一个木薯候选STK类抗病基因的分离及其生物信息学分析
2017-03-31陈奕鹏李超萍樊春俊时涛李博勋黄
陈奕鹏 李超萍 樊春俊 时涛 李博勋 黄贵修


摘 要 根据丝氨酸/苏氨酸蛋白激酶类(STK)结构域Ⅰ和Ⅷ氨基酸保守序列设计了1对简并引物,以细菌性萎蔫病抗/感木薯种质E1340和GR911基因组DNA为模板,通过PCR扩增,均获得大小约0.5 kb的扩增片段。两个片段纯化、克隆、测序后获得完全一致的505 nt序列,比对发现,木薯种质AM560带有和该序列高度同源的核苷酸片段。对包括同源片段上下游各约2.0 kb的大片段序列进行基因预测,获得1个有4个外显子、ORF全长1866 nt的预测基因,命名SSK1。序列分析表明,该基因编码621 aa,有细胞壁受体激酶结合、偶联位点和8个跨膜结构域,具有典型的STK类抗病基因的保守结构,是一个候选的抗病基因,可能在木薯抗病反应中发挥重要作用。
关键词 木薯 ;候选抗病基因 ;丝氨酸/苏氨酸蛋白激酶
中图分类号 S533 文献标识码 A Doi:10.12008/j.issn.1009-2196.2016.12.006
木薯(Manihot esculenta Crantz)为大戟科(Euphorbiaceae)木薯属(Manihot Mill.)灌木状多年生作物,是世界三大薯类(马铃薯、甘薯、木薯)之一,也是全球仅次于小麦、玉米、水稻、马铃薯、高粱的第6大粮食作物。木薯块根富含淀粉,是主要的收获物,是全球10亿人口日常生活中所需热能的主要来源。目前,木薯在我国华南地区广泛种植,以之为基础的食品和生物能源等产业是当地农业经济的重要部分[1]。木薯种植中,各类严重发生的病害是相关产业发展的限制性因素之一。国外报道为害木薯的病害多达30余种[2]。中国热带农业科学院环境与植物保护研究所的调查结果表明,危害中国木薯的病害有8种,其中细菌性萎蔫病最为严重[3]。种植抗病品种是作物病害防治中最为有效的措施之一,相关抗病基因资源是开展抗病育种工作的前提。目前木薯抗病机理方面的研究还很少,也缺少可供利用的基因资源,严重制约了相关研究工作的开展。
基因对基因假说[4]认为,寄主植物带有多个抗病基因,病原菌带有对应的无毒基因,二者的蛋白产物互作后通过一系列的级联反应调控植物表现出抗病性[5]。目前模式植物拟南芥(Arabidopsis thaliana)和番茄(Lypoersicum esculentum)抗病基因方面的研究最多[6-8]。已克隆的抗病基因蛋白产物通常具有1种或多种保守结构,丝氨酸/苏氨酸蛋白激酶(STK)结构域是抗病基因蛋白产物常见的结构之一。该类激酶作用方式包括蛋白质间相互作用,转录水平和翻译水平调控和DNA复制的调节等方式[9]。以抗病基因的保守结构域设计简并或特異引物通过PCR扩增或使用RGA探针从基因组DNA或cDNA文库中获得抗病基因类似序列,再从中筛选潜在的抗病基因,是作物抗性机理研究中的重要方法。本研究根据STK的保守区序列设计简并引物,通过PCR扩增获得木薯抗病基因类似序列并进行候选基因的分离和生物信息学分析,为进一步开展相关研究提供基础。
1 材料和方法
1.1 材料
1.1.1 供试木薯品种
木薯细菌性萎蔫病抗病种质E1340和感病种质GR911[10],由中国热带农业科学院热带作物品种资源研究所木薯种质资源圃提供。
