋茄和桐花树本应是该红树林区的优势种,但因生长较慢、树型小而被无瓣海桑替代,可见,无瓣海桑对乡土树种的生长存在抑制作用。建议提高先锋树种白骨壤Avicennia marina (Forsk.) Vierh.和桐花树(S1)种群数量,扩大乡土种秋茄(S2)的面积,提升这3个树种的优势度,以降低引进种无瓣海桑(S7)和半红树植物黄槿(S9)的优势度,形成以乡土树种为主的真红树植物群落,增强该区红树林群落的稳定性。扩种乡土树种将是该区今后红树林恢复的首要任务。
致谢:广州大学地理科学学院2014级硕士研究生陈康林协助制图,2011级学生冯展涛参与野外调查工作,在此一并感谢!
GIRI C, PENGRA B, ZHU Z L, et al. 2007. Monitoring mangrove forest dynamics of the Sundarbans in Bangladesh and India using multitemporal satellite data from 1973—2000 [J]. Estuarine Coastal & Shelf Science, 73(1-2): 91-100.
MAS J F. 1999. Monitoring land-cover changes: a comparison of change detection techniques [J]. International Journal of Remote Sensing, 20(1): 139-152.
SETO K C, FRAGKIAS M. 2007. Mangrove conversion and aquaculture development in Vietnam: A remote sensing-based approach for evaluating the Ramsar Convention on Wetlands [J]. Global Environmental Change, 17(3-4): 486-500.
SIMARD M, DE-GRANDI G E, SAATCHI S, et al. 2002. Mapping tropical coastal vegetation using JERS-1 and ERS-1 radar data with a decision tree classifier [J]. International Journal of Remote Sensing, 23(7): 1461-1474.
WANG B S, LIANG S C, ZHANG W Y, et al. 2003. Mangrove flora of the world [J]. Actor Botanica Sinica, 45(6): 644-653.
布仁仓, 胡远满, 常禹, 等. 2005. 景观指数之间的相关分析[J]. 生态学报, 25(10): 2765-2775.
常弘, 廖宝文. 2009. 广州市南沙湿地公园的鸟类多样性与禽流感监控[J]. 湿地科学与管理, 5(1): 64-65.
常永智, 丁四保. 2013. 消失的红树林: 生态之殇[J]. 生态经济, (11): 18-23.
陈桂珠, 缪绅裕. 1994. 广东澳头红树植物群落的物种多样性与种群分布格局研究[J]. 生态学杂志, 13(2): 34-35.
但新球, 廖宝文, 吴照柏, 等. 2016. 中国红树林湿地资源、保护现状和主要威胁[J]. 生态环境学报, 25(7): 1237-1243.
董翠芳, 梁国付, 丁圣彦, 等. 2014. 不同干扰背景下景观指数与物种多样性的多尺度效应——以巩义市为例[J]. 生态学报, 34(12): 3444-3451.
何鹏, 张会儒. 2009. 常用景观指数的因子分析和筛选方法研究[J]. 林业科学研究, 22(4): 470-474.
黎夏, 刘凯, 王树功. 2006. 珠江口红树林湿地演变的遥感分析[J]. 地理学报, 61(1): 26-34.
李皓宇, 彭逸生, 刘嘉健, 等. 2016. 粤东沿海红树林物种组成与群落特征[J]. 生态学报, 36(1): 252- 260.
李玫, 陈桂珠, 彭友贵, 等. 2009. 广州南沙湿地生物多样性现状及其保护[J]. 防护林科技, (3): 46-48.
李明顺, 蓝崇钰, 陈桂珠, 等. 1994. 深圳福田红树林的群落学研究Ⅱ.多样性与种群格局[J]. 生态科学, (1): 82-86.
李文华. 2011. 我国生态学研究及其对社会发展的贡献[J]. 生态学报, 31(19): 5421-5428.
廖宝文, 张乔民. 2014. 中国红树林的分布、面积和树种组成[J]. 湿地科学, 12(4): 435-440.
林雪玲. 2008. 广州市湿地植物群落特征与分布演替分析[J]. 防护林科技, (4): 115-117, 142.
马丹炜, 马宏, 李群, 等. 2009. 植物地理实验与实习教程[M]. 北京: 科学出版社: 72.
缪绅裕, 陈桂珠, 陈正桃, 等. 1998. 广东湛江保护区红树林的生物量和种群分布格局[J]. 广西植物, 18(1): 19-23.
缪绅裕, 李德宁, 邓鸿英, 等. 2011. 广州南沙3种红树植物生理生态特性的比较[J]. 广州大学学报(自然科学版), 10(1): 31-36.
彭友贵, 陈桂珠, 夏北成, 等. 2004. 广州南沙地区湿地生态系统的服务功能与保护[J]. 湿地科学, 2(2): 81-87.
