关帝山油松天然林林下植物组成及环境解释
2017-04-01王慧南宏伟刘宁
王慧,南宏伟,刘宁
山西农业大学林学院,山西 太谷 030801
关帝山油松天然林林下植物组成及环境解释
王慧,南宏伟,刘宁*
山西农业大学林学院,山西 太谷 030801
作为森林生态系统的重要组成部分,林下植物在调控森林结构和功能方面发挥着重要的作用。通过对华北山地干旱区油松(Pinus tabulaeformis Garr.)天然纯林林下植物群落的调查,探究林下植物物种组成与林分、环境因子之间的关系及其关键控制因子,旨在为华北山地干旱区林下生物多样性保护和维持提供依据。结果表明,(1)在调查的样地中,油松林下共出现植物物种72种,隶属33科65属,其中蔷薇科、菊科、豆科物种种类丰富,占全部种数38.8%;林下灌木的物种多为喜光或兼性树种,与油松林特殊的冠形有关,林下草本以多年生草本占优势。(2)CCA排序较好地揭示了该区油松天然林林下植物的分布格局与环境因子的关系,生物因子与非生物因子共同对林下物种的分布格局产生作用,对林下灌木层和草本层的解释率分别为59.92%和66.16%;其中,地形因子的作用大于林分因子和土壤因子,在油松天然林林下植物分布格局中起着举足轻重的作用。林下灌木层物种多样性在下坡最高、上坡最低,并表现为西北坡高于其他坡向;林下草本层物种多样性随坡位的变化与灌木层正好相反,在上坡最高、下坡最低,在所选样方范围内,草本层物种多样性随海拔的升高而升高。
华北山地;油松天然纯林;林下植物;分布格局;环境因子
林下植物是森林生态系统中的重要组成部分,是森林生态系统中生物多样性的主要来源,在维持森林群落内的营养平衡、生物多样性、林下更新和生态系统长期稳定性方面有着不可忽视的作用(Nilsson et al.,2005;Chastain et al.,2006;Gamfeldt et al.,2013;Wu et al.,2014)。近年来,除林下植物结构特征、分布格局、生态功能等被广泛研究外,环境因子与林下植被分布格局的影响也已被广泛讨论,逐步得到重视(丛静等,2013;刘宏文等,2014;尤业明等,2016)。
山地植物群落物种多样性的变化规律一直是生态学家感兴趣的问题(丛静等,2013)。干旱区因难以保持水分需求平衡而成为全球生态脆弱区,也是生物多样性保护和维持的重要研究区域。华北山地半干旱区森林生态系统是研究干旱区山地植物群落多样性的理想场所。油松(Pinus tabulaeformis Garr.)是华北地区的特有树种,也是华北中、低海拔山地的主要优势树种,在该区海拔1200~1800 m广泛分布且长势良好;辽东栎(Quercus liaotungensis)和山杨(Populus davidiana)为其主要伴生的阔叶树种。一般认为,油松林所具有的独特的林冠结构和群落条件,导致其林下环境极差难以形成连续的林下植被层,以致油松纯林结构单一而形成“绿色沙漠”(郭华等,2005),因此华北山地垂直层次分明的油松林则具有一定的群落多样性研究价值。
本研究以华北山地油松天然林群落为研究对象,利用典型对应分析(CCA排序)探究林下植物群落组成与环境因子之间的关系,确定华北半干旱区影响油松天然林林下物种分布的主要因子,以期为其林下生物多样性保护提供有力的理论依据。
