暗紫贝母种子形态结构研究
2017-03-31陈发军陈文年
黄 艳,陈发军,陈文年
(内江师范学院生命科学学院,四川 内江 641112)
暗紫贝母种子形态结构研究
黄 艳,陈发军,陈文年
(内江师范学院生命科学学院,四川 内江 641112)
用显微镜观察和电子天平称量相结合的方法,从外部形态和内部结构等方面对暗紫贝母种子进行了研究。结果表明,暗紫贝母种子种皮薄、细胞层次少,对种子的保护作用较小,使种子容易损伤、霉变和腐烂;种胚发育不完善,未分化出胚芽、胚轴、胚根及子叶等结构,说明胚在结构上并未成熟。因此,暗紫贝母种子萌发率低,主要跟种皮保护作用缺失和种胚发育不完善两方面的因素有关。
暗紫贝母种子;外部形态;内部结构;萌发率
暗紫贝母(Fritillaria unibracteata)为百合科贝母属多年生草本植物,是一种十分珍贵的药用植物,其鳞茎具有清热润肺、祛痰止咳的功效,主要分布于四川西部、青海南部及甘肃南部,生长于海拔2 800~4 400 m的高寒灌丛草地和高寒草甸中,分布范围十分有限[1,2]。
暗紫贝母在目前中药材市场上需求量较高,但其药用部分——鳞茎生长速度缓慢,从种子长成商品药材一般需要4年左右的时间[3]。由于商品药材主要依靠野生资源,导致采挖过度,产量急剧下降,造成自然资源极度匮乏、日趋枯竭,长期处于供不应求的状态[4]。因此,暗紫贝母人工种植成为解决供需矛盾的重要途径,具有极其重要的意义,而利用种子进行育苗又是人工栽种的重要方式。
但是,在自然条件下,暗紫贝母种子的萌发率相当低下,即使在人工栽培条件下,不经过适当处理的暗紫贝母种子萌发率也很低。那么,暗紫贝母种子萌发率低是否跟其自身的形态及结构有关呢?
本研究旨在对暗紫贝母种子进行形态结构方面的研究,试图找出导致其萌发率低的形态与结构方面的原因,为人工种子育苗提供理论依据。
1 材料与方法
1.1 材料
1.1.1 种子来源
暗紫贝母种子采集于2014年9月,原产地——四川省阿坝州松潘县。采集后得到的成熟种子于冰箱中低温暂时冷藏。
1.1.2 实验器材
显微镜、解剖镜、电子天平(万分之一)及烘干箱。
1.2 方法
1.2.1 暗紫贝母种子外部形态特征观察
随机选取若干粒饱满的暗紫贝母种子,置于解剖镜载物台上,观察种子的形状、颜色、质地等外部形态特征;同时,轻轻挑开种皮,透过半透明的胚乳观察其中胚的大小。
1.2.2 暗紫贝母种子内部结构观察
随机选取若干粒暗紫贝母种子,清水浸泡72 h,用吸水纸吸掉种子表面的水,置于解剖镜载物台上,用解剖针剥开种皮,并将种胚与胚乳分离开来,对种皮、种胚、胚乳各部分分别进行观察。之后,将种皮制成装片,在显微镜下观察其结构。
按上述方法,同样选取若干粒玉米及小麦种子,浸泡72 h,吸水完全后,用解剖针分别将每粒种子的种皮、种胚、胚乳进行剥离,并分别观察。同样将种皮制成装片,在显微镜下观察其结构。
1.2.3 暗紫贝母种子内部结构各部分干重所占比例及胚比测定
随机选取100粒暗紫贝母种子,清水浸泡72 h,用滤纸吸掉种子表面的水,置于解剖镜载物台上,用解剖针分别将每粒种子的种皮、种胚、胚乳进行剥离。之后将种子各部分分别放于烘干箱里,在60℃温度下烘48 h。然后用万分之一电子天平称取种子各部分质量,计算出各部分干重占整粒种子干重的百分比,同时计算胚比(胚/胚乳),重复实验9次。
按上述相同的方法,同样选取100粒玉米种子,浸泡72 h,吸水完全后,用解剖针分别将每粒种子的种皮、种胚、胚乳进行剥离、烘干和称量。计算出各部分干重所占整粒种子干重的百分比及胚比,也重复实验9次。
2 结果与分析