1.1.2 培养基和试剂
DNA 片段胶回收试剂盒、pMD18-T 载体购自宝生物工程(大连)有限公司。Taq酶、dNTPs和大肠杆菌Top10感受态细胞购自天根生化科技(北京)有限公司。引物由北京六合华大基因科技股份有限公司合成。大肠杆菌培养采用LB培养基(参照方中达[11]的方法制备),其他试剂均为国产分析纯。
1.2 方法
1.2.1 木薯基因组DNA的提取及目的序列扩增
田间采集健康木薯种质叶片,按闫庆祥等[12]的方法提取基因组DNA。根据小麦中与Lr10紧密连锁的一个STK激酶和水稻抗白叶枯病基因Xa21的STK激酶区保守氨基酸序列设计1对简并引物Ps-F(5′-GGNCARGGNGGDTTYGGDWSDG-3′)和Ps-R(5′-YTCNGGNGCDATRTADCCCATTG-3′)进行PCR扩增。
PCR反应体系:10×PCR缓冲液2.5 μL、10 mmol/L dNTPs 1 μL、10 μmol/L各引物溶液1 μL、DNA模板50 ng、Taq DNA 聚合酶1.0 U、补充无菌水至25 μL。PCR循环参数:94℃预变性3 min;94℃ 30 s、52℃ 30 s、72℃ 2 min,35 个循环;最后72℃延伸10 min。扩增产物用1.0 %琼脂糖凝胶电泳检测后,按试剂盒说明书进行扩增产物的纯化、回收和克隆,随机挑选3个阳性克隆送北京六合华大基因科技股份有限公司进行序列测定。
1.2.2 目的基因的预测和功能分析
在NCBI上进行预测基因同源性比对。采用本地Blast的方法,将获得的序列在木薯基因组数据库(木薯种质:AM560)中进行比对,获得其上下游序列后,在Softberry和GENSCAN上进行目的基因预测。利用SMART、NCBI中CDD数据库和Sanger中Pfam数据库对获得的编码蛋白氨基酸序列进行结构分析。用ProtParam进行氨基酸一级结构分析。用TMpred进行跨膜结构预测。将推导出的氨基酸序列与已克隆的该类基因小麦LRK10(登录号U51330.1)、LRK33(登录号AAK20740),番茄Pto(登录号U59316)和水稻Xa21(登录号U37133.1)进行氨基酸特有结构域分析。下载近源基因后,用MEGA version 4.0中的NJ(Neighbor-Joining)方法生成系统树,用Bootstrap 对系统树进行检验,1 000次重复。
2 结果和分析
2.1 木薯基因组DNA的提取及目的序列扩增
分别提取木薯抗病种质E1340和感病种质GR911基因组DNA后,用引物对Ps-F和Ps-R进行PCR扩增,均获得大小约0.5 kb的扩增产物。两个扩增片段回收、克隆后发现其序列完全一致,长度为505 nt。
2.2 目的基因的预测和功能分析
将获得的序列在NCBI上进行Blastx分析,发现该序列与来自大麦、蓖麻、玉米等作物的STK类蛋白的同源性在50%以上(表1)。
所获序列和木薯基因组(种质为AM560)比对结果表明,该种质带有同源性为98%的同源片段。提取包括该片段上下游各约2.0 kb的大片段序列,采用Softberry的Hevea(橡胶树)、Nicotiana tabacum(烟草)和Arabidopsis(拟南芥)3种模式和GENSCAN的Maize(玉米)模式进行基因预测,结果均表明该位点位于一个预测基因内部,该基因命名为SSK1(图1)。参照同属大戟科作物的Hevea模式分析结果,分析SSK1位于提取序列的负链上,有4个外显子CDSf、CDSi1、CDSi2和CDSl,TSS(转录起始位点)位于第一个外显子前0.6 kb处,PolyA位于最后一个外显子后的0.5 kb处,ORF全长1866 nt,编码621 aa。