1 材料与方法
1.1 研究地概况
研究区位于山西省中部吕梁山脉,地理坐标为东经110°0′~111°37′,北纬34°34′~37°43′,海拔为770~2850 m。研究区属温带大陆性季风气候,冬季寒冷干燥,夏季炎热多雨,降水主要集中在6—8月,约占全年的60%以上,降水量在400~500 mm之间,蒸发量在1600 m以上,无霜期平均145 d。山地成土母岩主要有石灰岩和砾岩等,土壤为风积黄土母质,表层土壤类型主要为褐土,土层10~60 cm厚薄不等,腐殖质层0~10 cm,一般取决于植被覆盖状况。
1.2 样地调查
在研究区采用完全随机设计法设置样地,共15个,样地面积至少1 hm2,所选样地均为完全郁闭纯林林分(郁闭度≥0.75,优势树种断面积比例≥70%)。于2015年夏季进行野外调查,在所选样地内设置调查样方,样方面积为20 m×20 m,对样方内林冠层活立木(DBH≥10 cm)进行每木检尺;在每个样方内采用生长锥法估测其优势树种年龄,以样地内优势树种平均年龄作为林分年龄的保守估计;选用标准木建立树高-胸径曲线模型计算林分平均高。
高度低于4 m的木本植物视为灌木层(Griffiths et al.,2007;Bartels et al.,2013),包括下木、未进入林冠层的幼苗和幼树;高度低于1.3 m的维管植物被视为草本层(Bartels et al.,2013)。将每个样方划分为25个4 m×4 m的网格和400个1 m×1 m的网格,随机选取能代表此样方灌木和草本群落结构的调查小样方各5个,各层调查小样方共计75个。在每一小样方内,记录灌木(幼树)、草本植物种类,统计其多度,估算其盖度(Légaré et al.,2001)。样地概况见表1。
1.3 土壤样品采集与测定
在每个样方内,按照“S”形取样法设5个土壤采样点,分别对腐殖质层、0~20 cm表层土壤进行取样,混合后采用四分法均匀取样后置于冰盒中带回实验室,冷冻待测(Brosofske et al.,2001)。采用烘干法测定土壤含水量(Riegel et al.,1992),复合电极测定土壤pH值(Riegel et al.,1992),灼烧法测定腐殖质层和表层土壤总C(Qian et al.,2003),凯氏定氮法测定总N,氟化铵-盐酸浸提法测定有效P(Chen et al.,2004)。记录样方的地形因子及土壤类型。样方环境特征见表2。
1.4 数据分析及处理
本研究采用物种重要值(IV)反映林下灌木层和草本层物种在其相对应群落中的地位:
重要值IV=(相对密度+相对频度+相对基盖度)/3
林下灌木层和草本层物种多样性采用Shannon-Weiner指数进行评价:

式中,H′为物种多样性指数,Pi为种i的相对重要值IV(Alatalo,1981;孟祥楠等,2012)。