2.1 暗紫贝母种子外部形态特征
暗紫贝母种子外观扁平,呈三角倒卵形。种皮浅棕黄色至棕黄色,在解剖镜下放大后,可明显观察到种皮有褶皱纹理。大多数暗紫贝母种子在解剖镜下能观察到种胚,但胚所占的空间比例极小,相反,胚乳含量相对较丰富,其在种子中所占比例也较大,如图1所示。
2.2 暗紫贝母种子内部结构
2.2.1 种皮结构
暗紫贝母种皮薄而透明,将种皮制成装片在显微镜下观察,如图2所示。
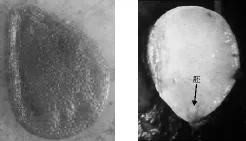
图1 暗紫贝母种子外部形态(80×)
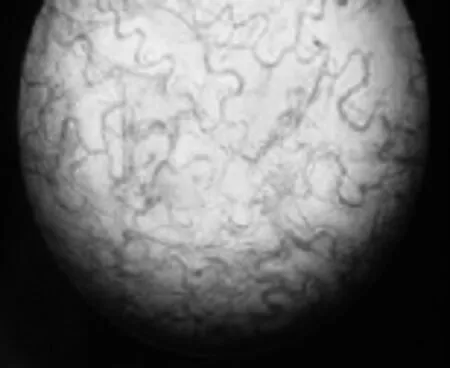
图2 暗紫贝母种皮显微结构(400×)
由图2可以明显观察到,种皮细胞间轮廓与纹理清楚,显然暗紫贝母种皮薄、细胞层次少,往往就是1~2层细胞。由于缺乏相对的坚韧性,种皮对种子起不到很好的保护作用,导致种子容易受损、霉变、腐烂等。
与暗紫贝母相比,日常生活中萌发率较高的玉米、小麦种子,其种皮与果皮愈合在一起,为多层细胞结构。将玉米种皮、小麦种皮置于显微镜下观察,如图3、图4所示。

图3 玉米种皮显微结构(400×)
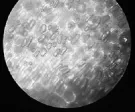
图4 小麦种皮显微结构(400×)
从图3和图4可以看出,玉米与小麦种皮厚实、细胞层次较多。由于种皮细胞重重叠叠,不能观察到相对清楚的细胞轮廓与纹理,这样的种皮对种子往往能起到较好的保护作用。
2.2.2 种胚结构
暗紫贝母种胚极其微小,在种子中所占比例很小,肉眼几乎观察不到。将种胚剥离出来,在显微镜下观察,只能看到一团细胞的原胚,不能观察到胚芽、胚轴、胚根和子叶,这表明其胚的发育还很不充分、很不完整。
与暗紫贝母相比,日常生活中萌发率较高的小麦、玉米种子,其种胚往往已发育完善,如图5、图6所示。
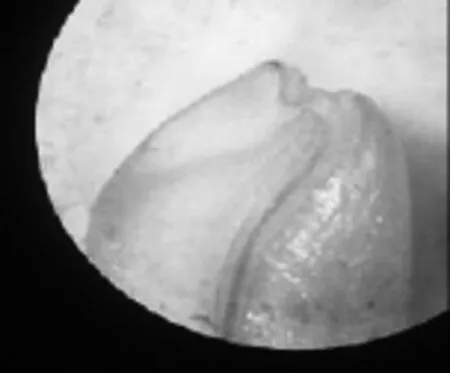
图5 小麦种胚结构(80×)
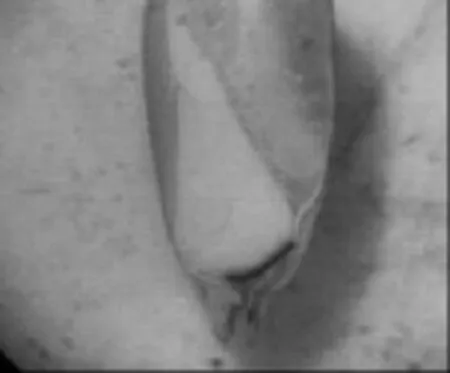
图6 玉米种胚结构(80×)
由图5、图6可以观察到,小麦种胚与玉米种胚中已分化出胚芽、胚轴、胚根以及子叶4个部分。
2.2.3 各部分干重所占比例及胚比
暗紫贝母种子种皮干重、胚干重、胚乳干重在整粒种子中所占比例及胚比见表1,以玉米种子作为对比。
由表1可以得出,暗紫贝母种胚含量极少,只占整个种子干重的2.65%(平均值),其胚乳含量相对多一点,占整个种子干重的比例为66.29%,其胚比平均为0.040。相对而言,玉米种胚含量较暗紫贝母多得多,所占干重比例平均为10.30%,胚乳所占比例为83.32%,胚比为0.124。显然,暗紫贝母与玉米相比,种胚所占比例低得多,仅为玉米的1/4左右,胚比也仅为玉米的1/3左右。另外,从表1中还可以看出,暗紫贝母种皮在种子中占有相对较高的比例,而胚和胚乳所占的比例则相对低一些。

表1 暗紫贝母种子内部结构各部分干重所占比例及胚比
3 结论与讨论
造成种子萌发率低的原因有很多,总体来说,受到外部环境因素和种子内部形态结构两方面的影响[5]。外部环境因素,主要包括种子萌发必需的光照、温度、水分等;而当种子处于适宜条件下时,外部环境对种子萌发的影响作用并不大,这时其萌发主要取决于种子的内部结构。就其内部结构来说,种子萌发困难主要受种皮和种胚的影响。本研究表明,暗紫贝母种皮的层次结构与种胚的形态结构是造成暗紫贝母种子萌发率极低的主要原因。