利用SMART软件、NCBI中CDD数据库和Sanger中Pfam数据库对SSK1的基因结构预测的结果均表明,SSK1蛋白产物具有跨膜的细胞壁受体激酶结合、偶联位点(WAK bind和WAK assoc),具有典型的类PKC家族结构特征(图2),进一步分析得出,SSK1具有典型的STK类抗病基因特异结构域。
利用ProtParam分析表明該蛋白质分子量为69 643.7 ku,等电点为5.74,平均疏水性(GRAVY)为-0.018。各氨基酸中,丝氨酸、亮氨酸和异亮氨酸的含量最高,分别为66个(10.6%)、64个(10.3%)和48个(7.7%),色氨酸含量最低,为6个(1%)。由TMpred程序预测表明,SSK1所编码蛋白在第17~35,174~199,260~285,377~403个氨基酸残基间存在4个由内到外的跨膜螺旋区域,在第18~35,170~188,267~290,383~400个氨基酸残基间存在4个由外到内的跨膜螺旋区域(图3)。
将SSK1基因蛋白产物与小麦LRK10(U51330.1)、LRK33(AAK20740)、LR10(U59316)和水稻Xa21(U37133.1)等STK类抗病基因进行氨基酸序列同源性分析,结果显示SSK1编码蛋白氨基酸序列均具有STK类抗病基因所特有的9个催化结构域,其中两个特征区域分别为Ⅵb(DaKXXN)和Ⅷ(GTaGYxAP(N/E)(见图4)。
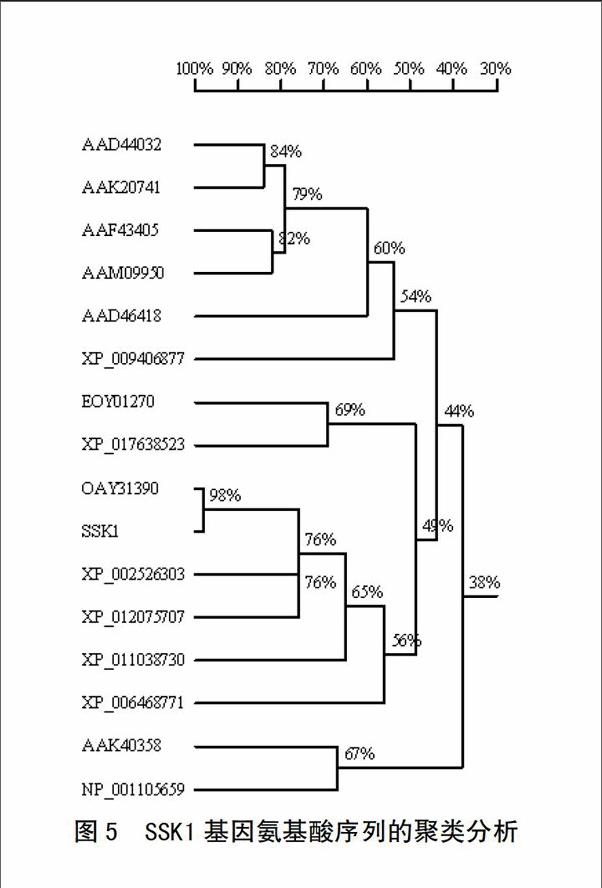
氨基酸序列比对结果表明,SSK1基因氨基酸序列和木薯预测基因MANES_14G108500(OAY31390)同源性为95%,和大戟科蓖麻预测受体类激酶基因At1g67000(XP_002526303)、麻疯树预测受体类激酶At1g67000(XP_012075707)的同源性分别为73%和72%,分析其为候选的STK类抗病基因。下载了部分近源序列后进行聚类分析,结果表明SSK1、MANES_14G108500、At1g67000、At1g67000等亲缘关系最近,和胡杨的预测受体类激酶(XP_011038730)、柑橘的预测受体类激酶(XP_006468771)为一个分枝,可可的预测激酶(EOY01270)和树棉的预测抗病受体类激酶(XP_017638523)聚类为亲缘关系较近的另一个分枝。SSK1和小麦预测的受体类激酶(AAK40358)、丝氨酸/苏氨酸受体激酶(NP_001105659)亲缘关系较远,和大麦的预测受体类激酶(AAD44032)、小麦的预测受体类激酶(AAK20741)、粳稻的预测丝氨酸/苏氨酸受体激酶(AAF43405)和预测受体类激酶(AAM09950)、玉米的预测受体类激酶(AAD46418)和柑橘的预测受体类蛋白激酶(XP_006468771)的亲缘关系最远(图5)。
3 讨论