野外调查样方、林下物种多样性及环境因子数据采用Excel建库,对数据进行初步整理。基于野外测定和分析,共得到12个生物及环境因子,包括林分因子2个:林龄(SA)、林分密度(Den);主要的土壤养分因子6个:表层土壤总碳含量(TC)、土壤总氮含量(TN)、土壤有效磷含量(SP)、腐殖质层总碳含量(HC)、腐殖质层总氮含量(HN)、腐殖质层有效磷含量(HP);地形因子4个:海拔(E)、坡位(P)、坡向(A)、坡度(S)。为便于建立环境数据矩阵,将坡位和坡向数据以等级制表示,将坡位划分为:0.2代表下坡,0.6代表中坡,1代表上坡;根据实地样地调查的结果,油松林天然林多分布在东北坡、北坡、西北坡、西坡4个坡向,因此将坡向划分为:0.2代表东北坡(以正北为0°,顺时针旋转22.5°~67.5°),0.4代表北坡(0°~22.5°,337.5°~360°),0.6代表西北坡(292.5°~337.5°),0.8代表西坡(247.5°~292.5°)。
首先,对所选的6个土壤养分因子进行Pearson相关分析,根据分析结果采用PCA进行降维处理以确定土壤环境因子。然后分别以15个样方林下灌木层、草本层物种重要值矩阵(A28×15,B44×15)和样地生物及环境矩阵(Zx×15)为基础进行典范对应分析(CCA)和偏分析(Partial methods),分析生物及环境因子对物种分布的影响,并采用蒙特卡罗置换检验(Monte Carlo permutation test)分析环境因子与物种分布之间相关关系的显著性,在此基础上以物种多样性指数为响应变量、具有显著影响作用的环境因子为解释变量,分析环境因子对林下物种多样性的影响。数据分析在R3.3.1下完成。

表1 油松天然林样地概况Table 1 Summary of site characteristics and sites sampled (Range)

表2 油松天然林样地环境特征概况Table 2 Environmental characteristics of the study stands (Range)
2 结果与分析
2.1 油松林林下植物组成
在所调查的15个样地中,共记录林下植物72种,分属33科65属(见表3),其中蔷薇科、菊科、豆科合计23属28种,占全部种数38.8%,表明这3大科植物在油松天然林林下植物多样性中占有重要地位。
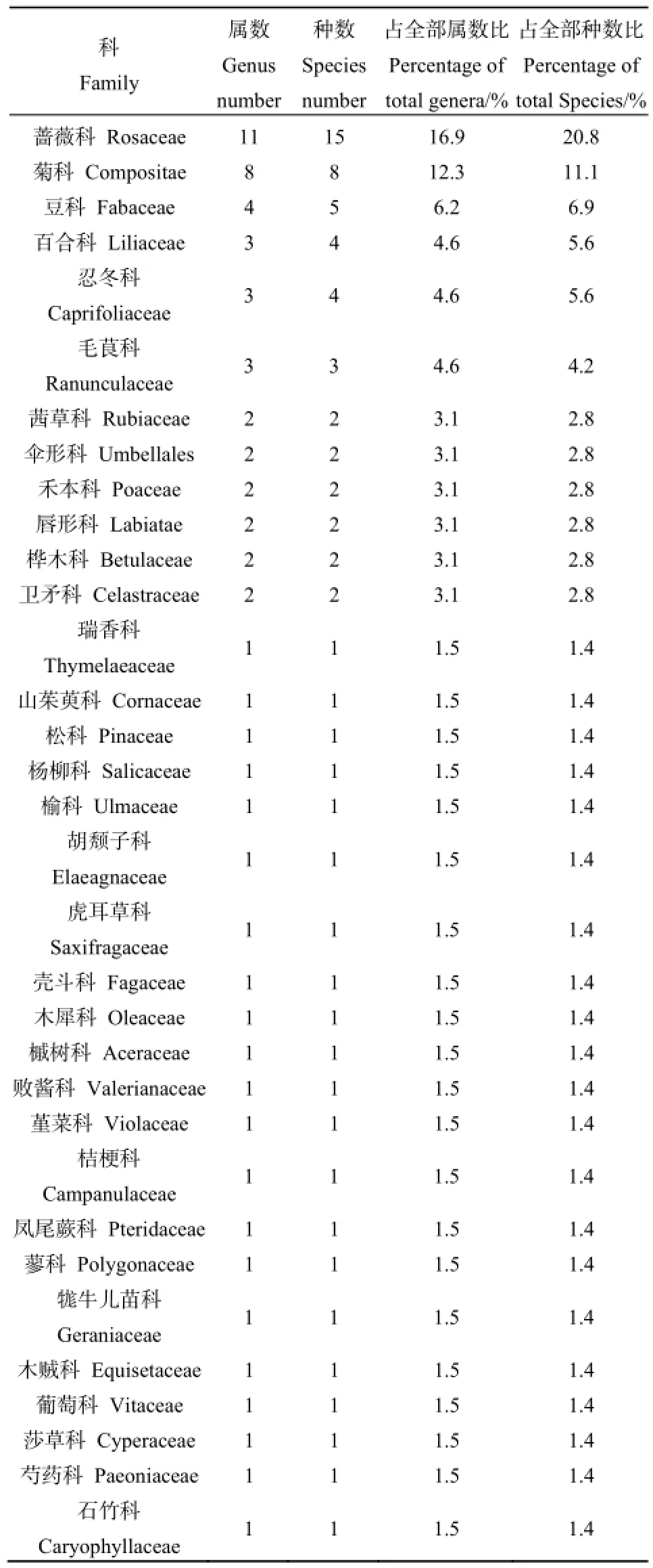
表3 油松林林下植物科属Table 3 Family and genus distribution of species
调查样方中林下灌木植物28种,分属于15科24属,其中36%的树种生长于林缘或疏林中,18%的树种自然分布于林下或密林中,而46%的树种为兼性树种。灌木层常见的植物种类有:土庄绣线菊(Spiraea pubescens Turcz.)、二色胡枝子(Lespedeza bicolor Turcz.)、虎榛子(Ostryopsis davidiana Decaisne)、水栒子(Cotoneaster multiflorus Bge.)、美蔷薇(Rosa bella Rehd. et Wils)、山刺玫(Rosa davurica Pall.)等。林下草本植物44种,分属于22科41属,其中1、2年生的草本植物有4种,分别为早熟禾(Poa annua L.)