一般而言,种皮主要是保护种子,可以起到防止机械损伤和病虫害入侵的作用[6]。通过本研究发现,暗紫贝母的种皮薄,不仅起不到应有的保护作用,反而成为影响种子萌发的主要因素之一。尽管其种皮在整个种子中所占比例较大,达到31.06%(表1),但其种皮细胞层次少、种皮极薄,不能很好地保护种子,使种子容易受到损伤、霉变和腐烂。另外在研究中还发现,暗紫贝母种皮虽然很薄,但种皮细胞外表面往往具有一层明显的脂类物质(蜡质),在种皮破损之前,这层物质会影响水分和气体的进入,影响种子的透水和透气性,导致种子萌发条件难以满足而不能萌发。
种子在外界环境条件达到适宜时,种胚是萌发的另一大关键因素。相关文献表明,很大一部分珍贵药用植物种子的胚都存在休眠现象[7],即种子虽然已经脱离了母体,但其胚的发育并不完全,还未分化出相应的组成部分[8]。本研究中的暗紫贝母种子就属于这种情况,虽在采收时已表现成熟,但种胚极小,尚未分化完善,还需要较长的一段时间来完成其形态上的发育成熟;孟繁蕴等对滇重楼种子进行研究时也得出了类似的结论[9]。种子萌发实质上就是种胚长成幼苗的过程,如果胚本身都没有发育完全,即使外界环境条件适宜,种子其他部位不对其产生阻碍作用,胚自身仍然不能得到萌发[10]。
综上所述,种皮的保护作用缺失和种胚的发育不完善,是导致暗紫贝母种子萌发率低的主要因素。
[1]四川植物志编辑委员会.四川植物志(第七卷)[M].成都:四川民族出版社,1991:57-59.
[2]徐国钧.中草药彩色图谱(修订本)[M].福州:福建科学技术出版社,1997:210-213.
[3]GAO S L,ZHU D N,CAI Z H,et a1.Organ culture of a precious Chinese medicinal plant-Fritillaria unibracteata[J].Plant Cell,Tissue and Organ Culture,1999,59(3):197-201.
[4]马吉义,陈志,李冬鸣.中药暗紫贝母的研究进展[J].安徽农学通报,2010,16(15):216-218.
[5]程汉亭,范志伟,李鸣光,等.金钟藤种子低萌发率原因探讨[J].广西植物,2012,32(4):522-526.
[6]戴思慧,孙小武,熊兴耀,等.三倍体西瓜种子萌发障碍与种皮的关系[J].湖南农业大学学报(自然科学版),2012,38(4):392-397.
[7]胡莹莹.川贝母种子萌发特性及育苗技术研究[D].成都:四川农业大学,2008:1-54.
[8]孙婷.栝楼(Trichosanthes kirilowii Maxim)种子的休眠特性及促萌措施研究[D].金华:浙江师范大学,2010:1-69.
[9]孟繁蕴,汪丽娅,张文生,等.滇重楼种胚休眠和发育过程中内源激素变化的研究[J].中医药学报,2006,34(4):36-38.
[10]HILHORST H W M.A critical update on seed dormancy.I.Primary dormancy[J].Seed Science Research,1995,5(2):61-73.
2016-12-15)
四川省科技厅应用基础项目(2014JY0130);四川省教育厅成果转化重大培育项目(13CZ0001);四川省教育厅创新团队项目(14TD0025);四川省教育厅科研项目(15ZB0267),内江师范学院植物学重点建设学科项目资助。
陈文年(1968-),男,四川成都人,教授,从事植物学方面教学与科研。
1005-2690(2017)03-0124-03
S567.23+1
B