本研究根据STK类抗病基因蛋白产物的保守结构域设计简并引物,采用PCR扩增分别从抗、感细菌性萎蔫病的木薯种质E1340、GR911基因组中分离到序列一致、和STK类蛋白具有较高同源性的1个核酸片段,比对发现木薯种质AM560同样带有该同源片段。基因预测发现该片段位于一个有4个外显子、编码621 aa的预测基因中。该基因蛋白产物典型的STK类蛋白的结构特征,聚类分析表明,该基因蛋白产物与预测的受体类激酶具有较高的同源性,可能在木薯抗病反应中发挥重要作用。
自1996年首次利用抗病基因产物的保守结构域扩增获得抗病基因类似序列RGAs(Resistance gene analogs)以来,利用RGAs方法来克隆、定位抗病基因已得到广泛应用,显示出广阔的应用前景[13]。利用该方法在马铃薯[14]、小麦[15]、大麦[16]、番茄[17]、黄瓜[18]、水稻[19]、香蕉[20]等作物上均已获得了大量的RGAs。这些RGAs与抗病基因R基因的关系可以分为3种:第一,RGA是R基因或其假基因的一部分,例如拟南芥中的一类RGA与RPP5紧密连锁,被证明是RPP5的一部分[21];第二,RGAs与R基因紧密连锁,例如水稻[22]、大麦[23-24]中都找到了与不同的R基因紧密连锁的RGA。安海山等[25]发现,仅有抗病核桃品种基因组中能分离到NBS类RGAs,表明这些序列是和抗性相关的;第三,有些非抗病基因同样具有NBS、LRR和丝氨酸/苏氨酸酸激酶等保守域,因此,克隆出的RGA有可能和目的抗病基因无关。
Lopez等[26]根据抗病基因的NBS(核苷酸结合位点)和TIR(白细胞介素-1受体)保守域设计特异引物,从木薯中分离到12类RGA,文库筛选和杂交分析发现其中11类RGA在基因组中为单拷贝。本研究采用类似方法获得预测基因SSK1,同时发现,木薯对细菌性萎蔫病抗感种质中均带有SSK1的同源基因,该基因是否参与木薯对细菌性萎蔫病或其它病害的抗性反应,还需要开展进一步的研究。
参考文献
[1] 方 佳,濮文辉,张慧坚. 国内外木薯产业发展近况[J]. 中国农学通报, 2010, 26(16): 353-361.
[2] 李开绵,黄贵修. 木薯主要病虫害[M]. 北京: 中国农业科学技术出版社,2008:142-144.
[3] 李超萍,时 涛,刘先宝,等. 国内木薯病害普查及细菌性萎蔫病安全性评估[J]. 热带作物学报,2011,32(1):116-121.
[4] Flor H H. Current status of the gene-for-gene concept[J]. Annu Rev Phytopathol, 1971(9): 275-296.
[5] Sastkwicz B J, Ausubel F M, Bkaer B J, et al. Molecular genetics of plant disease resistance[J]. Science, 1995, 268(5): 661-667.
[6] Marin G B, Brommonschenke S H, Chunwongse J, et al. Map-based cloning of a protein kinase gene conferring disease resistance in tomato[J]. Science, 1993, 262: 1 432-1 436.