、泥胡菜(Hemistepta lyrata Bunge)、苦苣菜(Sonchus oleraceus L.)和猪殃殃(Galium aparine Linn. var. tenerum (Gren. et Godr.) Rchb.);多年生的草本植物有40种,草本层常见的植物种类有:披针叶苔草(Carex lanceolata Boott)、早熟禾(Poa annua L.)、唐松草(Thalictrum aquilegifolium Linn. var. sibiricum Regel)、小红菊(Dendranthema chanetii)、茜草(Rubia cordifolia L.)、柴胡(Bupleurum chinense)等。
2.2 土壤因子相关性及PCA分析
对所选的6个土壤因子进行Pearson相关性分析,结果见表4。
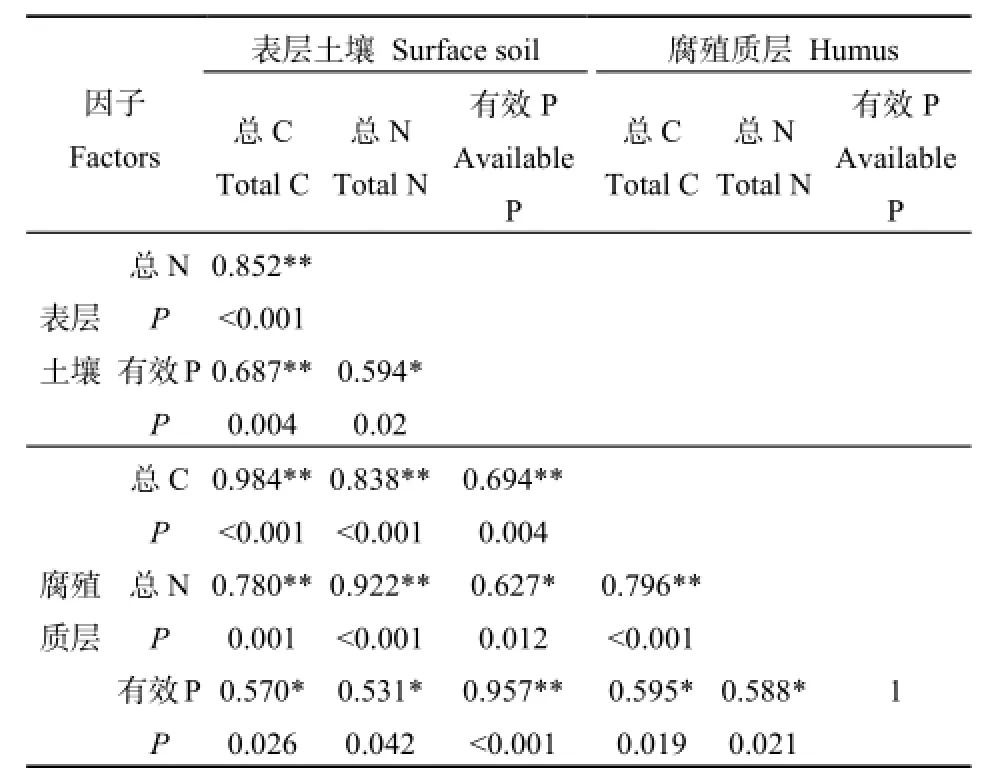
表4 土壤环境因子Pearson相关分析Table 4 Pearson correlation analysis of soil environmental factors
由Pearson相关分析可知,6个养分因子之间均呈不同水平的显著正相关关系,进一步对这6个养分因子进行主成分分析,结果见表5和图1。PCA分析结果显示,前2个主分量的累积贡献率已经达到了92%以上,其中,第一主分量的累积贡献率为78.087%,以表层土壤总C、总N、腐殖质层总C、总N含量为主,主要反映土壤C、N含量状况,重新命名为土壤碳氮(CN);第二主成分中主要因素为表层土壤有效P含量、腐殖质层有效P含量,主要反映土壤P含量状况,重新命名为土壤有效磷(AP)。因此,根据PCA结果,以2个主成分值土壤碳氮(CN)、土壤有效磷(AP)代表土壤环境因子进行后续分析。
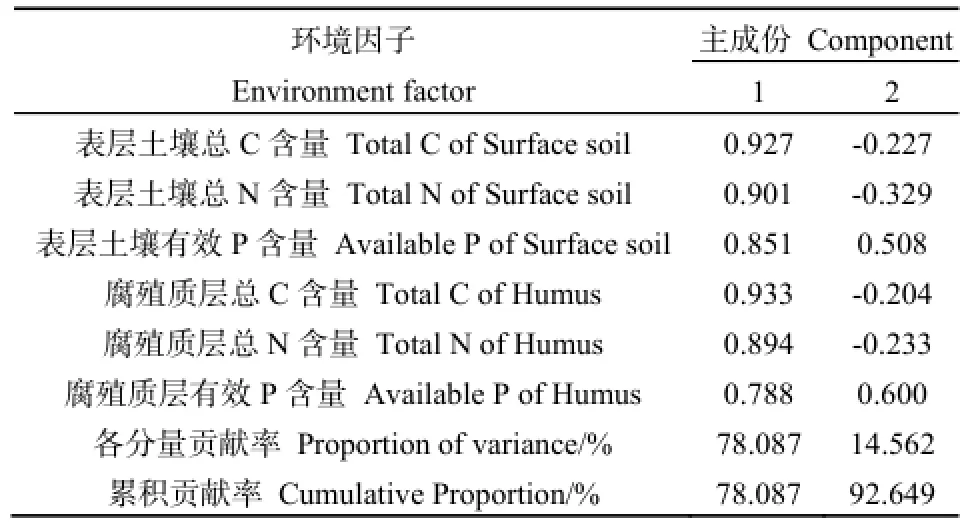
表5 不同土壤环境因子的贡献率和主分量值Table 5 The contribution rate and principal component values of different soil environmental factors
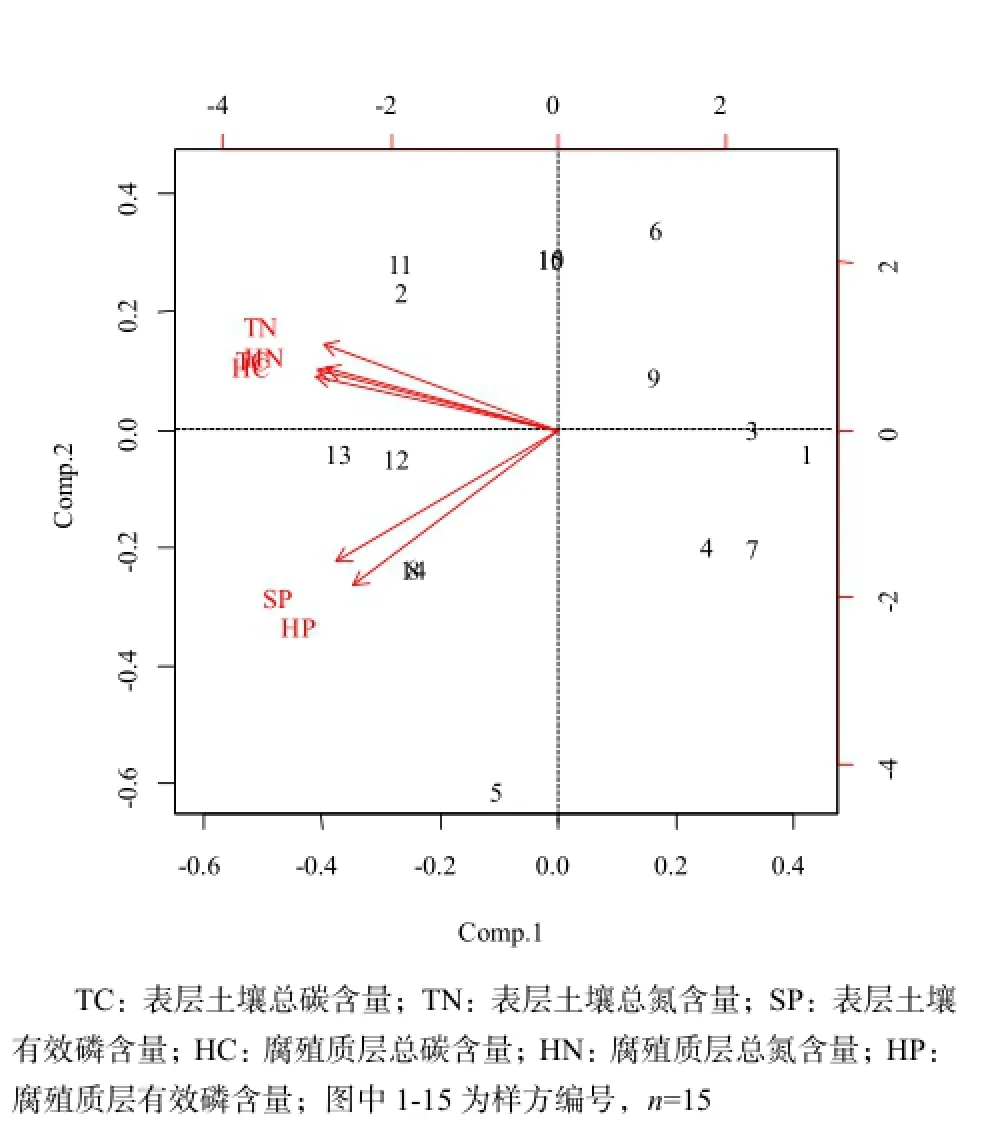
图1 主成分图Fig. 1 Principal component diagram
2.3 油松林林下植物与环境因子的CCA排序
基于野外调查和PCA分析,共得到8个样地生物及环境因子,包括林分因子(2个)、土壤环境因子(2个)、地形因子(4个),建立矩阵(Z8×15),采用CCA分析确定影响该区域油松天然林下物种分布格局的主要因子。8个生物及环境因子对林下灌木层物种分布的解释量为59.92%,采用偏分析法(Partial methods)得出林分因子即林龄、林分密度对灌木层物种分布的解释量为16.92%,土壤环境因子即土壤碳氮含量、土壤有效磷含量对灌木层物种分布的解释量为13.82%,地形因子即海拔、坡度、坡向、坡位的解释量为32.44%,尚有不能解释的物种分布变量为40.08%。根据蒙特卡罗置换检验,坡位(P=0.009,R2=0.57)和坡向(P=0.028,R2=0.42)是影响林下灌木层物种分布的最主要生境因子(图2)。