彭友贵, 陈桂珠, 夏北成, 等. 2005. 广州南沙地区湿地生态系统的特征分析[J]. 海洋环境科学, 24(3): 51-55.
邱彭华, 徐颂军, 符英, 等. 2011. 广州南沙区万顷沙人工次生湿地植物群落初步研究[J]. 生态科学, 30(1): 43-50.
宋焱, 徐颂军, 刘贤赵, 等. 2016. 南沙红树林湿地公园水环境质量时空差异分析——基于改进后倍斜率聚类分析的视角[J]. 地理科学, 36(2): 303-311.
田广红, 丁明艳, 杨雄邦. 2013. 珠海市淇澳岛肉实树群落及其物种多样性特征[J]. 植物科学学报, 31(5): 461-466.
王伯荪, 余世孝, 彭少麟, 等. 1996. 植物群落学实验手册[M]. 广州:广东高等教育出版社: 85-102.
邬建国. 2007. 景观生态学——格局、过程、尺度与等级(第二版)[M]. 北京: 高等教育出版社, 66-72.
徐越, 郑晓彤, 金佩欣, 等. 2013. 国家新区南沙湿地旅游资源生态效益探讨[J]. 广东农业科学, (17): 189-192.
张伟, 张义丰, 张宏业, 等. 2010. 生态城市建设背景下湛江红树林的保护与利用[J]. 地理研究, 29(4): 607-616.
张晓君, 管伟, 朱宁华, 等. 2014. 广州南沙人工林海桑属群落及土壤的动态变化[J]. 中南林业科技大学学报, 34(9): 103-106.
周葆华,余世金. 2008. 天柱山黄山松群落特征及其环境功能评价[J]. 地理研究, 27(2): 257-265.
朱可峰, 廖宝文, 章家恩. 2011. 广州南沙人工红树林凋落物组成与季节变化的研究[J]. 华南农业大学学报, 32(4): 119-121.
Species Diversity and Spatial Distribution Pattern of Mangrove in Nansha Wetland Park, Guangzhou, Guangdong Province, China
QIU Ni1,2, XU Songjun1*, QIU Penghua3, SONG Yan1, NIU Anyi1, XU Guanchang1
1. School of Geographical Sciences, South China Normal University, Guangzhou 510631, China; 2. School of Geographical Sciences, Guangzhou University, Guangzhou 510006, China; 3. College of Geography and Tourism, Hainan Normal University, Haikou 571158, China
The mangrove is a unique vegetation type along the coast, and its species diversity and spatial distribution are studied. This is of great significance to the restoration and protection of the mangrove forests. In this paper, the mangrove forest in the Nansha Wetland Park is a typical case and the species diversity, community characteristics and spatial distribution of the mangrove forests are studied by means of remote sensing image interpretation and quadratic surveys. The results showed that, (1) The mangrove species diversity index of the Nansha wetland park was higher than that of the mangrove community at a similar latitude. (2) The dominant species were Sonneratia apetala, Aegiceras corniculatum, Hibiscus tiliaceus and Phragmites australis. (3) The main community types were Sonneratia apetala community, Hibiscus tiliaceus community and Phragmites australis community, with a percentage of the total area of 86.72%. The area of the Sonneratia apetala community was 41.13% and the area of Hibiscus tiliaceus community was 36.44%. The two communities lead to become the absolute dominant of the mangrove community. (4) The spatial distribution of mangrove community was aggregated. The highest aggregation of the mangrove community was Sonneratia apetala community, followed by Hibiscus tiliaceus community, the lowest was Bruguiera gymnorrhiza community. (5)Through the introduction of the species Sonneratia apetala we saw it was good at growing and limited the local mangrove of growing. It is suggested to expand Avicennia marina, Aegiceras corniculatum and Kandelia candel, and therefore increase the plaque area to enhance the stability of the local mangrove community and effectively protect the mangrove.
mangrove; landscape pattern; spatial distribution; species diversity; Nansha Wetland Park
10.16258/j.cnki.1674-5906.2017.01.005
Q948; X176
A
1674-5906(2017)01-0027-09
邱霓, 徐颂军, 邱彭华, 宋焱, 牛安逸, 许观嫦. 2017. 南沙湿地公园红树林物种多样性与空间分布格局[J]. 生态环境学报, 26(1): 27-35.
QIU Ni, XU Songjun, QIU Penghua, SONG Yan, NIU Anyi, XU Guanchang. 2017. Species diversity and spatial distribution pattern of mangrove in Nansha Wetland Park, Guangzhou, Guangdong Province, China [J]. Ecology and Environmental Sciences, 26(1): 27-35.
国家自然科学基金项目(41271060;41671175);国家重点研发计划项目(2016YFC0502803)
邱霓(1974年生),女,高级实验师,博士研究生,主要研究方向为植物地理与景观生态。E-mail: qiuni2@sina.com *通信作者:徐颂军,教授,博士。E-mail: xusj@scnu.edu.cn
2016-12-26