[7] Salmeron J M, Oldroyd G E D, Staskawicz B J. Tomato PRf is a member of leucine-rich repeat class of plant disease resistance genes and lies embedded within the Pto kinase gene cluster[J]. Cell, 1996, 86: 123-124.
[8] Liu J L, Liu X L, Dao L Y, et al. Recent progress in elucidating the structure, function and evolution of disease resistance genes in plants[J]. Journal of Genetics and Genomics, 2007, 34 (9): 765-776.
[9] Nevin D Y. The genetic architecuter of resisnatce[J]. Cuerrnt Opinion in Plnat Biology, 2000, 3: 285-290.
[10] 卢 昕,李超萍,时 涛,等. 国内603份木薯种质对细菌性枯萎病抗性评价[J]. 热带农业科学,2013,33(4):67-70,90.
[11] 方中达. 植病研究方法(第3版)[M]. 北京. 中国农业出版社,1998.
[12] 闫庆祥,黄东益,李开绵,等. 利用改良CTAB法提取木薯基因组DNA[J]. 中国农学通报,2010,6(4):30-32.
[13] 易图永,谢丙炎,张宝玺,等. 植物抗病基因同源序列及其在抗病基因克隆与定位中的应用[J]. 生物技术通报,2002(2):16-20.
[14] Leister D, Ballvora A, Salamin F, et al. A PCR-based approach for isolating pathogen resistance genes from potato with potential for wide application in plants[J]. Nature Genet, 1996, 13: 421-429.
[15] Leister D, Kurth T, Schulze P, et al. RFLP and physical mapping of resistance gene homologues in rice (Q.sative) and Barley (H.vulgare)[J]. Theor Appl genet, 1999, 98: 509-520.
[16] 张 楠,王海燕,刘大群. 小麦STK类抗病基因同源序列的克隆和分析[J]. 华北农学报,2010,25(5):20-24.
[17] Leiser D, Baslvora A, Salamini F, et al. A PCR-based approach for isolating pathogen resistance gene from potato with potential for wide application in plants[J]. Nat Genet, 1996(4): 421-429.
[18] 丁国华,池春玉,周秀艳,等. 黄瓜抗病基因类似序列(RGA)的同源性分析和Southern鉴定[J]. 园艺学报,2007,34(2):355-360.
[19] 廖 承,黎 帅,晋佳路. 水稻抗病基因Lrr19类似物序列分析研究[J]. 現代农业科技,2013,15:13-15.
[20] 孟祥春,徐金刚,黄秉智,等. 香蕉抗病基因类似序列的克隆与分析[J]. 分子植物育种,2007,6(5):57-60.
[21] Aarts M G M, Lintel Hekkert B,Holub E B, et al. Identification of R-gene homologous DNA fragments genetically linked to disease resistance loci in Ardbidop.sis chdlidnd[J]. Mol. Plant Microbe Interact,1998, 11(4): 251-258.
[22] Chen X M. Line R F, Leung H. Genome scanning for resistance-gene analogs in rice barley, and wheat by high-resolution electrophoresis[J]. Theor. Appl. Genet., 1998, 97(3): 345-355.
[23] Madsen L H. Collins N C. Rakwalska M. et al. Barley disease resistance gene analogs of the NBS-LRR class: identification and mapping[J]. Molecular Genetics & Genomics, 2003, 269(1): 150-161.
[24] Mohler V, HIahr A, Wenzel G, et al. A resistance gene analog useful for targeting disease resistance genes against different pathogens on group 1S chromosomes of barley, wheat and rye[J]. Theor. Appl. Genet., 2002, 105 (2/3): 364-368.
[25] 安海山,杨克强. 核桃NBS类抗病基因类似物的序列特征及其与炭疽病的抗性[J]. 中国农业科学,2014,47(2):344-356.
[26] Lopez C E, Zuluaga A P, Cooke R, et al. Isolation of Resistance Gene Candidates (RGCs) and characterization of an RGC cluster in cassava[J]. Mol Gen Genomics, 2003, 269: 658-671.