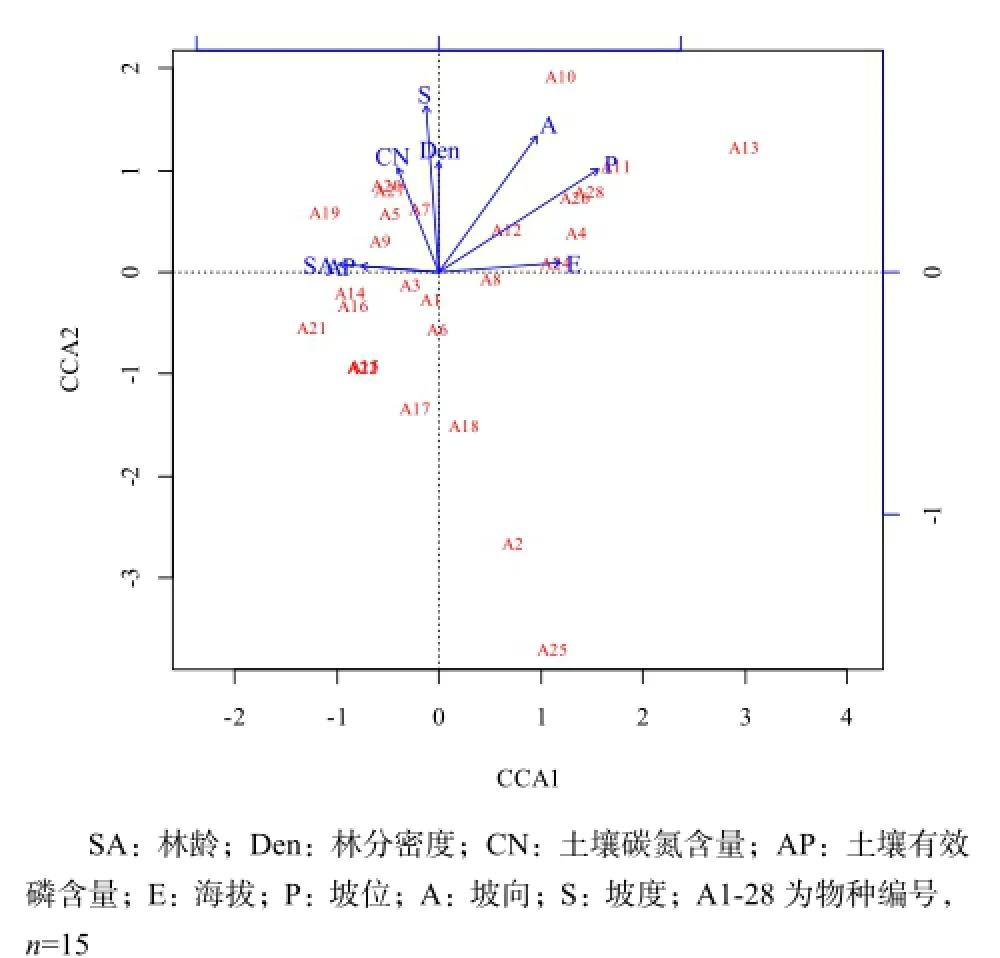
图2 林下灌木层与环境因子的CCA二维排序Fig. 2 CCA ordination diagram of shrub layer species composition and dominant environmental factors
草本层物种组成与生物及环境因子的CCA分析结果表明,第一主成分轴和第二主成分轴总共能解释草本层物种组成和生境因子关系变异的66.16%,其中林分因子对草本层物种分布的解释量为16.35%,土壤因子对草本层物种分布的解释量为13.87%,地形因子的解释量为34.38%,3组环境变量的共同解释量为1.56%,尚有不能解释的物种分布变量为33.84%。根据蒙特卡罗置换检验,影响林下草本层物种分布的最主要生境因子是海拔(P=0.030,R2=0.43)和坡位(P=0.013,R2=0.53)(图3)。
2.4 环境因子对物种多样性的影响
物种重要值和多样性相关性较为显著,分别对坡位和坡向对林下灌木层物种多样性、海拔和坡位对草本层物种多样性的影响关系进行作图(图4)。
图4中,林下灌木层物种多样性在下坡最高,其次为中坡,上坡最低,随坡向的变化表现为西北坡>东北坡>北坡>西坡;林下草本层物种多样性随坡位的变化与灌木层正好相反,在上坡最高、下坡最低,在所选样方范围内,草本层物种多样性随海拔的升高而升高。
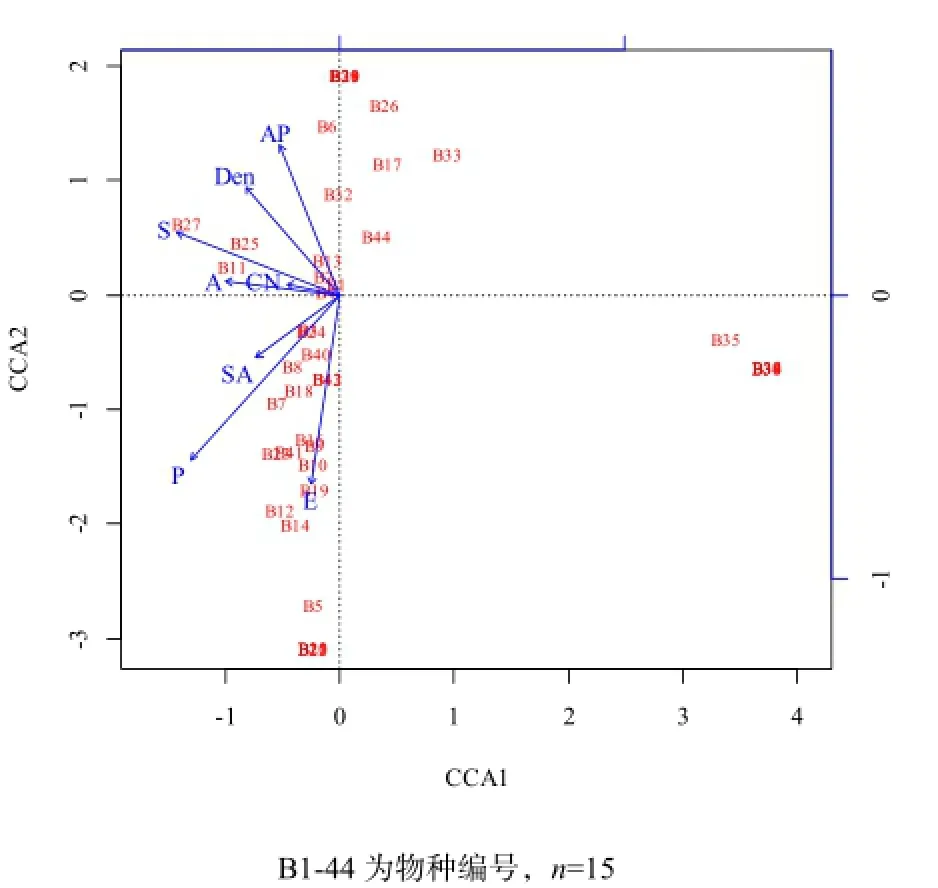
图3 林下草本层与环境因子的CCA二维排序Fig. 3 CCA ordination diagram of herb layer species composition and dominant environmental factors
3 讨论与结论
3.1 讨论
在该油松天然林林下植物中,蔷薇科、菊科、豆科所占比例较大,3大科植物在该地区的物种分布中占据优势,且在油松天然林林分中也较为重要。生态型反映植物的生物学特性,36%的林下灌木生长在林缘或疏林中,而46%的树种为兼性树种,仅18%的树种自然分布于林下或密林中,这和油松林的特性相关,油松林针叶直立、冠形特殊且均匀性较低(郭华等,2005;刘建等,2005),即使在郁闭度较高的林分中,光辐射也会透过林冠到达林下,这说明光照并不是油松林下灌木分布的决定性因素。而林下草本中干旱、半干旱的多年生草本占优势,是对该区域干旱环境的高度响应,由此可以得出水分仍然是该地区草本层物种分布的限制性因素。
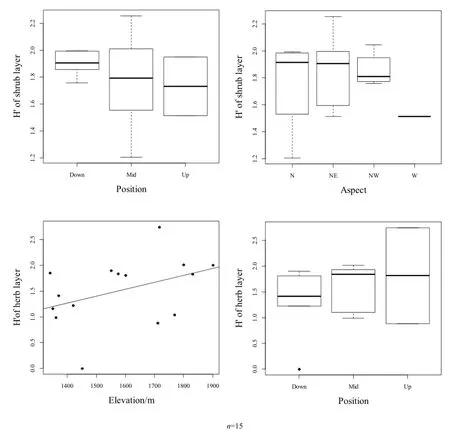
图4 林下物种多样性与环境显著因子关系Fig. 4 Relationship between understory species diversity and environmental significant factors
一般认为林下植物的生物多样性在大尺度上主要由气候类型、海拔高度所决定,而在小尺度范围内,则受到冠层类型、森林发育阶段、坡度、坡向、坡位以及光照和土壤养分条件等多种因素的共同影响。本研究的生物及环境因子对林下灌木层和草本层多样性分布格局的影响均在50%以上,表明所选的8个因子基本能代表林下植物物种分布的所有影响因素,尚未能解释的因素可能是气候条件、各种干扰以及各种因素的通过作用。在能够解释林下植物多样性分布格局的环境因子中,以地形因子所占的比例略大,表明地形因子的作用大于冠层乔木(Chipman et al.,2002)和立地条件(Huo et al.,2014),是影响油松林下植物群落组成的重要环境因子,与前人的研究结果一致(Cantón et al.,2004;Maltez-Mouro et al.,2005)。
对林下植物层起主要作用的影响因子分别为坡位、坡向和海拔,坡位和坡向直接影响了太阳辐射和水肥的空间再分配,代表着光照、水分、养分等环境因素的常年、综合生态梯度变化(刘宏文等,2014)。一般认为下坡较高的水分和养分含量会导致较高的林下植物多样性(Chipman et al.,2002),本研究灌木物种多样性在下坡较高说明影响其组成和分布的主要因素是水分和养分条件。在华北山地,下坡往往位于低海拔而上坡位于高海拔,而较短距离内海拔的快速变化会显著影响地表所受到的太阳辐射量,导致微环境、季节长度、地质状况以及人为干扰等出现较大差异(Körner,2007),从而影响了林下植物尤其是速生的草本层的分布格局(Maltez-Mouro et al.,2005),这和前人的研究结论相似(Li et al.,2011),可能是由于高海拔地区具有较高的光照有效性引起的。
3.2 结论
在本研究调查的样地中,关帝山油松天然林林下共出现植物物种72种,隶属33科65属,其中蔷薇科、菊科、豆科植物种类丰富,林下灌木多为喜光或兼性树种,林下草本以多年生草本占优势。该区生物因子与非生物因子共同对油松天然林林下植物的分布格局产生作用,对林下灌木层和草本层的解释率分别为59.92%和66.16%,其中地形因子的作用大于林分因子和土壤因子。林下灌木层物种多样性在下坡最高、上坡最低,并表现为西北坡高于其他坡向;林下草本层物种多样性随坡位的变化与灌木层正好相反,在上坡最高、下坡最低,在所选样方范围内,草本层物种多样性随海拔的升高而升高。
ALATALO R V. 1981. Problems in the measurement of evenness in ecology [J]. Oikos, 37(2): 199-204.
BARTELS S F, CHEN H. 2013. Interactions between overstorey and understorey vegetation along an overstorey compositional gradient [J]. Journal of Vegetation Science, 24(24): 543-552.
BROSOFSKE K D, CHEN J Q, CROW T R. 2001. Understory vegetation and site factors: Implications for a managed Wisconsin landscape [J]. Forest Ecology and Management, 146(1-3): 75-87.
CANTÓN Y, BARRIO G D, SOLÉ-BENET A, et al. 2004. Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain [J]. Catena, 55(3): 341-365.
CHASTAIN JR R A, CURRIE W S, TOWNSEND P A. 2006. Carbon sequestration and nutrient cycling implications of the evergreen understory layer in Appalachian forests [J]. Forest Ecology and Management, 231(1-3): 63-77.
CHEN H, LÉGARÉS, BERGERON Y. 2004. Variation of the understory composition and diversity along a gradient of productivity in Populus tremuloides stands of northern British Columbia [J]. Canadian Journal of Botany, 82(9): 1314-1323.
CHIPMAN S J, JOHNSON E A. 2002. Understory vascular plant species diversity in the mixed wood boreal forest of Western Canada [J]. Ecological Applications, 12(2): 588-601.
CHIPMAN S J, JOHNSON E A. 2002. Understory vascular plant species diversity in the mixedwood boreal forest of Western Canada [J]. Ecological Applications, 12(2): 588-601.
GAMFELDT L, SNÄLL T, BAGCHI R, et al. 2013. Higher levels of multiple ecosystem services are found in forests with more tree species [J]. Nature Communications, 4: 1-8.
GRIFFITHS M E, LAWES M J, TSVUURA Z. 2007. Understorey gaps influence regeneration dynamics in subtropical coastal dune forest [J]. Plant Ecology, 189(2): 227-236.
HUO H, FENG Q, SU Y H. 2014. The Influences of Canopy Species and Topographic Variables on Understory Species Diversity and Composition in Coniferous Forests [J]. The Scientif i c World Journal, (2): 252489.
KÖRNER C. 2007. The use of “altitude” in ecological research [J]. Trends in Ecology & Evolution, 22(11): 569-574.
LÉGARÉ S, BERGERON Y, LEDUC A, et al. 2001. Comparison of the understory vegetation in boreal forest types of southwest Quebec [J]. Canadian Journal of Botany, 79(9): 1019-1027.
LI L P, WANG X P, ZERBE S, et al. 2011. Altitudinal patterns of stand structure and herb layer diversity of Picea schrenkiana forests in the central Tianshan Mountains, Northwest China [J]. Journal of Arid Land, 3(4): 254-260.
MALTEZ-MOURO S, GARCÍA LV, MARAÑÓN T, et al. 2005. Understory floristic heterogeneity within a Mediterranean oak forest: how much is explained by edaphic, spatial and overstory components? [J]. Revista de Biologia (Lisboa), 23: 121-133.
NILSSON M C, WARDLE D A. 2005. nderstory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest [J]. Frontiers in Ecology and the Environment, 3(8): 421-428.
QIAN H, KLINKA K, ØKLAND R H, et al. 2003. Understorey vegetation in boreal Picea mariana and Populus tremuloides stands in British Columbia [J]. Journal of Vegetation Science, 14(2): 173-184.
RIEGEL G M, MILLER R F, KRUEGER W C. 1992. Competition forresources between understory vegetation and overstory Pinus ponderosa in northern Oregon [J]. Ecological Applications, 2(1): 71-85.
WU Y C, LI Z C, CHENG C F, et al. 2014. Responses of Soil Organic Carbon to Long-Term Understory Removal in Subtropical Cinnamomum camphora Stands [J]. International Journal of Ecology, (4): 1-6.
丛静, 尹华群, 卢慧, 等. 2013. 神农架保护区典型植被的物种多样性和环境解释[J]. 林业科学, 49(5): 30-35.
郭华, 王孝安. 2005. 黄土高原子午岭人工油松林冠层特性研究[J].西北植物学报, 25(7): 1335-1339.
刘宏文, 程小琴, 康峰峰. 2014. 油松人工林林下植物群落变化及其环境解释[J]. 生态学杂志, 33(2): 290-295.
刘建, 何维明, 房志玲. 2005. 东灵山油松林和辽东栎林下土壤资源和光资源的空间特征[J]. 生态学报, 25(11): 2954-2960.
孟祥楠, 赵雨森, 郑磊, 等. 2012. 嫩江沙地不同年龄樟子松人工林种群结构与林下物种多样性动态[J]. 应用生态学报, 23(9): 2332-2338.
尤业明, 徐佳玉, 蔡道雄, 等. 2016. 广西凭祥不同年龄红椎林林下植物物种多样性及其环境解释[J]. 生态学报, 36(1): 164-172.
Species Composition and Environmental Factors of Understory Plant Communities in Pinus tabulaeformis Garr. Natural Forest of Guandi Mountain
WANG Hui, NAN Hongwei, LIU Ning
College of Forestry, Shanxi Agricultural University, Taigu 030801, China
As one of the important components of forest ecosystem, understory plant communities plays an important role in regulation of forest structure and function. This paper studied 15 plots in Guandi mountain, north China, to explored the relationship between understory plant communities and environment factors, and revealed the key controlling factors of distribution pattern of understory plant communities in Pinus tabulaeformis natural forest.The results showed that: (1) 72 understory species belonging to 33 families and 65 genera were recorded on 15 plots, mainly in Rosaceae, Compositae and Leguminosae accounting for 38.8% in total species. Shrubs mostly are heliophile or facultative species, which had close relations with special crown-shape of Pinus tabulaeformis forest, and perennial herbs were dominant living types. (2) The results of CCA clearly reflected the relationship between the distribution of understory plants and environment factors. Biological factors and non-biological factors work together to the distribution pattern of understory plant, explained 59.92% and 66.16% of the variation in the shrub and herb layer, respectively. The effect of topographical factor was greater than stands or soil factors, plays an important role in understory plants’ composition and distribution. The species diversity of shrub layers were highest on down-slope and lowest on up-slope, northwest-wards was higher than other direction; herb layers were highest on up-slope, while lowest on down-slope and increasing along with the altitude.
mountainous regions of north China; Pinus tabulaeformis Garr. natural forest; understory plant; distribution pattern; environmental factors
10.16258/j.cnki.1674-5906.2017.01.003
Q948; X17
A
1674-5906(2017)01-0013-07
王慧, 南宏伟, 刘宁. 2017. 关帝山油松天然林林下植物组成及环境解释[J]. 生态环境学报, 26(1): 13-19.
WANG Hui, NAN Hongwei, LIU Ning. 2017. Species composition and environmental factors of understory plant communities in Pinus tabulaeformis Garr. natural forest of Guandi Mountain [J]. Ecology and Environmental Sciences, 26(1): 13-19.
国家自然科学基金项目(31470630)
王慧(1985年生),女,讲师,博士,研究方向为森林培育。E-mail: sxauwh@163.com *通信作者
2016-11-28
